










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.13 n.2 Córdoba jul./dic. 2003
TRABAJOS ORIGINALES
Incidencia de la estructura del paisaje sobre la composición de especies de aves de los talares del noreste de la provincia de Buenos Aires
Nathalie Horlent1,* , Marcos C Juárez1 & Marcelo Arturi2
1. Facultad de Ciencias Agrarias y Forestales, Facultad de Cs. Naturales y Museo, UNLP, La Plata, Argentina
2. Facultad de Ciencias Agrarias y Forestales, UNLP, La Plata, Argentina
*Laboratorio de Investigación de Sistemas Ecológicos y Ambientales, Fac. de Cs. Agrarias y Forestales-Fac. de Cs. Naturales y Museo UNLP. Email: talares@ceres.agro.unlp.edu.ar
Recibido: 4 de noviembre de 2001;
Fin de arbitraje: 30 de julio de 2002;
Revisión recibida: 25 de noviembre de 2002;
Versión final recibida: 14 de abril de 2003;
Aceptado: 22 de abril de 2003
RESUMEN. Se analizó la incidencia de la estructura del paisaje sobre la composición de la comunidad de aves de los talares de Magdalena, provincia de Buenos Aires. Se estimó la abundancia relativa por especie durante noviembre y diciembre de 2000. Se midió el área de bosque, de pastizal y la longitud total de borde bosque-pastizal en un área cuadrada de 200 m de lado en torno de cada punto de muestreo. El principal eje de variación de la composición estuvo asociado al área de bosque en torno del punto. Se detectaron 11 y 13 especies cuya abundancia estuvo asociada positiva y negativamente con el área de bosque, respectivamente. La riqueza total no varió con ese eje ni con el área de bosque. La riqueza específica de grupos de especies diferenciados por características tróficas y biogeográficas estuvo asociada al área de bosque. Se hallaron correlaciones positivas para las especies con distribución geográfica restringida a regiones de bosques subtropicales y para las que consumen insectos en el follaje arbóreo y frutos. Se hallaron asociaciones negativas con el área de bosque para las especies con distribución geográfica más amplia, presentes en las provincias Pampeana, del Monte o Patagónica, y para aquellas que consumen insectos en los estratos bajos y el suelo, y granos. La longitud de borde bosque-pastizal estuvo correlacionada con el área, y presentó correlaciones muy similares con la riqueza de los diferentes grupos de especies. El Análisis de Correlación Parcial indicó que la longitud de borde no aporta información diferente de la del área. Las relaciones área-diversidad halladas indican que la conservación de los talares en función de las especies de aves debería contemplar los diferentes requerimientos de los dos grupos encontrados. Las áreas con mayor diversidad corresponden a los sitios con más de 50% de cobertura arbórea y a aquellos con alrededor de 5%, independientemente de la estructura del bosque y de la proporción de borde de los parches. Tales situaciones maximizan la probabilidad de encontrar a las especies asociadas a bosque denso y bosque abierto, respectivamente.
Palabras clave: Fragmentación; Bosques; Celtis tala; Conservación; Análisis multivariado.
ABSTRACT. Incidence of landscape structure on bird species composition of a temperate forest in Buenos Aires, Argentina: The effect of landscape structure on bird species composition was studied in a temperate forest in Buenos Aires, central Argentina. A total of 50 sampling points were located along six transects perpendicular to the coastline. Transects were located about 2000 m apart each one and points inside transects were 200-250 m spaced. Relative abundance per species was determined in each point in November-December 2000. A 25 m fixed radius was used. A square area 200-m in side was used to estimate forest area and forest-grassland edge longitude around each point using aerial photographs scale 1:10000. Points were ordered using a Principal Components Analysis. Differences in trophic features and geographic distribution were analyzed for species showing opposite trends in the ordination diagram. The principal axis describing the compositional variation was associated to forest area surrounding sampled points. Total species richness was associated neither to the compositional axis nor to forest area. However, significant relationships were found between forest area and species richness when species were differentiated by biogeographical and trophic characteristics. A positive relation was found for species geographically restricted to subtropical forest regions as well as for those consuming insects on tree foliage and fruits. Negative relations were found for species with a wider geographical distribution including Pampa, Monte and Patagonia provinces, as well as for those consuming insects on the understory and on the ground, and seeds. Forest-grassland edge longitude was correlated to forest area, and both variables were similarly correlated to species richness of different groups. No effect of edge longitude was detected after removing the area effect. A high diversity can be found in areas with either more than 50% or less than 5% cover of forest. Such conditions maximize the probability of finding species associated to dense and open forest, respectively.
