










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.12 no.1 Córdoba Jan./June 2002
TRABAJOS ORIGINALES
Buceo eficiente en aguas poco profundas: comportamiento de buceo y patrón de alimentación del Cormorán Cuello Negro, Phalacrocorax magellanicus, en dos colonias de la costa patagónica
Flavio Quintana1,2,*, Federico Morelli3 & Yanina Benedetti3
1 Centro Nacional Patagónico (CENPAT) - CONICET, Puerto Madryn, Chubut, Argentina
2 Wildlife Conservation Society, Bronx, New York, EEUU
3 Universidad Nacional de la Patagonia San Juan Bosco, Puerto Madryn, Chubut, Argentina
* Centro Nacional Patagónico - CONICET; Blvd. Brown s/n; 9120 Puerto Madryn; Chubut, Argentina. Email: quintana@cenpat.edu.ar
Recibido: 13 septiembre 2001;
Revisado: 14 febrero 2002;
Aceptado: 20 febrero 2002
RESUMEN. Entre 1996 y 1998 se estudió, por medio de técnicas de radiotelemetría, el comportamiento de buceo y alimentación del Cormorán Cuello Negro (Phalacrocorax magellanicus) en Punta Loma y en Isla Vernacci, dos colonias de reproducción de la costa patagónica. En ambas colonias, los cormoranes bucearon por períodos prolongados (47–50 s) seguidos por breves intervalos de recuperación en superficie (14–18 s) y presentaron eficiencias de buceo altas (3.2–3.8, tiempo de buceo/tiempo en superficie). Tanto en Punta Loma como en Isla Vernacci, las aves permanecieron en el mar casi un tercio del total de horas luz (31–35%), bucearon la mayor parte del tiempo que estuvieron fuera de la colonia (aprox. 90%), realizaron numerosos viajes de alimentación diarios (2.6–3.7 viajes/día), de duración prolongada (1.7–2.6 h) y desarrollaron un elevado número de buceos por viaje (78–138). Estos resultados sugieren un alto esfuerzo de alimentación (tiempo dedicado al buceo por viaje) para cubrir los requerimientos energéticos propios y alimentar a los pichones. Una reducida disponibilidad de alimento (distribución, abundancia y accesibilidad) y el bajo rendimiento energético de las presas consumidas podrían ser algunos de los factores responsables del alto esfuerzo de alimentación sugerido.
ABSTRACT. Efficient dive in shallow waters: diving behavior and feeding pattern of the Rock Shag, Phalacrocorax magellanicus, at two Patagonian colonies: By means of radio-telemetry techniques, we study the diving behavior and feeding performance of adult Rock shags (Phalacrocorax magellanicus) from Punta Loma and Isla Vernacci, two colonies of the Patagonian coast, Argentina. In both colonies, Rock shags dived continuously (without diving bouts) during foraging trips (mean dive duration: 47-50 s, mean subsequent surface interval: 14-18 s) and showed high dive efficiencies (3.2–3.8, dive time/recovery time). Tracked birds from the two colonies spent 31- 35% of daylight hours at sea, diving (including dive and recovery times) for 92% of the foraging trip. They undertook many (2.6–3.7 trips/day) and prolonged (1.7–2.6 h) trips per day, and made a high number of dives per foraging trip (78–138). These results suggest a high foraging effort (diving time per foraging trip) for Rock shags from the two studied colonies, necessary to reach the daily food requirements. This high foraging effort could be associated with low prey abundance, the existence of a patchy and/or low quality food resource (in energetic terms), or both.
INTRODUCCIÓN
Los cormoranes son aves buceadoras que se alimentan debajo del agua propulsándose con sus patas (Ashmole 1971). La mayoría de estas especies se alimentan principalmente de peces bentónicos y de pequeños invertebrados, que capturan durante sus buceos en aguas de poca profundidad, en las cercanías de la costa (Johnsgard 1993). El buceo requiere usualmente la realización de un esfuerzo activo y representa una clara interrupción en el ritmo respiratorio del animal (Kooyman 1989). En muchas aves y mamíferos buceadores, el tiempo de ventilación en superficie se ajusta a la duración del buceo precedente (modelo de buceo reactivo), permitiendo una recuperación del esfuerzo físico realizado (Kooyman 1989). La mayor parte de las especies de cormoranes presenta un patrón de buceo caracterizado por: 1) inmersiones prolongadas, seguidas de cortos intervalos de recuperación en superficie y 2) una correlación positiva entre la masa corporal y la duración promedio de buceo (Cooper 1986). A pesar de este patrón generalizado, datos comparativos de la duración de los buceos y el tiempo de recuperación en superficie sugieren que distintas especies pueden emplear su tiempo debajo del agua de diferente forma, dependiendo de algunos parámetros de su ecología trófica tales como el tipo de presa (distribución, abundancia, disponibilidad), la profundidad de buceo o las características del fondo, entre otros (Ydenberg 1988).
Además del interés teórico, comprender el comportamiento de las aves marinas durante su permanencia en el mar y sus estrategias de alimentación resulta relevante a la hora de predecir la respuesta de sus poblaciones ante perturbaciones humanas o cambios ambientales. El patrón de alimentación y buceo puede, entonces, ser utilizado como un buen indicador de cambios en el ambiente marino (Huntley et al. 1991; Montevecchi 1993) y en la distribución y abundancia de las poblaciones presa (Monaghan et al. 1994). Por otra parte, el conocimiento preciso de la localización de las áreas de alimentación y del comportamiento de alimentación de los cormoranes permitiría comprender más detalladamente la ecología trófica de estas aves y el modo en que enfrentan una de las principales demandas de su existencia, como es la obtención de alimento.
