










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.13 no.2 Córdoba July/Dec. 2003
TRABAJOS ORIGINALES
The spawning migration of anadromous rainbow trout in the Santa Cruz River, Patagonia (Argentina) through radio-tracking
Carla Riva Rossi*, Milagros Arguimbau & Miguel Pascual
Centro Nacional Patagónico, Puerto Madryn, Chubut, Argentina
*Centro Nacional Patagónico, Blvd. Brown 3500, Puerto Madryn, 9120, Chubut, Argentina. Fax: 02965 - 451543, phone 02965 - 451024. Email: rivarossi@cenpat.edu.ar
Recibido: 29 de noviembre de 2002;
Fin de arbitraje: 1 de abril de 2003;
Revisión recibida: 5 de junio de 2003;
Aceptado: 9 de junio de 2003
ABSTRACT. Adult anadromous rainbow trout (Oncorhynchus mykiss) from the Santa Cruz River were implanted with radio-transmitters in May 2000 as they ascended the river from the ocean to locate spawning areas, assess the range and timing of the spawning migration, and determine the composition of the spawning populations. Radio-tagged fish revealed geographic and temporal migration patterns and previously unidentified spawning areas. Telemetry data indicated that anadromous fish dwell and spawn in the main stem river. Range of the total detected upstream migration varied between 2-280 km. Spawning begins by mid winter and peaks in the spring, matching the raise in water temperature, which ranged from 4ºC at the beginning of the spawning season to 10ºC at the end of the season in December. This pattern is similar to that of trout indigenous to the Pacific Northwest, USA, in which a raise in the water temperatures stimulates the onset of spawning. The arrangement of spatial locations of anadromous fish also indicated that spawning occurs in different sections along the main stem, giving some credence to the idea that subdivision at the intra-population level may exist. These results provide strong support to the idea that anadromous fish are at present more closely related to resident fish occurring in the upper Santa Cruz River, while resident fish from the lower reaches of the river may well constitute a separate spawning aggregation.
Keywords: Oncorhynchus mykiss; Steelhead; Anadromy; Residency; Radio-telemetry; Surgical implantation.
RESUMEN. La migración reproductiva de la trucha arco iris anádroma del río Santa Cruz mediante radio-marcado: El río Santa Cruz, en Patagonia, es el único ambiente en el mundo donde la trucha arco iris introducida desarrolló poblaciones migratorias facultativas, donde una fracción de los individuos completa su ciclo de vida enteramente en agua dulce, mientras que otros realizan migraciones periódicas al mar con fines tróficos. En otoño de 2000 se implantaron radiotransmisores en siete individuos anádromos durante su ascenso al río desde el océano al inicio de la migración reproductiva, a fin de localizar las áreas de desove dentro de la cuenca, determinar la duración de la temporada reproductiva y evaluar la composición de la población reproductiva. La re-localización de los peces radio-marcados reveló patrones generales de la migración reproductiva de estos peces y permitió identificar sus áreas de desove. Desde julio a diciembre de 2000, todos los peces radio-marcados fueron localizados en distintos sectores del cauce principal del río Santa Cruz. La distancia máxima recorrida por los individuos radiomarcados durante la migración reproductiva varió entre 2-280 km. La temporada de desove comenzó a mediados de invierno, alcanzando el nivel máximo hacia primavera. Hacia diciembre ninguno de los peces radio-marcados se encontraban en el río. El pico de desove coincidió con un ascenso notable de la temperatura promedio del agua, la cual se incrementó desde 4ºC a principios de la temporada hasta 10ºC hacia fines de la temporada reproductiva. Los datos de telemetría revelaron que las truchas anádromas utilizan distintas áreas de desove dentro del cauce principal, lo cual sugiere un cierto grado de segregación a nivel intra-poblacional. Estos resultados indicarían que la forma anádroma se encuentra más estrechamente relacionada con las truchas arco iris residentes del curso medio y superior que con las truchas residentes que habitan en la desembocadura del río, las cuales parecen constituir una unidad poblacional separada.
Palabras clave: Oncorhynchus mykiss; Steelhead; Anadromía; Residencia; Radio-telemetría; Implante quirúrgico.