Keywords: Forest; Fragmentation; Celtis tala; Conservation; Multivariate analysis.
INTRODUCCIÓN
La incidencia de la estructura del paisaje sobre la biodiversidad fue estudiada por diferentes autores con relación a las comunidades de aves. La riqueza de especies puede ser influenciada por la superficie de hábitat disponible. El número de especies de aves que aparecen en parches de bosque aislados está correlacionado con el área del parche (Burgess & Sharpe 1981; Williamson 1981; Harris 1984) y se ha demostrado que el número de especies que nidifican en parches aislados se incrementa con el área (Galli et al. 1976; Martin 1981; Whitcomb et al. 1981; Ambuel & Temple 1983; Blake & Karr 1984; Freemak & Merriam 1986; Blake 1987). Los parches de mayor tamaño podrían presentar mayor heterogeneidad y una mayor diversidad en la oferta de recursos, permitiendo la existencia de diferentes grupos tróficos (Galli et al. 1976; Willis 1979; Martin 1980, 1981, 1988; Blake 1983; Díaz et al. 1998). Además, las especies que por el tipo de alimento utilizado requieren grandes áreas de forrajeo no podrían estar presentes en los parches de área reducida (Rudnicky & Hunter 1993). La diversidad de aves puede verse afectada no solo por el tamaño de los parches sino también por la proporción de borde. Diferentes autores estudiaron el efecto de los bordes sobre la diversidad de aves, encontrando relaciones positivas, negativas o ningún efecto (Strelke & Dickson 1980; Hansson 1983; Rudnicky & Hunter 1993; Murcia 1995). Además, la sensibilidad de las especies a los cambios en la estructura del paisaje puede diferir entre grupos de especies definidos geográfica y tróficamente (Holmes et al. 1979; Ambuel & Temple 1983).
Los talares (bosques dominados por Celtis tala) de Magdalena constituyen un paisaje complejo con proporciones variables de bosque y pastizal, así como parches de bosque de distinto tamaño y distribución espacial. Estos bosques podrían considerarse como el límite austral de las formaciones boscosas de origen subtropical que corresponden a la Provincia Fitogeográfica del Espinal. Muchas de las especies de aves registradas en los talares son de origen paranense, mesopotámico o chaqueño, y su presencia en esa área está asociada a la existencia de bosques (Cueto 1996; Cueto & Lopez de Casenave 2000a, 2000b). A su vez, las diferencias estructurales entre bosques con diferentes grados de intervención podrían influir sobre la composición de aves, habiéndose encontrado una mayor proporción de insectívoros en los bosques mejor conservados (Cueto 1996). Los cambios en la proporción de bosque y pastizal podrían determinar variaciones en el número de especies que hacen uso de determinados recursos. En áreas con alta disponibilidad de pastizales podrían ser más frecuentes las especies que utilizan los estratos bajos y el suelo, mientras que las especies asociadas al dosel arbóreo podrían ser más frecuentes en áreas con mayor disponibilidad de bosque. Sobre la base de la información disponible es de esperar un cambio en la composición de especies de aves en relación con las diferentes proporciones de bosque y pastizal, y que las especies con tendencias opuestas difieran en sus características tróficas y biogeográficas. Los sitios con mayor disponibilidad de bosque deberían presentar (1) mayor diversidad, (2) mayor riqueza de especies que se alimentan en el follaje y (3) mayor riqueza de especies biogeográficamente relacionadas con áreas de bosques subtropicales que los sitios con menor disponibilidad de bosque. Además, dada una misma proporción de bosques, (4) las áreas con mayor longitud de borde bosque-pastizal deberían presentar mayor diversidad.