Este trabajo se basa en el estudio comparado del patrón de alimentación (comportamiento y uso de las áreas de alimentación) y buceo del Cormorán Cuello Negro (Phalacrocorax magellanicus) durante la temporada reproductiva en dos colonias de la provincia de Chubut.
MÉTODOS
Especie de estudio
El Cormorán Cuello Negro es una especie ampliamente distribuida en la costa patagónica. Sobre la costa atlántica argentina se extiende desde Península Valdés (42°25'S; 64°31'O) hasta Tierra del Fuego, incluyendo las islas Malvinas. Sobre la costa del Pacífico en Chile, su distribución alcanza, hacia el Norte, los 37°S (Harrison 1983; Johnsgard 1993). El tamaño poblacional estimado para la especie en la costa patagónica de Argentina es de 8100 parejas (Yorio 2000). Los sitios de nidificación a lo largo de dicha costa son numerosos (n = 143) pero de tamaño reducido (<400 nidos) (Yorio et al. 1998; Yorio 2000).
Áreas de estudio
El estudio se llevó a cabo en las colonias de Punta Loma (42°49'S; 64°53'O) y de Isla Vernacci (45°11'S; 66°29'O), Chubut (Figura 1). Ambas colonias están separadas por 300 km de costa. La colonia de Punta Loma se encuentra en el golfo Nuevo, es continental, posee un total de 164 nidos y está situada sobre una costa de acantilados de 30–50 m de altura y playas de canto rodado (Yorio et al. 1998). La isla Vernacci es la más externa de un complejo de islas dentro de la caleta Malaspina, ubicada al norte del golfo San Jorge (Figura 1). En esta colonia, el Cormorán Cuello Negro (188 nidos) nidifica en simpatría con el Cormorán Imperial (Phalacrocorax atriceps) sobre superficies de escasa pendiente, a no más de 10 m sobre el nivel del mar (Yorio et al. 1998).
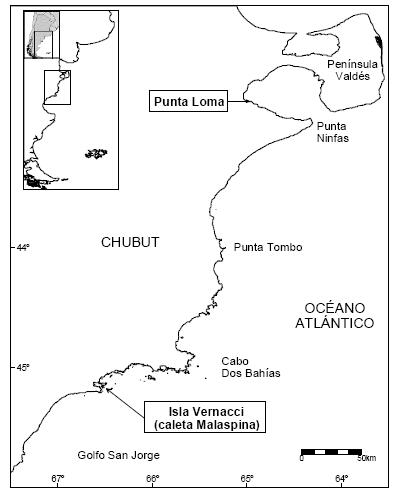
Figura 1. Mapa del área de estudio mostrando la ubicación de Punta Loma e Isla Vernacci, donde se encuentran las colonias estudiadas de Cormorán Cuello Negro.
Figure 1. Map of the study area showing the location of Punta Loma and Isla Vernacci where studied colonies of the Rock Shag are located.
En general, los sedimentos del fondo marino en las proximidades de ambas colonias están compuestos por grava, arena y arcillas limosas (Mouzo et al. 1978; Haller et al., CENPAT y Univ. Nac. de la Patagonia, com. pública; cartas náuticas H-215 y H-264, Servicio de Hidrografía Naval de la República Argentina). Los sustratos blandos presentan una comunidad constituida principalmente por anélidos poliquetos, crustáceos y nemertinos (Herrera 1997; C Pastor, CENPAT, com. pers.). En las acumulaciones de aglomerados y rodados semicubiertos de fango se desarrolla una comunidad muy diversa donde pueden distinguirse desde peces bentónicos (nototénidos) hasta otras especies de hábitos más pelágicos pertenecientes a la familia Atherinopsidae (A Gosztonyi, CENPAT, com. pers.). Las aguas costeras en las cercanías de la colonia de Punta Loma presentan un gradiente de profundidad menos abrupto que las que rodean a la isla Vernacci, las que, a su vez, están expuestas al oleaje del mar abierto. Sin embargo, en ambas colonias las aguas costeras (<2 km) alcanzan profundidades máximas inferiores a 20 m (cartas náuticas H-215 y H-264, Servicio de Hidrografía Naval de la República Argentina; F Quintana, datos no publ.).
Colocación de instrumentos
Los trabajos de campo se realizaron durante las primeras dos semanas del período de crianza de pichones, en diciembre de 1996 y 1997 (Punta Loma) y en diciembre de 1998 (Isla Vernacci). Durante los tres años se estudiaron 18 aves adultas (9 en cada colonia). Cada ave fue equipada con un transmisor VHF cuyo peso promedio fue de 23.6 g (DE = 1.5, n = 11), lo que representó menos del 1.6% del peso corporal de los animales. Los instrumentos fueron fijados a las plumas del dorso de las aves con resina epoxi (Evcon 5min®) y precintos plásticos.
Patrón de alimentación y comportamiento de buceo
El comportamiento de alimentación de las aves portadoras de instrumentos se determinó a través de observación directa de nidos y técnicas de radiotelemetría. En Punta Loma, el seguimiento visual de las aves se realizó entre las 04:00 h y las 22:30 h desde puestos de observación ubicados sobre los acantilados, con telescopios 20× y binoculares 10×35. Una vez que abandonaban la colonia las aves fueron monitoreadas desde dos puestos fijos de recepción equipados con antenas Yagi 9 elementos y un equipo radiorreceptor con barrido de frecuencias (Advanced Telemetry Systems, Bethel, Minnesota). A partir de cambios en la intensidad y otras características de la señal fue posible identificar cuándo las aves estaban en la colonia, volando, sobre la superficie del mar o debajo del agua (ver detalles en Wanless & Harris 1992). De esta forma fue posible determinar el tiempo de vuelo hacia y desde las áreas de alimentación, así como el inicio y fin de cada uno de los turnos de buceo. Así, para la colonia de Punta Loma se obtuvo información sobre hora de salida y arribo al nido e inicio y fin de cada uno de los turnos de buceo.