INTRODUCTION
The introduction of salmonids in lakes and rivers of Patagonia, the southernmost region of South America, started early in the 20th century with the primary goal of establishing wild populations (Tulian 1908). In Argentinean Patagonia, rainbow trout (Oncorhynchus mykiss) is the most widely distributed salmonid species, with feral populations inhabiting most river basins in the region (Pascual et al. 2001). Like all other known introduced rainbow trout around the world, typical Patagonian fish remain in freshwater throughout their entire life cycle, with a life history similar to that of resident populations in rivers and headwater lakes in western North America (Wydoski & Whitney 1979). Rainbow trout in the Santa Cruz River (Figure 1) display this "resident" behaviour, but there is an alternative life-history type: anadromous fish (steelhead) that migrate to the ocean both prior to maturation and between subsequent spawning events (Pascual et al. 2001). Whereas the coexistence of anadromous and resident forms is common in rivers throughout the native range of rainbow trout (Zimmerman & Reeves 2000), the Santa Cruz River is the only location in the world where introduced rainbow trout are known to have developed an anadromous life history.

Figure 1: Location of the Santa Cruz River and tributaries in southern Patagonia, Argentina, and sampling localities mentioned in the text.
Figura 1. Ubicación del Río Santa Cruz y de sus principales tributarios en el sur de Patagonia, Argentina, y de las estaciones de muestreo mencionadas en el texto.
As in many other salmonid populations with this dual life-history behaviour, the way and extent to which the two ecotypes intermingle in Santa Cruz River is uncertain. A previous genetic analysis based on microsatellite loci revealed no differentiation between anadromous and resident trout collected from a single location on the upper main stem (Pascual et al. 2001). Furthermore, several microsatellite loci showed a significant heterozygous deficiency for anadromous rainbow trout, a pattern consistent with a scenario where mating is spatially or temporally segregated within the migratory form. Altogether, these data hint at the existence of subdivision at the intra-population level (Bernatchez et al. 1998).
It is becoming apparent that the relationship between resident and migratory forms and the patterns of population structure in the basin cannot be assessed by using genetic data alone. This requires an assessment of the spawning migration that includes identification of spawning locations and timing of spawning events, identification of reproductive units in the basin, and a determination of the incidence of anadromy among different units. Despite extensive surveys throughout the basin since 1997 and numerous interviews with fishermen and residents of the upper basin, the progress of the upriver migration following river entrance and the exact location of the spawning areas remain unknown. The chalky water of this large, glacial- fed river precludes visual detection of spawners and spawning beds.
Radio-tracking experiments are increasingly used for locating and characterizing spawning runs of migratory salmonids in many large rivers of North America (Burger et al. 1985; Eiler 1990; Parkinson et al. 1999), and it appears to be an effective method for locating fish in remote or inaccessible areas. The purpose of this study was to use radio-telemetry to track adult anadromous rainbow trout as they ascend the Santa Cruz River from the ocean and thereby provide the first description of the timing and geographic extent of this anadromous spawning migration.
METHODS
Study area
The upper Santa Cruz basin is dominated by two large glacial-fed lakes, Viedma and Argentino, interconnected by La Leona River, the second most important river of the drainage after the Santa Cruz main stem (Figure 1). Numerous second to third order tributaries feed these lakes; some of them have remained largely isolated due to glacier closure. Landlocked populations of rainbow trout inhabit most of these streams. The main stem Santa Cruz River (50ºS) originates in Argentino Lake and flows for 382 km along the Patagonian plateau to drain into the Atlantic Ocean through a 25 km-long estuary (Figure 1). The river has unstable, braided, gravelrich beds forming valleys often more than 1 km wide. Few springs and small tributaries enter the main stem river, none of them significant from the point of view of their trout populations. The Santa Cruz River supports an important steelhead fishery from February through May. The relative inaccessibility of the upper reaches limits most fishing to the lowest 50 km. Most fishing occurs during migration; however, in recent years, as fishermen have ventured further from traditional fishing localities on the lower section, fishing also occurs in remote localities during the holding period before spawning.
