










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.12 n.2 Córdoba jul./dic. 2002
TRABAJOS ORIGINALES
Papel del alga Nostoc commune y efecto del pastoreo por vacunos sobre la colonización de suelos alcalinos en la Pampa Deprimida
Oscar E Ansín1,*, Alejandro Deregibus2 & Jorge W Lanfranco1
1 Facultad de Ciencias Agrarias y Forestales, Universidad Nacional de La Plata, La Plata, Argentina
2 Facultad de Agronomía, Universidad de Buenos Aires, Buenos Aires, Argentina
* Facultad de Ciencias Agrarias y Forestales, Univ. Nac. de La Plata; CC 31; 1900 La Plata; Buenos Aires, Argentina. Email: osansi@ceres.agro.unlp.edu.ar
Recibido: 19 febrero 2002;
Revisado: 14 junio 2002;
Versión final: 18 julio 2002;
Aceptado: 22 julio 2002
RESUMEN. El alga Nostoc commune (Vauch.), una cianofícea fotótrofa y fijadora de nitrógeno, es conspicua en los suelos alcalinos de la Pampa Deprimida. Se evaluó la posible relación positiva entre la presencia de Nostoc commune y la germinación de semillas y la emergencia de plántulas. Se propone que la presencia del alga tendría un papel ecológico significativo en los procesos de colonización de los suelos alcalinos desnudos y que el pastoreo con vacunos afectaría dicho papel. Se observó que los micrositios cubiertos por el alga mostraron un mayor número de plántulas en otoño, invierno y primavera, y que la mayor abundancia de plántulas (68.3 plántulas/dm2) ocurrió en las microdepresiones con algas. La germinación en condiciones de invernáculo de semillas de Hordeum pusillum (Steud.), un pasto anual característico de estos ambientes alcalinos, fue mayor cuando se las colocó tanto arriba como por debajo de los biodermas algales. Bajo estas condiciones, el suelo fue menos alcalino y se incrementó su contenido de materia orgánica, carbono y nitrógeno. El pastoreo afectó la presencia del alga y la cobertura vegetal. Se concluye que la presencia de los biodermas algales tiene un significativo impacto ecológico al mejorar las propiedades del suelo, estimulando la germinación, la emergencia y el establecimiento de las plantas anuales tales como Hordeum pusillum. El manejo controlado del pastoreo favorecería la presencia de Nostoc commune, contribuyendo a la recuperación del hábitat.
Palabras clave: Colonización vegetal; Nostoc commune; Pastizales naturales; Pastoreo; Establecimiento de plántulas.
ABSTRACT. Role of the blue green alga Nostoc commune and effect of cattle grazing on plant colonization of alkaline soils of the Flooding Pampa: Nostoc commune (Vauch.), a blue green algae that influences soil fertility through atmospheric nitrogen fixation, is common in alkaline soils of the Flooding Pampa, Argentina. The objectives of this work were to evaluate the relationship between the presence of Nostoc commune, plant germination and seedling emergence, as a way to understand whether or not Nostoc commune plays a significant ecological role in the colonization process of bare alkaline soils. More seedlings were observed in micropatches covered with algae during autumn, winter and spring, being recorded maximum seedling abundance (68.3 seedlings/dm2) in microdepressions with algae. More seeds of Hordeum pusillum (Steud.), an annual grass conspicuous to these alkaline soils, germinated under greenhouse conditions when were placed either above or under algae Nostoc commune. The soil below the algae showed lower pH values and higher organic matter, carbon and nitrogen content. Cattle grazing had an impact on both algae and plant cover. We concluded that the presence of algal crusts has a significant ecological impact in these alkaline soils, as it improves soil properties and stimulates germination, seedling establishment and persistence of annual plants. Through controlled cattle grazing management is possible to promote the presence of Nostoc commune and enhance habitat quality.
Keywords: Plant colonization; Nostoc commune; Natural grasslands; Cattle grazing; Seedling establishment.
INTRODUCCIÓN
La composición florística de una comunidad vegetal es el resultado de procesos físicos, químicos y biológicos del ambiente que impactan en la estructura del pastizal (Tilman 1993). Son estos cambios ambientales, inducidos por procesos de origen natural o humano, los que hacen dinámica la sucesión vegetal y modifican la composición específica de la comunidad vegetal (Berendse 1985; Weiher & Keddy 1995). Predecir la dinámica sucesional de las comunidades vegetales con propósitos conservacionistas o utilitarios requiere entender la relación entre las especies que responden a múltiples condicionamientos ambientales (Insausti et al. 1999). El establecimiento de una especie en una determinada comunidad del pastizal es un proceso que depende de la existencia de semillas viables en el sitio, de la germinación de esas semillas y de que la plántula emergida sea capaz de crecer y alcanzar la madurez reproductiva. Para esto, la semilla y la plántula dependen de la capacidad del ambiente para satisfacer los requerimientos de la especie (Connell & Slatyer 1977).