El comportamiento de alimentación de los cormoranes de la isla Vernacci se determinó exclusivamente mediante técnicas de telemetría. Para ello se utilizó un colector automático de datos (Advanced Telemetry Systems, Bethel, Minnesota) ubicado a unos pocos metros de los nidos monitoreados y, como en el caso de Punta Loma, estaciones receptoras fijas en tierra con antenas Yagi 9 elementos. De esta forma fue posible registrar la presencia o ausencia en el nido de cada uno de los animales portadores de instrumentos durante las 24 horas del día. El colector automático fue programado para realizar un barrido de muestreo cada diez minutos de todas las frecuencias de radio utilizadas. A partir de un reloj interno del receptor fue posible estimar la hora de salida y arribo al nido de todas las aves estudiadas.
Para este trabajo se definió como área de alimentación a la posición donde un ave buceaba repetidamente. La localización de dichas áreas se determinó mediante el método de triangulación, a partir de las direcciones provenientes de las señales emitidas por los radiotransmisores. Al menos dos direcciones fueron tomadas en forma simultánea desde los puestos de recepción equipados con antenas Yagi (ver arriba). Las áreas de alimentación se obtuvieron como la intersección de los dosángulos determinados por la dirección de las señales, con un error de 5–20°. Dado que en Punta Loma su obtuvieron los tiempos de vuelo hacia y desde las áreas de alimentación (ver arriba), fue posible corregir los límites de lasáreas obtenidas por triangulación. Dicha corrección se realizó sobre la base de las distancias estimadas a las áreas de alimentación (±0.5 km), obtenidas a partir de una velocidad de vuelo promedio de 52 km/h (Pennycuick 1987) y los tiempos de vuelo observados (observaciones previas indicaron que las aves volaban en línea recta desde la colonia a lasáreas de alimentación). Para Isla Vernacci, las distancias a las áreas de alimentación correspondieron a la distancia promedio estimada entre la colonia y todos los vértices del polígono determinado por triangulación (área de alimentación). Mediante la técnica utilizada fue posible monitorear a todas las aves portadoras de instrumentos durante su permanencia en el mar y determinar la localización de sus áreas de alimentación. Ninguno de los cormoranes estudiados se alimentó fuera del rango de recepción de la señal (aprox. 10 km).
Debido a que la señal de radio se interrumpe durante las inmersiones y se restaura cuando los cormoranes se encuentran en superficie al emerger luego de cada buceo (Wanless et al. 1991), fue posible identificar una secuencia de buceos como una serie de cortes regulares de la señal (ver Wanless et al. 1991; Quintana 1999). Durante el seguimiento de cada animal se determinó así el inicio y el final de la actividad de buceo, la duración de cada uno de los buceos (DB) y los intervalos de recuperación en superficie entre inmersiones consecutivas (IS). DB e IS se determinaron mediante cronómetros con precisión de un segundo a partir de las interrupciones de la señal emitida por los radiotransmisores. En ambas colonias, la profundidad del mar en las áreas utilizadas por los cormoranes fue determinada por medio de cartas náuticas y registros propios (cartas H264 y C13, Servicio de Hidrografía Naval de la República Argentina; F Quintana, datos no publ.).
Análisis de datos
Para cada una de las variables estudiadas se determinó en cada año la existencia de diferencias significativas entre individuos por medio de la prueba de ANOVA o la de Kruskal–Wallis (P < 0.05). Los datos fueron agrupados solo en los casos en que no se encontraron diferencias significativas entre individuos. En caso contrario, se promediaron los promedios para cada variable. Hurlbert (1984) mostró que el empleo de observaciones dependientes es válido en la medida que las réplicas sean agrupadas para estimar un valor promedio. Los datos correspondientes a las temporadas 1996 y 1997 de la colonia de Punta Loma fueron tratados como el mismo conjunto de datos por no haberse encontrado diferencias significativas entre años (Prueba de Mann–Whitney, P > 0.05).
RESULTADOS
Comportamiento de buceo
En ambas colonias, el patrón general de buceo consistió en una serie ininterrumpida de inmersiones prolongadas intercaladas con breves intervalos de recuperación en superficie. Todos los viajes de alimentación estuvieron constituidos por una única secuencia o turno de buceo. En general, las inmersiones fueron de mayor duración que los IS a lo largo de toda la secuencia de inmersiones en los turnos de buceo de todas las aves estudiadas. Tanto DB como IS se mantuvieron relativamente constantes a lo largo de la secuencia de buceos realizados en cada viaje de alimentación.