Fish collection and handling procedures
Anadromous rainbow trout were caught with hook and line in March and April 2000 on a 20-km section of the river adjacent to the town of Piedrabuena (Figure 1) as they entered the river from the ocean. Fish were kept in a natural pond belonging to the Piedrabuena municipal hatchery until the implantation of transmitters. On 29 April 2000, five healthy rainbow trout ranging 490-780 mm in fork length (Table 1) were recaptured from the pond by seining and transferred to a hatchery raceway. In preparation for surgical implantation of radio-transmitters, each individual was anaesthetized with 85 mg/L solution of MS-222 (tricane methanesulfonate) buffered with sodium bicarbonate until total loss of equilibrium, consciousness, muscular tone and reflexes (approximately 4-5 min). On 7 May, two additional rainbow trout (Table 1) were caught by hook and line at Estancia Rincón Grande (Figure 1) located 150 km upstream from the town of Piedrabuena and immediately anaesthetized for on-site implantation of radio transmitters.
Table 1. Fork length (mm) of the anadromous rainbow trout radio-tagged in the Santa Cruz River (Patagonia), dates and positions (in rkm) of tagging and release sites (7 May), and of relocated fish (August-December), and maximum distance travelled (km) during upstream migration.
Tabla 1. Longitud (mm) de las truchas arco iris anádromas radio-marcadas en el río Santa Cruz (Patagonia), fechas y posiciones (en kilómetros de río) de los sitios de marcado (7 de mayo) y de los peces relocalizados (agosto-diciembre), y la distancia máxima (km) recorrida durante la migración reproductiva.

Surgical tag implantation technique
Surgical procedures followed Adams et al. (1998), but several steps were modified for its application on larger fish. Trout were positioned ventral side upward in a wooden V-shaped surgical trough. A soaked soft towel was used to cushion the surface and to stabilize the fish, holding it in the trough with little tissue damage (Summerfelt & Smith 1990).
Prior to surgery the equipment was sterilized and the incision area disinfected with tincture of iodine applied with a soft brush. To implant the radio transmitter (155 MHz; 20g in air, 46 x 17 mm in diameter; 26-cm flexible external antenna; 40 pulses/min; Model 5955, ATS Inc., Isanti, MN, USA) a 60-mm long incision was made parallel to the midventral line, starting 100 -150 mm in front of the pelvic girdle. The incision was deep enough to penetrate the peritoneum. Antibacterial Terra-Cortril™ (oxytetracycline) was sprayed into the opening to prevent infections. The outlet for the antenna was made using an intravenous catheter and needle (Abbocath™-T Nº14). The skin was pierced from the outside with an Abbocath™, 50mm in a lateral and slightly caudal position relative to the main incision. The Abbocath™ was then pushed into the opening, inserted inwards through the body wall, and the needle withdrawn from the incision leaving the catheter in order to guide the antenna outwards.
Once the transmitter had been positioned, the antenna was threaded through the catheter, and both were pulled out while positioning the transmitter in the body cavity. Prior to closure the incision was again sprayed with antibacterial solution. The peritoneum and the skin were sutured separately with a continuous suture; the first, with sterile absorbable suture on 25-mm non-cutting curved 0.5-circle needle (Monocryl 2-0, Ethicon™), and the latter with sterile non-absorbable suture on 26 mm triangular cutting needle (Mononylon 2-0, Ethiconô™). Suturing was reinforced with stainless steel staples. The antenna was affixed to the side of the fish with a single stitch. The whole area was disinfected with antibacterial solution and painted with tincture of iodine. Finally, the sutured incision was smeared with petroleum jelly for temporary waterproofing (Summerfelt & Smith 1990).
Throughout the whole surgical procedure, gills were alternatively flushed with a solution of diluted anaesthetic (MS 222 10 mg/L) and oxygenated water using a 60 ml syringe. The flow rate of anaesthetic was approximately 120 ml/min. During the last minute of surgery, only freshwater was provided in order to start the recovery process. Each surgery averaged 10-15 min (digital photos of the surgical procedure are available from the authors).