MÉTODOS
El muestreo se llevó a cabo en los bosques de Celtis tala del partido de Magdalena, provincia de Buenos Aires, entre las localidades de Magdalena y Punta Indio. La estructura de estos bosques fue estudiada por Goya et al. (1992). Las especies arbóreas dominantes son Celtis tala y Scutia buxifolia. Los bosques están asociados a depósitos cuaternarios de materiales calcáreos de origen marino. Estos depósitos se disponen paralelos a la costa formando cordones de relieve positivo y tamaño variable que alternan con áreas más bajas ocupadas por pastizales. El tamaño, la forma y la distribución de los parches de bosque están condicionados por la disposición de los cordones calcáreos, así como por la historia de uso. Los parches varían entre 1-30 ha (Goya et al. 1992). El bosque fue afectado por la extracción de materiales calcáreos del subsuelo, el aprovechamiento de los árboles para leña y el desmonte para establecer áreas de pastoreo (Goya et al. 1992). En consecuencia, dentro del área estudiada se encuentran porciones con diferente grado de fragmentación y bosques con diferente desarrollo estructural. Las actividades económicas que provocaron la reducción de superficie de los talares permanecen en la actualidad sin cambios que pudieran adaptarlas a un manejo compatible con la conservación.
El muestreo de aves se realizó entre los meses de noviembre y diciembre de 2000; en estos meses se registra el mayor número de especies en el área (Cueto 1996; Soave et al. 1999). Se utilizó la técnica de conteo por puntos extensivo (Ralph et al. 1996), con un tiempo de observación de 10 min y un radio fijo de 25 m (20 m en tres sitios con baja visibilidad). Se realizaron registros visuales y auditivos desde y hasta cuatro horas después de la salida del sol. Se distribuyeron 50 puntos de muestreo en parches de bosque entre las localidades de Magdalena y Punta Indio. El área circular de muestreo quedó en todos los casos incluida dentro de los parches de bosque muestreados, por lo que los registros de aves corresponden solo a ese ambiente. Los puntos se localizaron a lo largo de seis transectas perpendiculares a la línea de la costa, separadas entre sí por aproximadamente 2000 m. La separación entre los puntos dentro de cada transecta fue de 200-250 m. Cada punto fue visitado dos veces durante el período de muestreo y se promediaron los resultados obtenidos. El distanciamiento entre puntos dentro de una misma transecta es el recomendado para evitar la falta de independencia entre observaciones vecinas (Ralph 1996). Se calculó la abundancia por especie para cada punto como el cociente entre el número de individuos registrados y el área circular correspondiente al radio utilizado. Se definieron áreas cuadradas de 200 m de lado en torno de cada punto, en las que se midió el área de bosque y de pastizal y la longitud total de borde bosque-pastizal. Dichas determinaciones se realizaron utilizando fotos aéreas a escala 1:10000, tomadas en 1991.
Los puntos fueron ordenados en función de la abundancia de las especies de aves mediante un Análisis de Componentes Principales (Digby & Kempton 1987). El análisis se realizó a partir de una matriz de correlación, por lo que los resultados reflejan similitudes en la distribución entre puntos aún cuando las especies presentan valores muy diferentes de abundancia. Debido a que los ejes obtenidos se utilizaron para analizar diferencias entre puntos respecto de la estructura del paisaje y su relación con la composición de especies, se evaluó la medida en que la pertenencia a una misma transecta determinó mayor similitud en la composición en contraposición a puntos de transectas diferentes. Con esa finalidad, se realizaron Pruebas de Kruskal-Wallis (Steel & Torrie 1992) para determinar si los puntos pertenecientes a una misma transecta se diferencian del resto por su posición sobre los ejes. Se calculó la Correlación por Rangos de Spearman (Steel & Torrie 1992) entre la posición de los puntos sobre los dos primeros ejes del ordenamiento y los valores de superficie de bosque en torno del punto, longitud de borde bosque-pastizal, riqueza total de especies del punto y riqueza de especies para diferentes grupos biogeográficos y tróficos. Los grupos biogeográficos se definieron a partir de las Provincias Ornitogeográficas (Narosky & Yzurieta 1987) en las que se distribuyen las especies: (1) especies de distribución geográfica restringida a regiones boscosas como las provincias del Chaco, Espinal o Paranense, y (2) especies de distribución geográfica amplia, que además de las anteriores incluyeron a las provincias Pampeana, del Monte o Patagónica. Los grupos tróficos analizados correspondieron a las especies que incluyeron insectos como ítem principal de su dieta, las que incluyeron frutos y las que incluyeron granos. A su vez, se diferenció entre especies que obtienen su alimento en los estratos bajos y el suelo, en el follaje arbóreo, en troncos y por encima del dosel arbóreo. Algunas especies fueron incluidas en más de un grupo debido a la amplitud de su dieta y comportamiento de forrajeo. La caracterización trófica se realizó a partir de los datos de Juárez (1995) y de observaciones personales de los autores.