La presencia del alga Nostoc commune (Vauch.), un microorganismo fotótrofo y fijador de nitrógeno, ha sido comunicada en trabajos fitosociológicos realizados en pastizales húmedos de Argentina (Vervoorst 1967; León et al. 1979), al igual que en ecosistemas similares del planeta (Zimmerman et al. 1980; Megharahj et al. 1993). Esta cianofícea vegeta en ambientes húmedos y cálidos, pudiendo tolerar inundaciones y sequías en áreas templadas (Dodds et al. 1995). La unión de un gran número de tricomas permite a estos organismos unicelulares desarrollar agregados o colonias (Mollenhauer et al. 1994). Se generan así biodermas algales (Halperin et al. 1976) que, cuando están secos, se asemejan a delgados y transparentes papeles negros (Hori et al. 1990), mientras que adoptan una textura gelatinosa cuando están húmedos y activos (Mollenhauer 1986). Los microorganismos fotótrofos desempeñan un importante papel ecológico (Brock 1973) en la promoción de procesos relacionados con la formación de agregados y la estabilización del suelo (Bailey et al. 1973; Malam Issa et al. 2001). Esta contribución es mayor cuando los microorganismos fijan el nitrógeno atmosférico (Rother & Whitton 1989), lo que también ocurre en áreas inundables (Rogers & Burns 1994).
Las comunidades vegetales de la Pampa Deprimida que se desarrollan sobre suelos hidromórficos y alcalinos, comúnmente llamados barros blancos, están dominadas por pastos duros conocidos como pelo de chancho (Distichlis spicata L., D. scoparia Kunth). En esas comunidades, Ansín (1995) encontró una relación positiva entre la presencia de Nostoc commune en otoño y la cobertura vegetal en la primavera siguiente, hecho que solo ocurría en los espacios de suelo desnudo, tanto en áreas pastoreadas como en áreas no pastoreadas. Este estudio se basó en la hipótesis que, en los pastizales de la Pampa Deprimida, Nostoc commune tendría un significativo papel ecológico al facilitar los procesos de colonización de los suelos alcalinos desnudos. Los objetivos de este trabajo fueron: 1) evaluar los efectos que la presencia de dicha alga tiene sobre las propiedades físico-químicas del suelo, la germinación de semillas y el establecimiento de monocotiledóneas y dicotiledóneas; y 2) evaluar si el pastoreo por vacunos condiciona la presencia del alga.
MÉTODOS
Área de estudio
La Pampa Deprimida, localizada al este de la provincia de Buenos Aires, abarca aproximadamente 60000 km2. Se la caracteriza como una extensa llanura con pendientes inferiores a 0.1%, con un clima templado, subhúmedo, con temperaturas promedio de 9 °C en invierno y de 22 °C en verano y con precipitaciones anuales que varían entre 850 y 1050 mm (Semmartin & Oesterheld 2001). Debido a su topografía plana y a la presencia de fuertes capas de arcilla entre los 15 y 60 cm de profundidad, es frecuente que estos suelos permanezcan inundados durante el invierno. Al no infiltrar el agua, en los períodos de sequías estivales los suelos se agrietan por la contracción de las arcillas.
El estudio se llevó a cabo en un establecimiento dedicado a la cría de vacunos ubicado cerca de la localidad de Verónica, al noreste de la Pampa Deprimida (35°30'S; 57°30'O). En un estudio previo realizado en el mismo establecimiento se determinó que el suelo ubicado en la parte más baja del paisaje corresponde principalmente al clasificado como Natracualf Típico, presentando un intrincado patrón de distribución con un Natracuol Típico (Alconada et al. 1993). Esto genera un mosaico de comunidades vegetales en las posiciones más deprimidas del paisaje, con áreas dominadas por Distichlis scoparia, Distichlis spicata, Sporobolus indicus (L.) y Sporobolus pyramidatus (Lam.), donde es conspicua la presencia de Nostoc commune. En zonas ubicadas topográficamente en posiciones más altas, dentro del área deprimida, prevalecen especies como Eryngium ebracteatum (Lam.), Danthonia montevidensis (Hack. et Arech.), Stenotaphrum secundatum (Walt.), Stipa charruana (Nees.) y Stipa papposa (Nees.). Cerca de los cursos de agua dominan Alternanthera philoxeroides (Mart.), Leersia hexandra (Sw.) y Ludwigia peploides (H.B.K.).