Los cormoranes de Punta Loma realizaron buceos de duración similar a la observada en las aves de Isla Vernacci (U = 29, P > 0.05; Prueba de Mann-Whitney). La duración promedio de los buceos realizados por los cormoranes fue 47 seg (n = 9 aves) y 50 seg (n = 8 aves) para Punta Loma e Isla Vernacci, respectivamente (Tabla 1). Asimismo, el tiempo de recuperación en superficie fue similar en ambas colonias (U = 20, P > 0.05; Tabla 1). En consecuencia, durante los turnos de buceo, los cormoranes de ambas colonias permanecieron la misma proporción de tiempo debajo del agua (U = 25, P > 0.05) y sus tasas de buceo no mostraron diferencias significativas (U = 25, P > 0.05). La eficiencia de buceo, definida como el tiempo de permanencia debajo del agua en función del tiempo en superficie (Dewar 1924), fue alta en las dos colonias estudiadas (Tabla 1). La máxima eficiencia de buceo alcanzada durante un turno de buceo fue de 8.6 y 7.4 para Punta Loma e Isla Vernacci, respectivamente.
Tabla 1. Valores promedio (± DE cuando corresponde) de las principales variables del comportamiento de buceo del Cormorán Cuello Negro en las colonias de Punta Loma e Isla Vernacci.
Table 1. Mean values (± SD when applicable) for diving behavior variables of the Rock Shag at Punta Loma and Isla Vernacci colonies.

Los cormoranes de Punta Loma presentaron en los dos años de estudio una correlación muy baja entre las DB y los IS posteriores a cada buceo (r = 0.06, P < 0.05 y r= 0.04, P < 0.05, para 1996 y 1997, respectivamente). En ambas temporadas, el tiempo de recuperación en superficie se incrementó en forma lineal con la duración del buceo previo. Sin embargo, las pendientes de las rectas obtenidas para ambos años fueron extremadamente bajas (b = 0.044 y b = 0.039 para 1996 y 1997, respectivamente), aunque significativamente distintas de cero (t4769= 4.7, P < 0.01 y t6173= 4.8, P < 0.01), probablemente como consecuencia del gran tamaño de muestra. La correlación entre DB e IS fue también baja para los cormoranes de Isla Vernacci (r = 0.33). Asimismo, la pendiente de la regresión entre ambas variables fue baja (b = 0.4) pero significativamente distinta de cero (t1369= 12.9, P < 0.01). Si bien en las dos colonias las pendientes de las regresiones obtenidas fueron muy bajas, la dependencia entre ambas variables fue mayor para los cormoranes de Isla Vernacci (t2,12309= 1.97, P < 0.05). Además, los IS resultaron independientes de DB solo en el 37% de los cormoranes de Isla Vernacci, frente al 56% de los cormoranes de Punta Loma.
Patrón de alimentación
Comportamiento de alimentación.– En líneas generales, el patrón de alimentación (comportamiento y áreas de alimentación) del Cormorán Cuello Negro presentó características similares en las dos colonias estudiadas. En ambos sitios, las aves portadoras de instrumentos alternaron con sus parejas períodos de atención del nido con períodos de alimentación en el mar. Luego de abandonar el nido, todas las aves se dirigieron directamente a las áreas de alimentación donde iniciaron una serie ininterrumpida de buceos, luego de la cual regresaron directamente a la colonia.
Las proporciones de tiempo (total horas luz) invertido por las aves en las actividades de permanencia en el nido (alimentación y cuidado de pichones) y de alimentación en el mar fueron similares entre ambas colonias (U = 18, P > 0.05; Prueba de Mann-Whitney; Tabla 2). En Punta Loma, los cormoranes realizaron en promedio 138 buceos por viaje de alimentación (n = 8 aves) y el número máximo registrado por viaje fue de 546 buceos (Tabla 2). Por su parte, los cormoranes de Isla Vernacci realizaron 78 buceos por viaje (n = 5 aves) (Tabla 2). Las diferencias entre colonias no resultaron significativas (U = 31, P > 0.05). En ambas colonias, la duración de los viajes de alimentación se incrementó significativamente con el número de buceos por viaje (r = 0.9, n = 69, P < 0.01 para Punta Loma; r = 0.7, n = 24, P < 0.01 para Isla Vernacci; Correlación de Spearman; Figura 2).
Tabla 2. Valores promedio (± DE) de las variables asociadas al patrón de alimentación del Cormorán Cuello Negro en las colonias de Punta Loma e Isla Vernacci. El asterisco indica diferencias significativas entre colonias (P < 0.05).
Table 2. Mean values (± SD) for feeding pattern variables of the Rock Shag at Punta Loma and Isla Vernacci colonies. Asterisk denotes significant differences between colonies (P < 0.05).
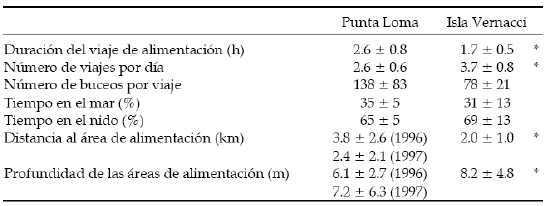

Figura 2. Relación entre el número de buceos por viaje y la duración de los viajes de alimentación del Cormorán Cuello Negro en las colonias de Punta Loma (arriba) e Isla Vernacci (abajo). Para Punta Loma: y = 0.87 + 0.011×x, P < 0.01; para Isla Vernacci: y = 0.39 + 0.015×x, P < 0.01.
Figure 2. Relationship between the number of dives per trip and the duration of the foraging trip performed by the Rock Shag at Punta Loma (above) and Isla Vernacci (below) colonies. For Punta Loma: y = 0.87 + 0.011×x, P < 0.01; for Isla Vernacci: y = 0.39 + 0.015×x, P < 0.01.