Se realizó un Análisis de Correlación para evaluar el efecto de la superficie de bosque y la longitud del borde bosque-pastizal sobre la riqueza de especies. Mediante Correlación Parcial (Steel & Torrie 1992) se evaluó la contribución del borde no relacionada con el área (i.e., el efecto del borde manteniendo constante el área). Tales correlaciones se realizaron para el total de especies así como por separado para los grupos tróficos y biogeográficos considerados.
RESULTADOS
Los autovalores de los primeros dos ejes del Análisis de Componentes Principales de los puntos basado en la abundancia de las especies de aves representaron un total de 21% de la varianza total [varianza explicada para el eje 1: 4.43 (11.9%); varianza explicada para el eje 2: 3.29 (8.9%)]. Los puntos pertenecientes a una misma transecta no presentaron valores similares de posición sobre los ejes (Eje 1: H = 5.15, n = 48, P > 0.39; Eje 2: H = 0.33, n = 48, P > 0.06), indicando que la variación en la composición dentro de las transectas es similar a la variación entre las mismas y que los puntos de una misma transecta pueden considerarse observaciones independientes.
A pesar del bajo valor de varianza explicada, el ordenamiento de los puntos permitió obtener un primer eje que se correlacionó con el área de bosque en torno del punto y con la longitud de borde bosque-pastizal (Tabla 1). Se detectaron 11 y 12 especies cuya abundancia aumentó hacia el extremo negativo y positivo del primer eje, respectivamente (Tabla 2). La correlación entre la posición de los puntos sobre el eje y el área de bosque indicó que ambos grupos constituyen especies que responden positiva y negativamente a la cobertura arbórea, respectivamente. La longitud de borde también se correlacionó con el primer eje (Tabla 1) y estuvo positivamente asociada al área de bosque en torno de cada punto (r = 0.78, P < 0.01). El segundo eje de ordenamiento no se correlacionó con el área de bosque ni con la longitud de borde.
Tabla 1. Correlaciones por Rangos de Spearman (R) entre la posición de los puntos de conteo de aves sobre los primeros dos ejes de un Análisis de Componentes Principales y el área de bosque en torno del punto, la longitud de borde bosque-pastizal, la riqueza total de especies del punto y la riqueza de especies para diferentes grupos biogeográficos y tróficos, en talares del noreste de la provincia de Buenos Aires.
Table 1. Spearman´s Correlations (R) between the position of birdís point counts along first two axis from a Principal Component Analysis and forest area around the point, forest-grassland edge longitude, total species richness, and species richness for different biogeographic and trophic groups, in northeastern Buenos Aires Province forests.
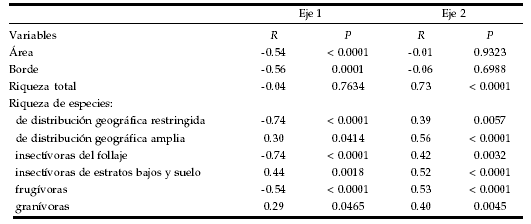
Tabla 2. Contribuciones de las variables (especies) a los primeros dos ejes del Análisis de Componentes Principales de los puntos de conteo de aves en talares del noreste de la provincia de Buenos Aires. Para cada especie se indican los grupos biogeográficos y tróficos a los que fue asignada. No se clasificaron los nectarívoros ni los insectívoros que se alimentan sobre troncos o por encima del dosel, debido al bajo número de especies representadas. B: especies de distribución restringida a regiones boscosas; NB: especies presentes en regiones no boscosas; IB: insectívoros de estratos bajos y suelo; IF: insectívoros que se alimentan en el follaje; FR: frugívoros; GR: granívoros; NA: especies no analizadas debido a su baja frecuencia.
Table 2. Factor loadings for variables (species) for the first two axis of the Principal Component Analysis on bird´s point counts in north-eastern Buenos Aires Province forests. Biogeographic and trophic groups in which species were included are indicated. Nectarivorous birds and insectivorous birds eating on branches or above the forest canopy were not classified due to the low number of species in these groups. B: species restricted to forest regions; NB: species not restricted to forest regions; IB: low-stratum and ground insectivores; IF: foliage insectivores; FR: frugivores; GR: granivores; NA: species not analized due to low occurrence.