Presencia de plántulas
Se realizó un ensayo para evaluar la presencia de plántulas en áreas de suelo previamente cubiertas con Nostoc commune. Áreas en las cuales se desarrollaba la comunidad alcalina dominada por Distichlis spp. fueron sometidas a diferentes condiciones de pastoreo: a) sin pastoreo, b) pastoreo continuo y con carga alta (0.98 equivalentes vaca.ha-1.año-1), y c) pastoreo controlado y con carga moderada (0.54 equivalentes vaca.ha-1.año-1). El control del pastoreo determinaba ocupaciones y descansos de los potreros que variaron con la estación climática conforme a la velocidad de crecimiento de las plantas forrajeras y a las condiciones del suelo. Se controló la presión del pastoreo, utilizándose como especie clave a Sporobolus indicus, de manera que la biomasa aérea remanente tuviera una altura de aproximadamente 5 cm. Para cada tratamiento se eligieron cuatro situaciones que difirieron según la microtopografía del suelo y la presencia de algas (i.e., microdepresiones con o sin biodermas algales, microlomas con o sin biodermas algales). La presencia de plántulas en cada condición de pastoreo y situación de suelo se determinó en 10 estaciones permanentes circulares de 6.15 cm2 (Matteucci & Colma 1982), evitando los sitios con deyecciones de vacunos dado que las mismas pueden tener efecto sobre la productividad primaria (McNaughton et al. 1997). En el otoño de 1998 se realizaron censos florísticos para determinar la emergencia de plántulas y en la primavera siguiente se registró (en un solo recuento) el número de plántulas que habían logrado establecerse.
Establecimiento de la vegetación anual y efecto del pastoreo
En un área excluida al pastoreo con grandes herbívoros se registraron, desde el otoño hasta la primavera de 1999, el número de plántulas establecidas en las cuatro situaciones de suelo. Por cada tratamiento se ubicaron 16 estaciones de muestreo permanentes de 10×10 cm cada una. Simultáneamente, en cada estación climática se estimó la superficie cubierta por biodermas de Nostoc commune, por vegetación anual y por suelo desnudo. Se utilizó el método de interceptación lineal ideado por Canfield (Brown 1954). Para ello, en cada uno de los potreros correspondientes a las tres condiciones de pastoreo se seleccionaron áreas homogéneas de la comunidad alcalina donde se distribuyeron al azar cuatro líneas de 5 m de longitud cada una (Facelli et al. 1988), sobre las cuales se hicieron las lecturas para estimar la cobertura y la riqueza florística.
Variación en las propiedades edáficas
Se evaluó la existencia de posibles diferencias en las propiedades físico-químicas del suelo colonizado o no por el alga. Se consideró que esta información permitiría identificar algunas de las razones que determinan un comportamiento distinto de la germinación de semillas y de la supervivencia de plántulas en los sitios en que está presente el alga. Para ello, se extrajeron 40 muestras del horizonte superficial A (0–8 cm) de un suelo Natracualf ubicado fuera de las estaciones de muestreo y correspondientes a las situaciones de microlomas con y sin Nostoc commune. Las 20 muestras de cada situación se estabilizaron y homogeneizaron; previamente, en forma manual, se separaron los biodermas algales en los casos en que estaban presentes. Mediante métodos estándar (Bighams & Bartels 1996) se realizaron los siguientes análisis de laboratorio: humedad presente (determinada en estufa a 105 °C), pH (relación suelo/agua 1:2.5, determinación potenciométrica), carbono fácilmente oxidable (método de Walkley y Black), nitrógeno total (digestión húmeda, evaluación por el método de Microkjeldhal), relación carbono/nitrógeno, y materia orgánica (carbono x 1.72).