Durante las dos primeras semanas del período de crianza de pichones, los cormoranes de Isla Vernacci realizaron, en promedio, un viaje de alimentación más por día que los cormoranes de Punta Loma (3.7 vs. 2.6, n = 7 y n = 3 aves, respectivamente; U = 20, P < 0.05, Prueba de Mann-Whitney; Tabla 2). En ambas colonias, el número de viajes diarios se correlacionó en forma negativa con la duración promedio diaria de los mismos (r = -0.8, n = 36, P < 0.05 para Punta Loma; r = -0.7, n = 19, P < 0.01 para Isla Vernacci; Correlación de Spearman). Los viajes realizados por los cormoranes de Punta Loma fueron más prolongados que los llevados a cabo por las aves de Isla Vernacci (2.6 vs. 1.7 horas, n = 8 aves en cada colonia; U = 54, P < 0.05; Tabla 2). El 56% de los viajes realizados por los cormoranes de Punta Loma fueron superiores a dos horas, mientras que solo el 21% de los de Isla Vernacci resultó superior a dicho valor.
Áreas de alimentación.– Las aves de ambas colonias se alimentaron en aguas costeras ubicadas a menos de 5 km de las colonias, explotaron un solo parche durante cada viaje de alimentación y fueron consistentes en el uso de sectores particulares (Figura 3). Así, los cormoranes de Punta Loma utilizaron principalmente (92% de los casos) aguas ubicadas al oeste de la colonia (Figura 3a y 3b), mientras que los de Isla Vernacci utilizaron exclusivamente áreas ubicadas en la boca de la caleta Malaspina (Figura 3c).


Figura 3. Localización de las áreas de alimentación del Cormorán Cuello Negro de la colonia de Punta Loma en 1996 (a), en 1997 (b), y de la colonia de Isla Vernacci en 1998 (c), determinadas por radiotelemetría.
Figure 3. Location of feeding areas of the Rock Shag at Punta Loma colony during 1996 (a), during 1997 (b), and at Isla Vernacci colony during 1998 (c), as assessed by radio-telemetry.
En líneas generales, los cormoranes de Isla Vernacci recorrieron, en promedio, menos distancia entre la colonia y las áreas de alimentación que los de Punta Loma (Tabla 2). Sin embargo, estas diferencias resultaron significativas solo para las distancias de Punta Loma en 1996 y las de Isla Vernacci en 1998 (Z = 4.42, P < 0.001; Prueba de Mann-Whitney). Durante 1996, el 64% de los viajes de alimentación realizados en Punta Loma fueron de menos de 4 km, mientras que la mayoría de los viajes realizados por los cormoranes de Isla Vernacci (82%) y por los de Punta Loma en 1997 (83%) tuvo una distancia inferior. En ninguna de las dos colonias la distancia al área de alimentación fue un buen indicador de la duración de los viajes (r = 0.4, n = 88, P < 0.01 para Punta Loma; r = 0.4, n = 22, P > 0.05 para Isla Vernacci; Correlación de Spearman).
En general, las áreas de alimentación utilizadas por las aves no superaron los 10 m de profundidad (77 vs. 96% de los casos para Punta Loma e Isla Vernacci, respectivamente). De acuerdo a la batimetría de las áreas de estudio (carta náutica H264; F Quintana, datos no publ.), la profundidad promedio de las áreas de alimentación fue de 6.1 m, 7.2 m y 8.2 m para 1996, 1997 y 1998, respectivamente (Tabla 2).
DISCUSIÓN
Comportamiento de buceo
Estudios comparados del patrón de buceo en aves marinas sugieren que distintas especies pueden administrar su tiempo debajo del agua de diferentes maneras, de acuerdo a su ecología de alimentación (Ydenberg 1988). Wilson & Wilson (1988) sugieren que el comportamiento de alimentación está fuertemente condicionado por las características del área de alimentación. Así, la duración de los buceos resulta usualmente proporcional a la profundidad a la que los individuos se alimentan y podría esperarse que otros factores, tales como la topografía del fondo, la turbidez del agua, el tipo y la disponibilidad de presas, también influencien el tiempo de buceo. Los resultados de este trabajo muestran una marcada similitud entre colonias respecto a los parámetros de buceo estudiados. Dicha similitud podría ser atribuible a semejanzas en las características del ambiente que tienen efectos sobre el comportamiento de buceo. La escasa diferencia de profundidad entre las aguas que rodean a ambas colonias podría explicar la similitud encontrada en la DB y el IS. Lamentablemente, otras características físicas y biológicas de las áreas de alimentación (tipo de substrato, distribución, abundancia y tipo de alimento) no fueron determinadas durante este trabajo. Sin embargo, datos provenientes de otras investigaciones indican que las áreas de alimentación utilizadas por las aves de ambas colonias presentan sustratos constituidos principalmente por restingas rocosas, rodados, arena y fango (Mouzo et al. 1978; Herrera 1997; Lanfiutti 2000; Quintana 2001) y que la dieta del Cormorán Cuello Negro (constituida principalmente por peces bentónicos de la familia Nototheniidae y especies tales como Ribeiroclinus eigenmanni y Triathalassothia argentina) es similar en ambas colonias (Punta et al. 1993; Malacalza et al. 1997). En ambas colonias los cormoranes parecen seleccionar áreas de alimentación de características ambientales similares con respecto a la profundidad, el tipo de fondo y la disponibilidad de presas.