El primer eje no se correlacionó con la riqueza total de especies pero sí con la riqueza de distintos grupos biogeográficos y tróficos (Tabla 1). Las tendencias resultaron opuestas entre especies con distribución más restringida a regiones boscosas y aquellas presentes en las provincias Pampeana, del Monte o Patagónica. Se halló una relación negativa con el primer eje para la riqueza de especies clasificadas como insectívoros del follaje y frugívoros, y positiva para insectívoros de los estratos bajos y el suelo. Se halló una tendencia positiva para las especies que consumen granos, aunque no resultó significativa con la corrección para pruebas múltiples (corrección de Bonferroni: a/18 = 0.0028). Debido a que algunas especies fueron incluidas en más de un grupo, esos resultados no fueron independientes entre sí. Sin embargo, algunos grupos con hasta 3 o 4 especies en común presentaron tendencias opuestas en su riqueza (Tabla 3), indicando que esos coeficientes estuvieron poco afectados por las especies comunes. El segundo eje se asoció a la riqueza total y a la calculada por grupos tróficos y biogeográficos. Todos los grupos presentaron la misma tendencia de cambio de la riqueza respecto del segundo eje del ordenamiento.
Tabla 3. Correlaciones de la riqueza de especies de los distintos grupos tróficos y biogeográficos con el área de bosque en cada punto de conteo de aves en talares del noreste de la provincia de Buenos Aires. La correlación parcial de la longitud de borde bosque-pastizal indica su asociación con la riqueza después de haber removido el efecto del área. Se indica el número de especies en común entre distintos grupos.
Table 3. Correlations between species richness for different trophic and biogeographic groups and forest area in birdís point counts in north-eastern Buenos Aires Province forests. Partial correlation between species richness and forest-grassland edge longitude indicates the association after removing forest area effect. Number of common species among groups is indicated.

Las relaciones entre la riqueza y el área de bosque resultaron coherentes con las halladas en el primer eje del ordenamiento (Tabla 3). La riqueza total de especies no mostró una relación significativa con el aumento del área de bosque. Sin embargo, al analizar por separado los diferentes grupos biogeográficos (Figura 1) y tróficos (Figura 2) se hallaron relaciones similares a las obtenidas en el primer eje de ordenamiento. El Análisis de Correlación Parcial indicó que no existe una relación entre la longitud de borde y esas variables que no quede explicada por la relación con el área de bosque (Tabla 3).

Figura 1. Relación entre la riqueza de especies y el área de bosque en torno de cada punto de conteo de aves en talares del noreste de la provincia de Buenos Aires para el grupo de especies con distribución geográfica restringida a regiones de bosques subtropicales (abajo) y con distribución geográfica más amplia (arriba).
Figure 1. Relationship between species richness and forest area around bird´s point counts in north-eastern Buenos Aires Province forests for species geographically restricted to subtropical forest regions (below) and species not restricted to forest regions (above).

Figura 2. Relación entre la riqueza de especies y el área de bosque en torno de cada punto de conteo de aves en talares del noreste de la provincia de Buenos Aires para grupos de especies que consumen diferentes recursos.
Figure 2. Relationship between species richness and forest area around bird´s point counts in northeastern Buenos Aires Province forests for species consuming different resources.
DISCUSIÓN
El principal eje de variación de la composición de la comunidad de aves estuvo asociado a las variaciones en la proporción de bosque. Esto indica que la estructura del paisaje constituiría una de las principales fuentes de variación de la abundancia de especies. Las áreas con mayor proporción de bosque corresponden a los puntos que se ubicaron sobre parches de bosque con dosel continuo, mientras que aquellas con menor proporción correspondieron a los puntos ubicados sobre bosques abiertos o parches menores de 0.5 ha aislados en pastizales. La escala espacial a la que fueron determinadas las caracterÌsticas del paisaje constituye otra fuente de variación. El área elegida fue de 4 ha en torno de cada uno de los puntos de muestreo de aves, que cubrían aproximadamente 0.13 ha. Las diferencias entre puntos con respecto al ·rea de bosque, de pastizal y a la longitud de borde podrÌan cambiar si cambia el área de medida de esas variables. Los resultados obtenidos permiten afirmar que existe una relación entre la estructura del paisaje y el principal eje de variación de la composición de especies de aves, pero se requiere de un trabajo de mayor detalle para determinar si un cambio de escala mejoraría el estudio de esa relación. La relación encontrada entre la riqueza total y el segundo eje de ordenamiento no estuvo asociada al área de bosque ni a la longitud de borde, por lo que podría deberse a diferencias estructurales entre los parches de bosque o a diferencias al nivel de paisaje a una escala diferente de la considerada en este trabajo.