Germinación de semillas y emergencia de plántulas de Hordeum pusillum
Para investigar el efecto de Nostoc commune sobre la germinación de semillas y la emergencia de plántulas de Hordeum pusillum (una gramínea anual cuya presencia es frecuente en las situaciones descriptas), se llevó a cabo un experimento en invernáculo. A fines de la primavera de 1998 fueron cosechadas manualmente semillas maduras de esta gramínea en comunidades alcalinas del pastizal natural estudiado. Al día siguiente de cosechadas, las semillas se comenzaron a incubar en una cámara de germinación a una temperatura constante de 20 °C en oscuridad continua para evaluar su viabilidad. Las semillas se colocaron en cajas de Petri de vidrio transparente sobre papel de filtro y se mantuvieron húmedas con agua destilada. Se utilizaron 4 cajas con 40 semillas cada una. El número de semillas germinadas se registró diariamente durante 28 días (United States Department of Agriculture 1965; Davidovich Boberi 1980).
En el otoño de 1999, en las depresiones inundadas de las comunidades alcalinas estudiadas, se recolectaron colonias activas de Nostoc commune. Estas colonias fueron almacenadas y transportadas en frascos de vidrio transparente, abiertos y llenos de agua del mismo lugar donde fueron recogidas. Los biodermas algales fueron utilizados el mismo día en que fueron recolectados y dos semanas más tarde.
El experimento tuvo un diseño en bloques completamente al azar con ocho repeticiones y, tratando de reproducir las distintas condiciones de germinación de Hordeum pusillum observadas en el pastizal, cinco tratamientos: dos colocando semillas sobre a) suelo desnudo y b) bioderma algal, y tres cubriendo las semillas con c) una capa de bioderma algal, d) una capa de 1 cm de suelo, y e) una capa de suelo igual a la del tratamiento anterior y una capa de bioderma algal.
La experiencia se llevó a cabo en un invernáculo de la Facultad de Ciencias Agrarias y Forestales de la Universidad Nacional de La Plata. Se usaron 40 macetas de plástico, cada una de 6 cm de alto, 6 cm de diámetro superior, 4 cm de diámetro basal y un orificio de 2 mm en el centro de la base. Las macetas se llenaron con material extraído del horizonte superficial del suelo estudiado e inmediatamente fueron inundadas con agua destilada. Previo a ese riego, de acuerdo al tratamiento correspondiente, el suelo de las macetas fue cubierto con el alga. A partir de ese momento las macetas se ubicaron dentro de bandejas de plástico en un arreglo de 5×4. Las bandejas se mantuvieron con un volumen constante de agua destilada hasta 2 cm de altura desde sus bases. Dos semanas más tarde fueron sembradas ocho semillas de Hordeum pusillum en cada maceta de acuerdo con las condiciones establecidas en cada tratamiento, y fueron regadas con 2 cm3 de agua destilada. Los riegos se repitieron 24, 48 y 72 h más tarde. La densidad de siembra utilizada (1 semilla/3.5 cm2) tuvo por objeto evitar que las raíces se entrelacen y confundan los resultados (Davidovich Boberi 1980). Las observaciones sobre la germinación de las semillas y la emergencia de plántulas se realizaron diariamente hasta 60 días después de que comenzó el experimento.
Análisis estadístico
Los resultados fueron evaluados estadísticamente utilizando Análisis de Varianza. Los porcentajes fueron previamente transformados para cumplir con el supuesto de homogeneidad de varianza. La transformación utilizada fue arcsen √P/100, donde P es el porcentaje. Cuando se encontraron diferencias significativas (P < 0.01), los promedios de los valores fueron comparados mediante la Prueba de Tukey (Steel & Torrie 1990).
RESULTADOS Y DISCUSIÓN
Abundancia de plántulas
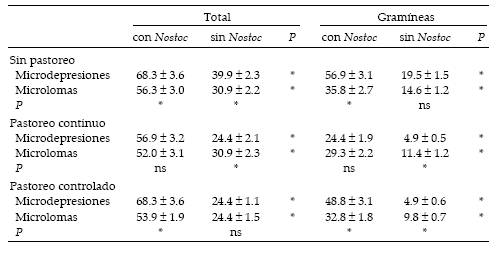
En la primavera fue evidente el predominio de las plántulas en aquellos sitios donde el suelo estaba cubierto con biodermas algales respecto a aquellos que carecían de los mismos (Tabla 1). Este hecho fue más frecuentemente observado en el caso de Hordeum pusillum. Si bien bajo pastoreo continuo ocurrió la pérdida de una estación de muestreo debido al pisoteo de los animales, la disminución de la cobertura del alga registrada en las áreas en esa condición de pastoreo no se asoció a dicha pérdida. Las diferencias fueron más notorias en las microdepresiones que en las microlomas, tanto en áreas pastoreadas como en aquellas excluidas al pastoreo, alcanzando en ambos casos un valor máximo de 68.3 plántulas/ dm2. El menor número de plántulas (4.9 gramíneas/dm2) se registró en las microdepresiones sin el alga en las dos áreas pastoreadas.