Following surgery, fish implanted at the hatchery were returned to a raceway with fresh running water until they swam upright. They were then placed in a holding tank in the hatchery and kept for three days to verify the healing process and ensure full recovery before release. Tagged fish were transported to the release sites in 1 x 0.65 x 0.65 m chests filled with river water. Before release, the condition of all fish was checked.
The surgical procedure for fish operated in the field was the same as that applied in the hatchery. Fish were held for 20 min in an individual net pen anchored to the shoreline and were released as soon as they regained consciousness and equilibrium. The entire process from capture of fish to final release lasted approximately 35 min.
Surveillance equipment and radio-tracking techniques
Each transmitter operated on a unique frequency, within the range 155000-155100 MHz. Individual fish were relocated with a manual Fieldmaster Model R161N receiver and a scanning Challenger Model R2100 receiver, and a High Frequency 4-Element Yagi Antenna (ATS Inc., Isanti, MN, USA). These units were used in vehicles, boats, and aeroplanes. From August to December 2000, five telemetry surveys were conducted upriver from Puerto Santa Cruz to Argentino Lake (Figure 1) with a low-winged Piper Archer aircraft. During aerial tracking, the antenna was fixed to a custom-made V shaped aluminium holder attached to the lower face of the aircraft right wing, with elements in the horizontal position pointing frontward and backwards. In this way, the most sensitive planes of the antenna pointed towards the river, perpendicularly to the direction of travel. Average flight altitude was 300 m above ground, an altitude at which transmitter signal range did not exceed 500 m.
The location of tagged fish was determined when the observer recorded the peak radio signal and the geographic coordinates from a portable GPS unit. River sites where relocation occurred more than once after upstream movement ceased were noted as potential spawning areas (Burger et al. 1985). When possible, aerial surveillance was corroborated with radio tracking by boat or from shore. A map of the Santa Cruz River main stem was digitized and referenced with geographic coordinates. This information was then used to plot the locations and movements of individual fish, computing distances in river-kilometres (rkm; i.e., distance in kilometres from the mouth of the river).
Determination of spawning areas and times
Determination of spawning areas and times followed the criteria established by Burger et al. (1985): subsequent relocations of one fish without upstream movements connoted a spawning area; spawning times were periods when a tagged fish was relocated within the same river reach on two or more consecutive flights. Onset of spawning was determined as the period when tagged fish ceased to move upstream and remained stationary in the same stretch of river.
Field census
We attempted to verify spawning at particular locations where fish lingered during consecutive telemetry surveys (Estancia Nueva Lubeck and Estancia El Rincón, Figure 1). We visited the area three times: 6-10 September, 26-29 September, and 24-27 October. During those surveys, gill nets were set overnight (monofilament, of mesh size 120 and 180 mm) at two river sections known as Primer Laberinto (rkm 320) and Segundo Laberinto (rkm 350) (Figures 1) for a total of 15 nights. Scale pattern analysis was implemented for distinguishing anadromous fish from resident fish and for determining the frequency of repeat spawning (Lamond 1916; Nall 1930; Shapovalov & Taft 1954; Ducharme 1969; Pascual et al. 2001). We also conducted a thorough recognizance of a section of approximately 30 km of river from a boat and from shore in search of spawning fish, redds or any other signs of spawning activity. The peak of the spawning season was determined as the time interval when the frequency of mature fish caught in gillnets was highest.
RESULTS
Of the seven fish originally radio-tagged, Male 1 and Male 2 were relocated in the same exact position throughout all the flights, as well as from the coast, indicating that the fish had either died or shed the tag. The remaining five fish moved upstream after tagging, all of them staying in the main stem Santa Cruz throughout the study (Table 1 and Figure 2). Males 1-4 and Female 1 implanted at the hatchery remained stationary at the site of release for at least two days. Male 5 and Female 2, tagged at the field and released immediately after tag implantation, resumed upriver movements immediately. Male 3 and Female 1 moved 25 and 16.8 km upstream, respectively, from the site of release, remained stationary at that river reach until 21 October, when Male 3 was no longer detected and Female 1 resumed migration and moved 32.6 km upstream. Male 5 first moved 64 km upstream from the site of release and then migrated downstream. Male 4 and Female 2 migrated upstream to a river section previously considered suitable for spawning and remainedr at that reach, moving within a stretch of 2 km until 21 October when Female 2 migrated to the ocean. Relocations of Male 3 and Male 4 were accomplished from the coast on 4 September and 21 October, respectively (Figure 2). By 3 September, all fish remained in the river, by 21 October four of them, and by the time of the last flight on 17 December only Male 5 remained in the river (Figure 2).