La diversidad del conjunto de las especies no aumentó con el área de bosque. Esto se debió a la superposición de especies con tendencias opuestas en el conjunto analizado. Cuando se analizó por separado al grupo de especies que a priori se consideraron con afinidad por las áreas boscosas, sí se hallaron relaciones con la disponibilidad de bosque. Las tendencias opuestas de la riqueza de especies para grupos con diferentes características tróficas y biogeográficas a lo largo del gradiente de composición reflejan diferencias entre especies en la selección del hábitat. Las que responden positivamente a la cobertura de bosques dependen de la disponibilidad de insectos y frutos que obtienen en el dosel. Las que responden negativamente a la cobertura de bosques utilizan principalmente granos e insectos que obtienen en los estratos bajos y el suelo, en áreas con alta disponibilidad de pastizal. Además, estos grupos se diferenciaron por su distribución geográfica, aunque la mayoría de las especies se encuentra presente en las regiones Paranense, Chaqueña y Mesopotámica. La principal diferencia reside en que las especies asociadas a sitios con mayor cobertura arbórea presentan una distribución más restringida a áreas de bosques subtropicales, mientras que las restantes se distribuyen, además, por las regiones Pampeana, del Monte y, en algunos casos, Patagónica. Algunas especies de este último grupo presentan características tróficas claramente relacionadas con los pastizales, como la dieta granívora. Estudios realizados en el Hemisferio Norte demostraron que los migrantes neotropicales se encuentran entre los grupos más seriamente afectados por la fragmentación del bosque (Whitcomb et al. 1981; Ambuel & Temple 1983). En cambio, en los talares, dos migrantes de regiones de bosques tropicales estuvieron asociadas a bosque denso (Pachyramphus polychopterus y Elaenia parvirostris) y tres a bosques abiertos (Tyrannus savana, Tyrannus melancholicus y Pyrocephalus rubinus), sugiriendo que la condición de migrantes tropicales no permite predecir el tipo hábitat seleccionado.
La relación entre la diversidad y el área de hábitat disponible fue atribuida a diferentes causas. Según Rudnicky & Hunter (1993), la riqueza de aves se correlaciona positivamente con el área debido a un aumento en la heterogeneidad del hábitat y al requerimiento de área de determinadas especies en función de sus características tróficas. Además, los autores indican que pueden existir problemas metodológicos en el estudio de esa relación. Cuando el esfuerzo de muestreo es mayor en los parches más grandes aumenta la probabilidad de encontrar mayor número de especies que en los más pequeños. Las tendencias encontradas en este trabajo no pueden atribuirse a diferencias en el esfuerzo de muestreo, ya que todos los puntos fueron muestreados con la misma intensidad. Según Holmes et al. (1979), la estructura del bosque, caracterizada por la altura del dosel, la distribución de alimento en el tronco de los árboles y las características del follaje, se relaciona con la organización de la comunidad de aves en gremios tróficos. Por otra parte, MacArthur et al. (1962) concluyeron que la fragmentación de los bosques es el principal factor que afecta la diversidad de aves y que este efecto es mucho más importante que la complejidad de su estructura vertical (Roth 1976). En los talares, la heterogeneidad del bosque puede deberse a variaciones en las proporciones de las especies arbóreas dominantes y a variaciones estructurales debidas a la historia de uso (Goya et al. 1992). Cueto (1996) encontró que esas variaciones estructurales pueden afectar la composición de especies y la proporción de grupos tróficos. Las tendencias encontradas en este trabajo se corresponden en parte con lo afirmado por MacArthur et al. (1962), ya que las diferencias estructurales entre parches no fueron tenidas en cuenta y aún así se detectó el efecto del área. Sin embargo, también se espera que la heterogeneidad estructural se correlacione con el tamaño de parche, por lo que ambos efectos podrían superponerse. Los resultados son más concluyentes con respecto al efecto del borde bosque- pastizal. El incremento del borde es uno de los principales efectos de la fragmentación del hábitat, aunque su incidencia sobre la diversidad ha sido objeto de discrepancias entre los distintos autores (Rudnicky & Hunter 1993). Algunos trabajos encontraron una relación positiva entre esas variables (Strelke & Dickson 1980; Hansson 1983), mientras que otros no hallaron asociación entre ellas (Elliot 1987; Small & Hunter 1989; Rudnicky & Hunter 1993). Según Murcia (1995), los efectos producidos por los bordes de los fragmentos pueden ser abióticos o bióticos. En el caso de los talares, el borde de los fragmentos de bosque presenta condiciones de mayor iluminación que el interior y, consecuentemente, mayor disponibilidad de hojas, flores, frutos, así como de invertebrados asociados a esos recursos. Esas diferencias podrían resultar en variaciones de la abundancia de diferentes especies de aves entre el interior y el borde de los fragmentos. Podría esperarse que en sitios con similar proporción de bosque la diversidad sea mayor en el que presenta mayor longitud de borde en virtud de la mayor heterogeneidad de hábitat. Sin embargo, el borde no aportó información diferente de la del área en las relaciones con la diversidad.