Tabla 1. Abundancia de plántulas (promedio ± DE) en cuatro situaciones de suelo (microdepresiones y microlomas con o sin biodermas algales) y tres condiciones de pastoreo durante la primavera en el noreste de la Pampa Deprimida. Los datos están expresados como número de plantas/dm2. También se muestran las diferencias estadísticas entre tratamientos. *: P < 0.01; ns: no significativo.
Table 1. Seedling abundance (mean ± SD) in four soil situations and three grazing conditions during spring in the north-eastern portion of the Flooding Pampa. Data are expressed as plant number/dm2. Statistical differences among treatments are also shown. *: P < 0.01; ns: not significant.
Variación en las propiedades edáficas
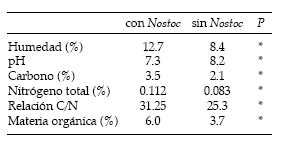
Las áreas en las que predominó Nostoc commune presentaron una mayor concentración de nitratos y nitritos, un mayor contenido de humedad, un pH cercano a la neutralidad, un aumento en el contenido de carbono, de nitrógeno y de materia orgánica, y un incremento de la relación C/N (Tabla 2). Estas condiciones crearían un ambiente favorable que habría determinado una mayor emergencia y supervivencia de plántulas donde los biodermas del alga se establecieron sobre la superficie del suelo.
Tabla 2. Propiedades físico-químicas de suelo cubierto o no con Nostoc commune en el noreste de la Pampa Deprimida. Se muestran las diferencias estadísticas entre ambas condiciones (*: P < 0.01).
Table 2. Properties of soils with or without Nostoc commune in the north-eastern portion of the Flooding Pampa. Statistical differences between both conditions are also shown (*: P < 0.01).
El incremento de nitrógeno y de materia orgánica observado en las áreas colonizadas coincide con los resultados de varios autores que trabajaron con algas fijadoras de nitrógeno (Fuller et al. 1960; Witty et al. 1979; Loftis & Kurts 1980; Zimmerman et al. 1980). Una vez fijado y liberado, el nitrógeno beneficia el crecimiento de las plantas vasculares (Maylad & McIntosh 1966; Rodgers et al. 1979; Dodds et al. 1995).
Establecimiento de la vegetación anual
El recuento de plántulas de otoño mostró que los micrositios cubiertos con Nostoc commune continuaban teniendo un mayor número de plántulas que los correspondientes a aquellos en donde el alga estuvo ausente (Figura 1). Es destacable que la mayoría de las plántulas (el 72%) emergió a través de los biodermas algales mientras que el resto lo hizo entre las colonias o en sus bordes.
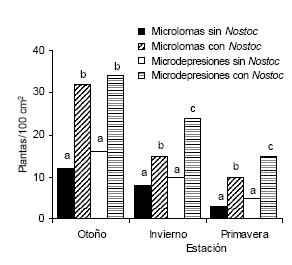
Figura 1. Abundancia de plántulas en cuatro situaciones de suelo (microdepresiones y microlomas con o sin biodermas algales) en tres estaciones del año en el noreste de la Pampa Deprimida. Dentro de una misma estación, letras diferentes indican diferencias significativas entre situaciones (P < 0.01).
Figure 1. Seedling abundance in four soil situations in three seasons in the north-eastern portion of the Flooding Pampa. Within a season, different letters indicate significant differences among situations (P < 0.01).
Si bien el número de plántulas disminuyó en invierno y en primavera, persistió una mayor abundancia de las mismas en los lugares originalmente cubiertos con Nostoc commune (Figura 1). En los censos de la primavera, Hordeum pusillum fue la más numerosa entre todas las gramíneas anuales que vegetaban en las cuatro situaciones de suelo.
Efecto del pastoreo sobre la cubierta vegetal
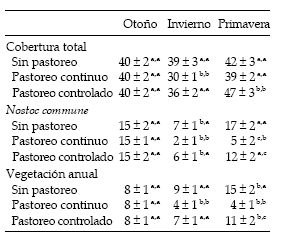
La condición de pastoreo afectó la presencia del alga y la cobertura vegetal (Tabla 3). Esto concuerda con lo afirmado por Alconada et al. (1993), quienes manifestaron que el pastoreo por vacunos ejerce un importante disturbio sobre los suelos y la vegetación de los pastizales de la Pampa Deprimida. Ello justificaría que tanto en la exclusión como en el pastoreo controlado las coberturas de Nostoc commune y de la vegetación anual fueran superiores a los correspondientes al pastoreo continuo. Una vez más surge la necesidad de desarrollar estrategias de pastoreo teniendo en cuenta que los efectos pueden ser positivos o negativos según el momento, intensidad y frecuencia con que la perturbación ocurra (Deregibus et al. 2001).