Tanto en Punta Loma como en Isla Vernacci, los cormoranes presentaron períodos de buceo más prolongados que los predichos por Cooper (1986) sobre la base de la relación entre la DB y la masa corporal. En este estudio, la DB resultó el doble de la esperada para un cormorán de 1.5 kg. En contraposición a lo descripto por Cooper (1986) y otros autores para varias especies de cormoranes (Croxall et al. 1991; Wanless & Harris 1991), en ninguna de las dos colonias se encontró una correlación alta entre la DB y los IS posteriores. Esta baja dependencia podría ser el resultado del buceo en aguas poco profundas, que resultó en una baja exigencia fisiológica para el buceo. Wanless et al. (1993) sugieren que el costo energético del buceo es superior cuando los cormoranes se alimentan en aguas profundas. En ese caso, luego de un buceo extenso el intervalo de recuperación en superficie debería incrementarse. Los cormoranes de ambas colonias se alimentaron esencialmente en aguas poco profundas, donde el esfuerzo de buceo sería menor y las largas inmersiones no necesariamente requerirían tiempos prolongados en superficie para recuperar las reservas de oxígeno. De esta forma, la tendencia observada respecto a la realización de buceos más profundos por parte de los cormoranes de Isla Vernacci se corresponde con una mayor correlación entre la DB y el IS registrados en esa colonia.
La eficiencia de buceo (tiempo sumergido/ tiempo en superficie) observada tanto en los cormoranes de Punta Loma como en los de Isla Vernacci fue alta comparada con la reportada para otras especies de cormoranes, tales como Phalacrocorax atriceps, Ph. aristotelis y Ph. capillatus, cuyas eficiencias de buceo fueron menores a 1 (Croxall et al. 1991; Kato et al. 1992; Wanless et al. 1993; Watanuki et al. 1996; ver revisión en Cooper 1986). Las aves pasaron entre el 74 y el 76% del turno de buceo debajo del agua. A una profundidad determinada, el incremento del tiempo debajo del agua durante un ciclo de buceo permite maximizar el tiempo en el fondo disponible para búsqueda y captura de presas (Kooyman & Davis 1987). Las aves estudiadas bucearon en aguas someras, donde el tiempo de tránsito hacia y desde el fondo debería haber sido bajo, por lo que es razonable pensar que en los cormoranes de ambas colonias el tiempo de búsqueda en el fondo representó casi la totalidad del tiempo de buceo. Si bien este comportamiento podría resultar ventajoso ante cualquier circunstancia, podría serlo aún más en condiciones de escasez de alimento (baja abundancia o disponibilidad) o de bajo rendimiento energético de las presas disponibles (ver abajo).
Patrón de alimentación
En términos generales, el patrón de alimentación observado fue similar en ambas colonias. Durante las primeras dos semanas del período de crianza de pichones, los cormoranes alternaron diariamente viajes de alimentación en el mar con períodos de permanencia en el nido para alimentar y cuidar a los pichones. En cada viaje de alimentación las aves volaron directamente a los parches de alimentación, donde iniciaron una serie ininterrumpida de buceos, luego de la cual regresaron nuevamente al nido. Durante estos viajes, todos los individuos recorrieron distancias cortas (<5 km) para alcanzar las áreas de alimentación y fueron marcadamente recurrentes en el uso de áreas específicas.
Los únicos datos publicados acerca de la distribución y características de las áreas de alimentación del Cormorán Cuello Negro en Patagonia corresponden a los registros visuales realizados por Punta et al. (1993), que son similares a los obtenidos en este estudio. Los resultados presentados en este trabajo confirman el uso de aguas costeras poco profundas por parte del Cormorán Cuello Negro comoáreas de alimentación. Solo durante 1996 los cormoranes de Punta Loma recorrieron distancias mayores entre la colonia y las áreas de alimentación y se alimentaron en aguas ligeramente menos profundas. Sin embargo, al momento de sacar conclusiones, estas diferencias deben ser tomadas con cautela debido a que ambas variables fueron estimadas por métodos indirectos.
Los resultados indican que, tanto en Punta Loma como en Isla Vernacci, las aves bucearon la mayor parte del tiempo que estuvieron fuera de la colonia (aprox. 90%), realizaron varios (2.6–3.7) y prolongados (1.7–2.6 h) viajes de alimentación diarios, y llevaron a cabo un alto número de buceos por viaje (78–138). Estos resultados sugieren que, en ambas colonias, los cormoranes dedicaron una alta proporción de su tiempo en el mar para obtener alimento para sí mismos y sus pichones. El número de viajes diarios de alimentación observado resultó similar a los reportados para otras especies del género tales como Phalacrocorax aristotelis (Wanless & Harris 1992), P. georgianus (Wanless et al. 1992) y P. nivalis (Green & Williams 1997). Aunque la duración de los viajes de alimentación realizados en las dos colonias también se encontró dentro del rango observado para las especies mencionadas, todos los cormoranes estudiados realizaron viajes de alimentación prolongados, superiores a las 2 h (máx = 5.6 h). Para las dos colonias la duración del viaje de alimentación fue un buen indicador del número de buceos realizados por viaje. De esta forma, dado el costo energético implícito en el buceo de los cormoranes (Wanless et al. 1993), la extensión del viaje podría interpretarse como un buen indicador del esfuerzo de alimentación.
Una de las principales diferencias encontradas al comparar el patrón de alimentación del Cormorán Cuello Negro con el de otras especies de cormoranes fue el alto número de buceos realizados por día. Considerando un promedio de 138 buceos por viaje y un valor promedio de 2.6 viajes por día, en Punta Loma los cormoranes realizarían, en promedio, 360 buceos por día. La estimación para Isla Vernacci indica 280 buceos diarios. Si bien la comparación con otras especies de cormoranes debe ser realizada con precaución dadas las diferencias en las profundidades de buceo, el número de buceos diarios registrado para el Cormorán Cuello Negro es marcadamente superior al de otras especies. Wanless et al. (1992) reportaron un promedio de 107 buceos por día en Phalacrocorax georgianus, mientras que Green & Williams (1997) registraron 61 buceos diarios para los cormoranes de Heard Island. Por su parte, Nehls & Gienapp (1997) reportaron un máximo de 160 buceos por viaje para P. carbo carbo en el mar de Wadden.