La respuesta de las especies a la disponibilidad de hábitat también puede deberse a limitaciones de área. Tal causa fue señalada por Rudnicky & Hunter (1993) como causante de la relación área-diversidad, debido a que algunas especies requieren grandes áreas de alimentación por el tipo de alimento consumido. Algunas especies de bosque denso de los talares solo están presentes en los sitios con más de 50% de cobertura arbórea. Sin embargo, no presentan un tipo de alimentación que sugiera la necesidad de grandes áreas de alimentación, ya que se trata de insectívoros del follaje (Parula pitiayumi y Pachyramphus polychopterus). Su restricción a las áreas con mayor disponibilidad de bosque puede responder a la ventaja conferida por una mayor relación entre el área de bosque y la distancia recorrida, disminuyendo la relación costo-beneficio.
Las relaciones área-diversidad halladas indican que la conservación de los talares en función de las especies de aves debería contemplar los diferentes requerimientos de los dos grupos encontrados. En coincidencia con Cueto (1996), se encontró un grupo de especies asociadas a los bosques que están biogeográficamente relacionadas con bosques subtropicales, pero se halló, además, un grupo asociado a bosques abiertos. Las áreas con mayor diversidad corresponden a los sitios con más de 50% de cobertura arbórea y a aquellos con alrededor de 5%, independientemente de la estructura del bosque y de la proporción de borde de los parches. Tales situaciones maximizan la probabilidad de encontrar a las especies asociadas a bosque denso y bosque abierto, respectivamente.
BIBLIOGRAFÍA
AMBUEL, B & SA TEMPLE. 1983. Area-dependent changes in the bird communities and vegetation of southern Wisconsin forest. Ecology 64:1057-1068. [ Links ]
BLAKE, JG. 1983. Trophic structure of bird communities. Condor 82:149-158. [ Links ]
BLAKE, JG & JR KARR. 1984. Species composition of bird communities and the conservation benefit of large versus small forests. Biol. Conserv. 30:173-187. [ Links ]
BLAKE, JG. 1987. Species-area relationships of winter residents in isolated woodlots. Wilson Bull. 99:243-252. [ Links ]
BURGESS, RL & DM SHARPE. 1981. Forest island dynamics in man-dominated landscapes. Springer- Verlag. New York. [ Links ]
CUETO, VR. 1996. Relación entre los ensambles de aves y la estructura de la vegetación. Un análisis a tres escalas espaciales. Tesis Doctoral, Universidad de Buenos Aires, Buenos Aires. [ Links ]
CUETO, VR & J LOPEZ DE CASENAVE. 2000a. Bird assemblages of protected and exploited coastal woodlands in east-central Argentina. Wilson Bull. 112:395-402. [ Links ]
CUETO, VR & J LOPEZ DE CASENAVE. 2000b. Seasonal changes in bird assemblages of coastal woodlands in east-central Argentina. Stud. Neotrop. Fauna Environ. 35:173-177. [ Links ]
DÍAZ, M; R CARBONELL; T SANTOS & JL TELLERÍA. 1998. Breeding bird communities in pine plantations of the Spanish plateaux: biogeography, landscape and vegetation effects. J. Appl. Ecol. 35:562-574. [ Links ]
DIGBY, PGN & RA KEMPTON. 1987. Multivariate analysis of ecological communities. Chapman & Hall. Londres. [ Links ]
ELLIOT, CA. 1987. Songbird species diversity and habitat use in relation to vegetation structure and size of forest stands and forest-clearcut edge in northcentral Maine. Dissertation, University of Maine, Orono. [ Links ]