Tabla 3. Porcentaje (promedio ± DE) de cobertura total, de cobertura de Nostoc commune y de cobertura de vegetación anual bajo tres condiciones de pastoreo en tres estaciones del año en el noreste de la Pampa Deprimida. Letras diferentes indican diferencias significativas (P < 0.01); para cada par de letras, la primera de ellas señala diferencias entre estaciones del año (en una misma fila), mientras que la segunda señala diferencias entre condiciones de pastoreo (en una misma columna).
Table 3. Percentage (mean ± SD) of total cover, of Nostoc commune cover, and of annual plant cover under three grazing conditions in three seasons in the north-eastern portion of the Flooding Pampa. Different letters indicate significant differences (P < 0.01); for each pair of letters, the first one indicates differences among seasons (in a row), while the second letter indicates differences among grazing conditions (in a column).
Germinación de semillas y emergencia de plántulas de Hordeum pusillum
El porcentaje de semillas de Hordeum pusillum que germinaron tanto por encima como por debajo de los biodermas de Nostoc commune superaron el 80% (Tabla 4). Estos valores superan levemente al porcentaje de semillas germinadas obtenido en la cámara de germinación (75%) y significativamente a la germinación de las semillas ubicadas sobre el suelo desnudo. Las semillas cubiertas por una capa de suelo germinaron mucho menos. El efecto benéfico de la presencia de algas en el establecimiento de especies como maíz (Zea mays L.), arroz (Oriza sativa L.), trigo (Triticum aestivum L.) y lechuga (Lactuca sativa L.) fue señalado en los trabajos de Rother & Whitton (1989), Roger et al. (1993) y Svircev et al. (1997).
Tabla 4. Germinación (promedio ± DE) de semillas de Hordeum pusillum del noreste de la Pampa Deprimida expuestas a distintas condiciones. Letras diferentes indican diferencias significativas entre condiciones (P < 0.01).
Table 4. Germination (mean ± SD) of Hordeum pusillum seeds from the north-eastern portion of the Flooding Pampa exposed to different conditions. Different letters indicate significant differences among conditions (P < 0.01).

En cuanto al tiempo de germinación, las semillas ubicadas sobre el suelo desnudo lo hicieron tempranamente (en los primeros cuatro días); las últimas plántulas en emerger, 35 días luego de sembradas, fueron las cubiertas con una capa de suelo. A partir de dicho momento no se hicieron nuevos registros.
CONCLUSIONES Y ORIENTACIONES DE MANEJO
Los resultados de esta investigación indican que Nostoc commune desempeñaría un papel ecológico significativo en los procesos de colonización de los suelos desnudos en las comunidades vegetales de los pastizales hidromórficos y alcalinos de la zona estudiada. La germinación de semillas, la emergencia de plántulas y su establecimiento en los suelos alcalinos serían favorecidos por la presencia de esta alga. Esta acción sobre la vegetación estaría asociada a cambios en las propiedades físico-químicas de los suelos colonizados por el alga. La mayor disponibilidad de nitrógeno que se halló en estos estudios en las áreas colonizadas por el alga concuerda con lo hallado por Halperin et al. (1976) y Loftis & Kurts (1980).
Desde el punto de vista de la productividad de los suelos alcalinos de la Pampa Deprimida, se estima que la presencia de biodermas algales favorece la sustentabilidad pastoril de los mismos, en función de una mayor cobertura vegetal y diversidad específica. La importancia que tiene la biodiversidad sobre la estabilidad y el funcionamiento de los ecosistemas naturales sometidos a pastoreo ha sido destacada por varios autores (Archer & Smeins 1991; Tilman & Downing 1994).
Los resultados de este trabajo muestran que el pisoteo de los vacunos disminuye la presencia del alga. De allí que quienes manejan el pastoreo con vacunos en la Pampa Deprimida y utilizan la vegetación natural casi comoúnico recurso forrajero deben tomar conciencia de las ventajas que tiene asegurar la presencia del alga como un integrante más de la flora regional. Ello requiere un manejo controlado del pastoreo, evitando el mismo cuando el suelo se halla muy húmedo o encharcado y realizándolo cuando está preponderantemente seco.