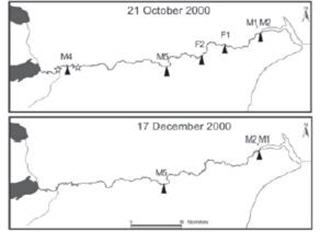
Figure 2. Tagging site and movements of radio-tagged anadromous rainbow trout in the Santa Cruz River (Patagonia) during spawning migration. Triangles indicate fish positions and stars indicate gill net sites. Fish codes are the same as in Table 1.
Figura 2. Sitio de marcado y desplazamientos de individuos anádromos radio-marcados en el Río Santa Cruz (Patagonia) durante la temporada de migración reproductiva. Referencias: los triángulos indican la posición de los peces y las estrellas los sitios de captura. Los códigos de los peces son los mismos que en la Tabla 1.
Tracking results suggest that the sections of the Santa Cruz River visited by Male 3 and Female 1 (rkm 55-80), and Male 4 and Female 2 (rkm 320-350) are potential spawning beds (Figure 2). In order to verify spawning in those particular locations, from August to October the area was explored and gill nets were set overnight. During these surveys, only one anadromous mature female was caught in the area located between rkm 55-80. However, several anadromous, river resident and lake-dwelling fish were caught in river reach located between rkm 320 - 350 (Table 2). Although all anadromous fish were at an advanced stage of maturation, most nonmigratory individuals appeared to be immature fish, because they were feeding and no secondary sexual characters were evident. The proportion of mature anadromous fish increased significantly during mid to late September, suggesting that the peak of the spawning season occurred around that time. The section of the river visited by Male 5 (rkm 202) could not verified as a spawning site, because the area was not surveyed.
Table 2. Characteristics of fish caught at sites where radio-tagged fish were relocated in the Santa Cruz River (Patagonia), during September and October 2000.
Tabla 2. Características de los peces capturados en los sitios donde los peces radio-marcados fueron relocalizados en el río Santa Cruz Patagonia), durante septiembre y octubre de 2000.

DISCUSSION
Radio tracking is quickly becoming a popular research tool in fish ecology research. However, relatively little work has been conducted in Argentina using this technique. The primary reasons for the scarcity of telemetry studies, especially in aquatic habitats, have been the cost and the practical difficulties related to tagging a representative sample of the stock. Another obvious concern when conducting behavioural studies is that tagging does not alter fish behaviour, or that the behavioural effects are at least minimal and known to the researchers (Demko et al. 1998). Many recent studies have indicated that fish behaviour remained unchanged after tag implantation (Adams et al. 1998; Demko et al. 1998; Parkinson et al. 1999). Adams et al. (1998) indicated that the relation of transmitter size to fish size played an important role in determining how well the tagged fish performs after being fitted with either surgical or gastric transmitters. Holding time prior to transmitter implantation and final release is another putative factor affecting fish behaviour. For instance, holding time may have influenced the migratory performance of Male 3 that showed a relatively shorter upstream movement as compared to other implanted fish (Table 2). Furthermore, surgical procedures could have produced detrimental effects on the two fish that did not resume upstream migration after tagging, which could either died or shed the tag after release. Other than this, we did not observe any considerable impact on fish behaviour that invalidates our results.
Radio tracking was successfully applied to Santa Cruz River migratory trout, gathering some basic information regarding their spawning behaviour. Although we tracked a small number of fish, the consistency of some results allows us to draw extremely helpful conclusions to direct future research. Given the physical characteristics of the Santa Cruz River, the remoteness of the spawning areas, and the lack of effective mass-capture techniques, conventional mark-recapture techniques were either not feasible or would provide estimates that may be imprecise. Radio telemetry provided us with a means of monitoring fish movements and locating spawning areas that were not otherwise readily visible.