FREEMARK, KE & HG MERRIAM. 1986. Importance of area and habitat heterogeneity to bird assemblages in temperate forest fragments. Biol. Conserv. 36:115-141. [ Links ]
GALLI, A; C LECK & R FORMAN. 1976. Avian distribution in forest islands of different sizes in central New Jersey. Auk 93:356-364. [ Links ]
GOYA, JF; LG PLACCI; MF ARTURI & AD BROWN. 1992. Distribución y características estructurales de los talares de la Reserva de Biosfera "Parque Costero del Sur". Revista de la Facultad de Agronomía La Plata 68:53-64. [ Links ]
HARRIS, LD. 1984. The fragmented forest. University of Chicago Press. Chicago. [ Links ]
HANSSON, L. 1983. Birds numbers across edges between mature conifer forest and clearcuts in central Sweden. Ornis Scand. 14:97-103. [ Links ]
HOLMES, RT; RE BONNEY, JR & SW PACALA. 1979. Guild structure of the Hubbard Brook bird community: a multivariate approach. Ecology 60:512-520. [ Links ]
JUÁREZ, MC. 1995. Estatus de residencia y categorización trófica de las aves en la reserva natural isla Martín García, río de La Plata superior. Neotropica 41:83-88. [ Links ]
MARTIN, TE. 1980. Diversity and abundance of spring migratory birds using habitats islands on the Great Plains. Condor 82:430-439. [ Links ]
MARTIN, TE. 1981. Limitation in small habitat islands: chance or competition? Auk 98:715-734. [ Links ]
MARTIN, TE. 1988. Processes organizing open-nesting bird assemblages: competition or nest predation? Evol. Ecol. 2:37-50. [ Links ]
MCARTHUR, RH; JW MCARTHUR & J PREER. 1962. On bird species diversity. II. Prediction of bird census from habitat measurements. Am. Nat. 96:167-174. [ Links ]
MURCIA, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends Ecol. Evol. 10:58-62. [ Links ]
NAROSKY, T & D YZURIETA. 1987. Guía para la identificación de las aves de Argentina y Uruguay. Asociación Ornitológica del Plata. Buenos Aires. [ Links ]
RALPH, CJ; GR GEUPEL; P PYLE; T MARTIN; DF DESANTE & B MILÁ. 1996. Manual de métodos de campo para el monitoreo de aves terrestres. Gen. Tech. Rep. PSW- 159, USDA Forest Service. Albany. [ Links ]
ROTH, RR. 1976. Spatial heterogeneity and bird species diversity. Ecology 57:773-782. [ Links ]
RUDNICKY, T & M HUNTER, JR. 1993. Reversing the fragmentation perspective: effects of clearcut size on bird species richness in Maine. Ecol. Appl. 3:357-366. [ Links ]
SMALL, MF & ML HUNTER, JR. 1989. Response of passerines to abrupt forest-river and forest-powerline edges in Maine. Wilson Bull. 101:77-83. [ Links ]
SOAVE, GE; G MARATEO; P REY; D GLAZ & C DARRIEU. 1999. Evolución estacional de los ensambles de aves en un talar del nordeste de la provincia de Buenos Aires, Argentina. Comisión de Investigaciones Científicas. La Plata. [ Links ]
STEEL, RG & JH TORRIE. 1992. Bioestadística: principios y procedimientos. 2da edn. McGraw-Hill, México DF. [ Links ]
STRELKE, WK & JG DICKSON. 1980. Effect of forest clear-cut edge on breeding birds in east Texas. J. Wildlife Manage. 44:559-567. [ Links ]
WHITCOMB, RF; JF LYNCH; MK KLIMKIEWICZ; CS ROBBINS; BL WHITCOMB & D BYSTRAK. 1981. Effects of forest fragmentation on avifauna of the eastern deciduous forest. Pp. 125-205 en: L Burgess & DM Sharpe (eds). Forest island dynamics in man-dominated landscapes. Springer-Verlag. New York. [ Links ]
WILLIAMSON, M. 1981. Island populations. Oxford University Press. Oxford. [ Links ]
WILLIS, EO. 1979. The composition of avian communities in remnant woodlots in southern Brazil. Papeis Avulsos de Zoologia 33:1-25. [ Links ]