AGRADECIMIENTOS
Agradecemos al Ing. Agr. E. A. Hoffmann por su valioso apoyo brindado durante los trabajos desarrollados en el invernáculo.
BIBLIOGRAFÍA
1. ALCONADA, M; OE ANSÍN; RS LAVADO; VA DEREGIBUS; G RUBIO & FH GUTIÉRREZ BOEM. 1993. Effect of retention of run-off water and grazing on soil and on vegetation of a temperate humid grassland. Agr. Water Manage. 23:233–246. [ Links ]
2. ANSÍN, OE. 1995. Pastoreo de comunidades halomórficas de la Pampa Deprimida. Tesis de Magister, Facultad de Agronomía, Universidad de Buenos Aires. 141 pp. [ Links ]
3. ARCHER, S & FE SMEINS. 1991. Ecosystems-level processes. Pp. 109–140 en: RK Heitschmidt & JW Stuth (eds). Grazing management: an ecological perspective. Timber Press. Portland, EEUU. [ Links ]
4. BAILEY, DA; AP MAZURAK & JR ROSOWKSI. 1973. Aggregation of soil particles by algae. J. Phycol. 9:99–101. [ Links ]
5. BERENDSE, F. 1985. The effect of grazing on the outcome of competition between plant species with different nutrient requirements. Oikos 44:35–39. [ Links ]
6. BIGHAMS, JM & JM BARTELS. 1996. Methods of soil analysis. Soil Science Society of America. Wisconsin, EEUU. 1385 pp. [ Links ]
7. BROCK, TD. 1973. Primary colonization of Surtsey with special reference to the blue-green algae. Oikos 24:239–243. [ Links ]
8. BROWN, D. 1954. Methods of surveying and measuring vegetation. Commonwealth Bureau of Pastures and Field Crops. Hurley, Reino Unido. [ Links ]
9. CONNELL, JH & RO SLATYER. 1977. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 111:1119–1144. [ Links ]
10. DAVIDOVICH BOBERI, A. 1980. Técnicas de laboratorio utilizadas en los análisis de semillas. Boletín de divulgación técnica N° 48, INTA EERA Pergamino. Pergamino, Argentina. [ Links ]
11. DODDS, WK; DA GUDDNER & D MOLLENHAUER. 1995. The ecology of Nostoc. J. Phycol. 31:2–18. [ Links ]
12. DEREGIBUS, VA; E JACOBO & OE ANSÍN. 2001. Grassland use and plant diversity in grazed ecosystems. Pp. 879–882 en: JA Gomide, WR Soares Mattos & SC da Silva (eds). Proceedings of the XIX International Grassland Congress. Brazilian Society of Animal Husbandry. Piracicaba, Brasil. [ Links ]
13. FACELLI, JM; RJC LEÓN & VA DEREGIBUS. 1988. Community structure in grazed and ungrazed grasslands sites in the Flooding Pampa Argentina. Am. Midl. Nat. 121:125–133. [ Links ]
14. FULLER, WH; RE CAMERON & E RAICA, JR. 1960. Fixation of nitrogen in desert soils by algae. Pp. 617–624 en: CA Black & NG Carr (eds). Transactions of the 7th International Congress of Soil Science. American Soil Science Society. Berkeley, EEUU. [ Links ]
15. HALPERÍN, DR; MCZ MULE & GZ CAIRE. 1976. Biodermas algales como fuente de nitrógeno en suelos sub-húmedos y semi-áridos (Provincia de Chaco y Formosa, Argentina). Darwiniana 20:341–370. [ Links ]
16. HORI, K; T UENO-MOHRI; T OKITA & G ISHIBASHI. 1990. Chemical composition, in vitro protein digestibility and in vitro available iron of blue green alga, Nostoc commune. Plant Food. Hum. Nutr. 40:223–229. [ Links ]
17. INSAUSTI, P; EJ CHANETON & A SORIANO. 1999. Flooding reverted grazing effects on plant community structure in mesocosms of lowland grassland. Oikos 84:266–276. [ Links ]
18. LEÓN, RJC; SE BURKART & CP MOVIA. 1979. Relevamiento fitosociológico del pastizal del norte de la Depresión del Salado. INTA, Buenos Aires. 90 pp. [ Links ]
19. LOFTIS, SG & EB KURTS. 1980. Field studies of inorganic nitrogen added to semiarid soils by rainfall and blue-green algae. Soil Sci. 129:150–155. [ Links ]