The main finding of this study is that anadromous fish dwell and spawn in the upper main stem river. This observation is consistent with the pattern of population structure depicted by previous genetic data (Pascual et al. 2001), providing strong support to the idea that the anadromous form is at present more closely related to resident fish occurring in the upper Santa Cruz River, while the resident fish from the lower reaches of the river may well constitute a separate spawning population. Spawning interactions may occur in sympatric populations with overlapping spawning distributions and timing. In the upper Yakima River where the two forms occur sympatrically, Pearsons et al. (1993) speculated that the small population of steelhead, combined with the large population of resident rainbow trout increases the probability for spawning interactions. On the contrary, sympatric populations of anadromous steelhead and resident rainbow trout in the Deschutes River (Oregon, USA) occur with little or no spawning interactions because of spatial or temporal spawning separation (Schroeder & Smith 1989; Zimmerman & Reeves 2000).
Telemetry data indicated that upstream movement of Santa Cruz River anadromous rainbow trout proceeds rapidly. Once the marked fish were released after tag implantation, they moved directly to the spawning locations, holding position or "wandering" within the same river reach until the time of spawning, at least four months later. The tracking experiment also revealed the general timing of the spawning migration. Spawning does not begin until mid winter (August) and appears to peak in the spring (September-October). Given that the peak of river entrance occurs in early April and that most fish appear to stay in the river until early/mid November, we can estimate river dwelling of repeat spawners to last for approximately seven months and ocean migration for five months. The peak of spawning events appears to coincide with the raise in water temperature, temperature, which went from 4ºC at the onset of the spawning season to 10ºC at the end of the season. This pattern is similar to that of trout indigenous to the Pacific Northwest, USA, in which a raise in the water temperatures stimulates the onset of spawning (Hicks 1999).
Although these results do not provide information about the degree of relatedness between ecotypes or about the mechanisms underlying their introgression, they allow us to sketch a preliminary map of likely subpopulations, setting the stage for future research. We were able to verify the occurrence of spawning in the upper section by collecting anadromous spawners in the field, together with their likely resident kin: river and lake resident fish that spawn in the upper section of the Santa Cruz. The utilization of lower sections of the river by spawning anadromous and resident fish remains unexplored. Samples from those spawning aggregations, together with those already collected from the upper section, will allow us to perform a finescale genetic characterization of anadromous and resident fish and infer patterns of microgeographic genetic structure at the intrapopulation level (Foote et al. 1989).
ACKNOWLEDGEMENTS
The Subsecretaría de Pesca y Actividades Portuarias, Santa Cruz Province, and the Agencia Nacional de Promoción Científica y Tecnológica (PICT-01- 04582 to MP) supported this work. We thank three anonymous reviewers for their comments on the manuscript. We also would like to thank N. Adams for his valuable assistance on tag implant techniques. F. Quintana for sharing with us his knowledge about radio-telemetry techniques. L. Pellanda and P. Fernández for assistance in the field and during telemetry surveys. Dr. M. Triputti and I. Vidal provided us instruction on surgery techniques. Municipalidad of Piedra Buena for providing us with field transportation. R. Hudson and personnel from Estación Municipal de PisciculturaìIsla Pavónî for their support and assistance. J. Uhrig, R. Moreno, H. Maldonado, P. Chaparro, C. and F. Calisto, and M. González, fishermen of Piedra Buena and El Calafate, for helping us with fish collection. We thank O. Olais for his technical advice on aerial radio tracking techniques. Pilot M. Calvo and the board of Aeroclub Puerto Santa Cruz for their support, air transportation and assistance on telemetry surveys, M. Novara for assistance on GIS data analysis. J. Ciancio for field support, assistance on data collection and surgery procedures. D. Derejibus and V. Cherbukov provided us with access to Estancia Nueva Lubeck and Estancia El Rincón, respectively, as well as their hospitality.