Viajes largos de alimentación con un alto número de buceos estuvieron relacionados a una reducida disponibilidad de presas en otras aves y mamíferos marinos. Monaghan et al. (1994) encontraron que durante los años de menor disponibilidad de presas, los araos comunes (Uria aalge) realizaron viajes de alimentación más prolongados y mayor número de buceos por día. Resultados similares han sido reportados en dos especies de pinnípedos: el Lobo Fino Antártico (Arctocephalus gazella) (Costa et al. 1989) y el Lobo Marino Californiano (Zalophus californianus) (Feldkamp et al. 1989). De esta forma, los viajes de alimentación prolongados y el elevado número de buceos diarios registrados para los cormoranes en ambas colonias podrían ser interpretados como una necesidad de satisfacer los requerimientos energéticos diarios como consecuencia de una baja tasa de captura producida por una reducida disponibilidad de presas (distribución, abundancia, accesibilidad). Además, las eficiencias de buceo registradas en ambas colonias son altas comparadas con las reportadas para la mayoría de las especies de cormoranes (ver arriba) y, en esta especie, muestran una clara optimización del tiempo de permanencia en el fondo durante las inmersiones (ver Quintana 1999). En este sentido, Wanless et al. (1998) realizaron estudios sobre cormoranes europeos y encontraron altas eficiencias de buceo asociadas a bajas tasas de captura de presas en aguas con baja concentración de alimento.
Además de la disponibilidad de alimento, el bajo rendimiento energético de las presas consumidas podría ser otro de los factores responsables del alto porcentaje de tiempo invertido por estas aves en la alimentación durante su permanencia en el mar. La dieta del Cormorán Cuello Negro de Punta Loma y de áreas adyacentes a Isla Vernacci está constituida principalmente por peces bentónicos e invertebrados de pequeña talla y bajo valor energético (Punta et al. 1993; González 1995; Malacalza et al. 1997). Si bien no existe información sobre distribución, abundancia y hábitos de las especies presa, el patrón de alimentación y buceo observado en las colonias estudiadas sugiere que los cormoranes explotan fuentes de alimento restringidas, distribuidas en parches discretos y/o de baja accesibilidad. Si este supuesto es cierto, podría constituir una de las razones para entender el reducido tamaño de las colonias de esta especie en todo su rango de distribución a lo largo de la costa patagónica.
AGRADECIMIENTOS
Este trabajo se realizó mediante subsidios de investigación aportados por Ecocentro Puerto Madryn, Wildlife Conservation Society, CONICET y ANPCyT. Agradecemos a D Posdena, V Massara y V Osorio por la asistencia durante los trabajos de campo, a la Lic. MR Marín por su asistencia profesional en el diseño de la base de datos y la edición final de los figuras, a la Lic. A Sapoznikow por sus comentarios sobre versiones preliminares del manuscrito, al Organismo Provincial de Turismo y la Dirección Provincial de Fauna Silvestre, Chubut, por facilitar los permisos para trabajar en Punta Loma y caleta Malaspina y al Centro Nacional Patagónico por el apoyo institucional.
BIBLIOGRAFÍA
ASHMOLE, NP. 1971. Seabird ecology and the marine environment. Pp. 224–271 en: DS Farner & JR King (eds). Avian biology. Academic Press, New York. [ Links ]
COOPER, J. 1986. Diving patterns of cormorants Phalacroracidae. Ibis 114:360–366. [ Links ]
COSTA, D; JO CROXALL & CD DUCK. 1989. Foraging energetics of Antarctic fur seals in relation to changes in prey availability. Ecology 70:596–606. [ Links ]
CROXALL, JP; Y NAITO; A KATO; P ROTHERY & DR BRIGGS. 1991. Diving patterns and performance in the Antarctic Blue-eyed shag, Phalacrocorax atriceps. J. Zool. 225:177–199. [ Links ]
DEWAR, JM. 1924. The bird as a diver. Witherby, Londres. [ Links ]
FELDKAMP, SD; RL DELONG & GA ANTONELIS. 1989. Diving patterns of California sea lions, Zalophus californianus. Can. J. Zool. 67:872–883. [ Links ]
GONZÁLEZ, ML. 1995. Calidad de la dieta del cormorán real, Phalacrocorax albiventer, durante el ciclo reproductivo en la colonia de punta León. Seminario de Licenciatura, Universidad Nacional de la Patagonia San Juan Bosco. Puerto Madryn, Argentina. [ Links ]
GREEN, K & R WILLIAMS. 1997. Biology of the Heard Island Shag Phalacrocorax nivalis. 3. Foraging, diet and diving behaviour. Emu 97:76–83. [ Links ]
HARRISON, P. 1983. Seabirds. An identification guide. Hougthon Mifflin Company, New York. [ Links ]
HERRERA, GO. 1997. Dieta reproductiva de la gaviota de olrog, Larus atlanticus, en la provincia del Chubut. Seminario de Licenciatura, Universidad Nacional de la Patagonia San Juan Bosco. Trelew, Argentina. [ Links ]