20. MALAM ISSA, O; Y LE BISSONNAIS; C DÉFARGE & J TRICHENT. 2001. Role of a cyanobacterial cover on structural stability of sandy soils in the Sahelian part of western Niger. Geoderma 101:15–30. [ Links ]
21. MATTEUCCI, SD & A COLMA. 1982. Metodología para el estudio de la vegetación. OEA, Washington DC. 173 pp. [ Links ]
22. MAYLAD, HF & TH MCINTOSH. 1966. Distribution of nitrogen fixed in desert algal-crust. Proc. Am. Soil Sci. Soc. 30:606–609. [ Links ]
23. MCNAUGHTON, SJ; FF BANYKWA & MM MCNAUGHTON. 1997. Promotion of the cycling of diet-enhancing nutrients by African grazers. Science 278:1798–1800. [ Links ]
24. MEGHARAHJ, M; AS RAO & K VENKATESWARLU. 1993. Influence of rice straw amendment on persistence and algal toxicity of p-nitrophenol in soil. Soil Biol. Biochem. 25:1185–1188. [ Links ]
25. MOLLENHAUER, D. 1986. Blaualgen der Gattung Nostoc. Ihere Rolle in Forschung und Wissenschaftsgeschichte III. Nat. Mus. 116:43–59. [ Links ]
26. MOLLENHAUER, D; B BUDEL & B MOLLENHAUER. 1994. Algological studies. Arch. Hydrobiol. 105:189–209. [ Links ]
27. RODGERS, GA; B BERGMAN; E HENRIKSSON & M UDRIS. 1979. Utilization of blue-green algae as biofertilizers. Plant Soil 52:99–107. [ Links ]
28. ROGER, PA; WJ ZIMMERMAN & TA LUMPKIN. 1993. Microbiological management of wetland rice fields. Pp. 417–455 en: FBL Metting (ed). Soil microbial ecology: applications in agricultural and environmental management. Dekker, New York, EEUU. [ Links ]
29. ROGERS, SL & RG BURNS. 1994. Changes in aggregate stability, nutrient status, indigenous microbial populations, and seedling emergence, following inoculation of soil with Nostoc muscorum. Biol. Fert. Soils 18:209–215. [ Links ]
30. ROTHER, JA & BA WHITTON. 1989. Nitrogenase activity of blue-green algae on seasonally flooded soils in Bangladesh. Plant Soil 113:47–52. [ Links ]
31. SEMMARTIN, M & M OESTERHELD. 2001. Effects of grazing pattern and nitrogen availability on primary productivity. Oecologia 126:225–230. [ Links ]
32. STEEL, RGD & JM TORRIE. 1990. Bioestadística: principios y procedimientos. 1ra edn. McGraw Hill Latinoamericana SA. México. 622 pp. [ Links ]
33. SVIRCEV, Z; I TAMAS; P NENIN & A DROBAC. 1997. Cocultivation of N2-fixing cyanobacteria and some agriculturally important plants in liquid and sand cultures. Appl. Soil Ecol. 6:301–308. [ Links ]
34. TILMAN, D. 1993. Community diversity and succession: the roles of competition, dispersal, and habitat modification. Pp. 327–344 en: ED Schulze & HA Mooney (eds). Biodiversity and ecosystem function. Springer-Verlag. Berlin, Alemania. [ Links ]
35. TILMAN, D & JA DOWNING. 1994. Biodiversity and stability in grasslands. Nature 367:363–365. [ Links ]
36. UNITED STATES DEPARTMENT OF AGRICULTURE. 1965. Manual for testing agricultural and vegetable seeds. Ed. Herrero. México. 525 pp. [ Links ]
37. VERVOORST, F. 1967. Las comunidades vegetales de la Depresión del Salado (Prov. de Bs. As.). INTA, Buenos Aires. 259 pp. [ Links ]
38. WEIHER, E. & PA KEDDY. 1995. The assembly of experimental wetland plant communities. Oikos 73:323–335. [ Links ]
39. WITTY, JF; PJ KEAYM; PJ FROGATT & PJ DART. 1979. Algal nitrogen fixation on temperate arable fields. Plant Soil 52:151–164. [ Links ]
40. ZIMMERMAN, W; B METTING & W RAYBURN. 1980. The occurrence of blue-green algae in silt loams of Whitman County, Washington. Soil Sci. 130:11–18. [ Links ]