REFERENCES
ADAMS, NS; DW RONDORF; SD EVANS; JE KELLY & RW PERRY. 1998. Effects of surgically and gastrically implanted radio transmitters on swimming performance and predator avoidance of juvenile Chinook salmon (Oncorhynchus tshawytscha) Can. J. Fish. Aquat. Sci. 55:781-787. [ Links ]
BERNATCHEZ, L; JB DEMPSON & S MARTIN. 1998. Microsatellite gene diversity analysis in anadromous artic char, Salvelinus alpinus, from Labrador, Canada. Can. J. Fish. Aquat. Sci. 55:1264-1272. [ Links ]
BURGER, CV; RL WILMOT & DV WANGAARD. 1985. Comparison of spawning areas and time for two runs of Chinook salmon (Oncorhynchus tshawytscha) in the Kenai river, Alaska. Can. J. Fish. Aquat. Sci. 42:693-700. [ Links ]
DEMKO, DB; C GEMPERLE; SP CRAMER & A PHILLIPS. 1998. Evaluation of juvenile chinook behavior, migration rate and location of mortality in the Stanislaus River through the use of radio tracking. SP Cramer & Associates. Gresham. [ Links ]
DUCHARME, LJ. 1969. Atlantic salmon returning for their fifth and sixth consecutive spawning trips. J. Fish. Res. Board Can. 26:1661-1664. [ Links ]
EILER, JH. 1990. Radio transmitters used to study salmon in glacial rivers. American Fisheries Society Symposium 7:364-369. [ Links ]
FOOTE, CJ; CC WOOD & RE WITHLER. 1989. Biochemical genetic comparison of sockeye salmon and kokanee, the anadromous and nonanadromous forms of Oncorhynchus nerka. Can. J. Fish. Aquat. Sci. 46:149-158. [ Links ]
HICKS, M. 1999. Evaluating standards for protecting aquatic life in Washington's surface water quality standards. Preliminary draft of draft discussion paper (vol. 1) and draft supplementary appendix (vol. 2). WA Dept. of Ecology, Water Quality Program. Olympia. [ Links ]
LAMOND, H. 1916. The sea-trout. A study in natural history. Sherratt and Hughes. Londres. [ Links ]
NALL, GH. 1930. The life of sea trout. Seeley. Londres. [ Links ]
PARKINSON, D; JC PHILIPPART & E BARAS. 1999. A preliminary investigation of spawning migrations of grayling in a small stream as determined by radio-tracking. J. Fish Biol. 55:172-182. [ Links ]
PASCUAL, MA; P BENTZEN; C RIVA ROSSI; G MACKEY; M KINNISON & R WALKER. 2001. First documented case of anadromy in a population of introduced rainbow trout in Patagonia, Argentina. T. Am. Fish. Soc. 130:53-67. [ Links ]
PEARSONS, T; G MCMICHAEL; E BARTRAND; M FISCHER; J MONAHAN & S LEIDER. 1993. Yakima River species interactions studies. Contract DE-BI79-89BP01483, Bonneville Power Administration. Portland. [ Links ]
SCHROEDER, RK & LH SMITH. 1989. Life history of rainbow trout and effects of angling regulations, Deschutes River, Oregon. Information Report N° 89-6, Oregon Department of Fish and Wildlife. Portland. [ Links ]
SHAPOVALOV, L & AC TAFT. 1954. The life histories of the steelhead rainbow trout (Salmo gairdneri) and silver salmon (Oncorhynchus kisutch) with special reference to Waddell Creek, California, and recommendations regarding their management. California Department of Fish and Game Fish Bulletin 98:1-373. [ Links ]
SUMMERFELT, L & LS SMITH. 1990. Anesthesia, surgery, and related techniques. Pp. 213-263 in: CB Schreck & PB Moile (eds). Methods for fish biology. American Fisheries Society. Bethesda. [ Links ]
TULIAN, EA. 1908. Acclimatization of American fishes in Argentina. Bulletin of the Bureau of Fisheries 18:957-965. [ Links ]
WYDOSKI, RS & RR WHITNEY. 1979. Inland fishes of Washington. University of Washington Press. Seattle. [ Links ]
ZIMMERMAN, CE & GH REEVES. 2000. Population structure of sympatric anadromous and nonanadromous Oncorhynchus mykiss: evidence from spawning surveys and otolith microchemistry. Can. J. Fish. Aquat. Sci. 57:2152-2162. [ Links ]