HUNTLEY, ME; MDG LOEZ & DM KARL. 1991. Top predators in the Southern Ocean: a major leak in the biological carbon pump. Science 253:64–66. [ Links ]
HURLBERT, SH. 1984. Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54:187–211. [ Links ]
JOHNSGARD, PA. 1993. Cormorants, darters and pelicans of the world. Smithsonian Institution Press, Washington D.C. [ Links ]
KATO, A; JP CROXALL; Y WATANUKI & Y NAITO. 1992. Diving patterns and performance in male and female Blue-eyed Cormorants, Phalacrocorax atriceps at South Georgia. Mar. Ornithol. 19:117–129. [ Links ]
KOOYMAN, GL. 1989. Diverse divers. Springer-Verlag, Berlín, Alemania. [ Links ]
KOOYMAN, GL & RW DAVIS. 1987. Diving behaviour and performance with special reference to penguins. Pp. 63–75 en: JP Croxall (ed.). Seabirds: feeding ecology and role in marine ecosystems. Cambridge University Press, Cambridge. [ Links ]
LANFIUTTI, M. 2000. Distribución, abundancia y hábitat de la ballena franca austral, Eubalaena australis, en la península Valdés. Seminario de Licenciatura, Universidad Nacional de la Patagonia San Juan Bosco. Puerto Madryn, Argentina. [ Links ]
MALACALZA, V; M BERTELOTTI & T PORETTI. 1997. Variación estacional de la dieta de Phalacrocorax magellanicus (Aves: Phalacrocoracidae) en Punta Loma (Chubut, Argentina). Neotropica 43:35–37. [ Links ]
MONAGHAN, P; P WALTON; S WANLESS; JD UTTLEY & MD BURNS. 1994. Effects of prey abundance on the foraging behaviour, diving efficiency and time allocation of the breeding guillemots Uria aalgae. Ibis 136:214–222. [ Links ]
MONTEVECCHI, WA. 1993. Birds as indicators of changes in marine prey stocks. Pp. 217–266 en: RW Furness & JJD Greenwood (eds). Birds as monitors of environmental change. Chapman & Hall, London. [ Links ]
MOUZO, FG; ML GARZA; JF IZQUIERDO & RO ZIBECCHI. 1978. Rasgos de la geología submarina del golfo Nuevo (Chubut). Acta Oceanográfica 2:69–91. [ Links ]
NEHLS, G & P GIENAPP. 1997. Prey selection and foraging behaviour of cormorants Phalacrocorax carbo in the Wadden sea. Vogelwelt 118:33–40. [ Links ]
PENNYCUICK, CJ. 1987. Flight of auks (Alcidae) and other northern seabirds compared with southern procellariforms. J. Exp. Biol. 128:335–347. [ Links ]
PUNTA, GE; JRC SARAVIA & PM YORIO. 1993. The diet and foraging behaviour of two patagonian cormorants. Mar. Ornithol. 21:27–36. [ Links ]
QUINTANA, F. 1999. Diving behavior of Rock Shag at a Patagonian colony of Argentina. Waterbirds 22:466–471. [ Links ]
QUINTANA, F. 2001. Foraging behaviour and feeding locations of Rock Shags Phalacrocorax magellanicus from a colony in Patagonia, Argentina. Ibis 143:547–553. [ Links ]
WANLESS, S; T CORFIELD; MP HARRIS; ST BUCKLAND & A MORRIS. 1993. Diving behaviour of the shag Phalacrocorax aristotelis (Aves: Pelecaniformes) in relation to water depth and prey size. J. Zool. 231:11–25. [ Links ]
WANLESS, S; D GREMILLET & MP HARRIS. 1998. Foraging activity and performance of Shags Phalacrocorax aristotelis in relation to environmental characteristics. J. Avian Biol. 29:49–54. [ Links ]
WANLESS, S & MP HARRIS. 1991. Diving patterns of full-grown and juvenile Rock shags. Condor 93:44–48. [ Links ]
WANLESS, S & MP HARRIS. 1992. At-sea activity budgets of a pursuit-diving seabird monitored by radiotelemetry. Pp. 591–598 en: JG Priede & SM Swift (eds). Wildlife telemetry: remote monitoring and tracking of animals. Ellis Horwood, New York y London. [ Links ]
WANLESS, S; MP HARRIS & JA MORRIS. 1991. Foraging range and feeding locations of Shags Phalacrocorax aristotelis during chick rearing. Ibis 133:30–36. [ Links ]
WANLESS, S; MP HARRIS & JA MORRIS. 1992. Diving behaviour and diet of the Blue-eyed shag at South Georgia. Polar Biol. 12:713–719. [ Links ]
WATANUKI, Y; A KATO & Y NAITO. 1996. Diving performance of male and female Japanese Cormorants. Can. J. Zool. 74:1098–1109. [ Links ]
WILSON, RP & MPT WILSON. 1988. Foraging behaviour in four sympatric cormorants. J. Anim. Ecol. 57:943–955. [ Links ]
YDENBERG, RC. 1988. Foraging by diving birds. Proceedings of the International Ornithological Congress 19:1832–1842. [ Links ]
YORIO, P. 2000. Breeding seabirds of Argentina: conservation tools for a more integrated and regional approach. Emu 100:367–375. [ Links ]
YORIO, P; E FRERE; P GANDINI & G HARRIS. 1998. Atlas de la distribución reproductiva de aves marinas en el litoral patagónico argentino. Plan de manejo integrado de la zona costera patagónica. Fundación Patagonia Natural y Wildlife Conservation Society. Inst. Salesiano de Artes Gráficas. Buenos Aires, Argentina. [ Links ]

