










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.13 n.2 Córdoba jul./dic. 2003
TRABAJOS ORIGINALES
Relative abundance and autecology of three piscivorous fishes in lakes of the upper basin of Salado River (Buenos Aires Province, Argentina)
María J Barla1,*, María S Vera2 & Emma D O'Brien2
1. Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Argentina
2. Departamento de Biología, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina
* Fac. de Ciencias Naturales y Museo. Univ. Nac. de La Plata. Buenos Aires. Argentina. Dirección postal: calle 13 Nº 384 (1896) City Bell. Email: barla@netverk.com.ar
Recibido: 26 de enero de 2002;
Fin de arbitraje: 8 de abril de 2002;
Primera revisión recibida: 30 de abril de 2002;
Segunda revisión recibida: 25 de octubre de 2002;
Versión final recibida: 3 de junio de 2003;
Aceptado: 3 de junio de 2003
ABSTRACT. The Salado River is an important watercourse in northern Pampean region of Argentina which includes several lakes in the upper part of its basin. This upper part shows lesser fish species richness than the lower section of the river. In this paper, an analysis of community structure of secondary and tertiary consumers and its autecology are presented. Among the 14 fish species recorded, 3 were mainly or totally piscivorous as adults. They constituted 11.4% of the numeric abundance and 10.8% of the biomass. Their numeric abundance was 89% (Oligosarcus jenynsii), 11% (Rhamdia sapo), and 0.5% (Hoplias malabaricus malabaricus), and their relative biomass was 61%, 33%, and 6%, respectively. Length/weight ratio, condition factor, and cephalic index indicate that the three populations presented normal development. Stomachs were classified according to the degree of repletion. Differences in repletion were attributable to the feeding behaviour of the different species. The relative abundance differed from the one observed in other environments within the same basin. This variation could be the result of interspecific competition or of differences in resource availability.
Palabras clave: Characiformes; Siluriformes; Predation; Biocenology; Lentic environments: Pampean lakes; Trophic levels.
RESUMEN. Peces piscívoros en lagunas de la cuenca superior del río Salado (Buenos Aires, Argentina: abundancia relativa y autoecología: El objetivo de este trabajo fue analizar la estructura de la comunidad de peces consumidores secundarios y terciarios, considerando aspectos autoecológicos que dieran cuenta del estado de estas poblaciones, en las lagunas de la cuenca superior del río Salado. Se registraron 14 especies de peces, de las cuales 3 eran depredadores piscívoros que representaban un 11.4% de la abundancia numérica y un 10.8% de la biomasa. La abundancia relativa de este nivel trófico fue de 89% (Oligosarcus jenynsii), 11% (Rhamdia sapo) y 0.5% (Hoplias malabaricus malabaricus). Considerando la biomasa, el 61% correspondió a Oligosarcus jenynsii, el 33% a Rhamdia sapo y el 6% a Hoplias malabaricus malabaricus. La relación longitud-peso, el factor de condición y el índice cefálico indicaron un desarrollo normal de las poblaciones de estas tres especies. Se clasificaron los estómagos de las mismas de acuerdo a la cantidad de alimento, observándose diferencias en el grado de repleción atribuibles al comportamiento alimentario de las especies. La abundancia relativa difirió de la encontrada en otros ambientes de la misma cuenca. Esta variación podría atribuirse a la competencia interespecífica o a las diferencias en la disponibilidad de recursos.
Keywords: Characiformes; Siluriformes; Depredación; Biocenología; Ambientes lénticos; Lagunas pampásicas; Niveles tróficos.
INTRODUCTION
Predation, together with resource availability, competition and environmental modifications, is one of the main factors determining community structure (Bechara 1993). Certainly, predation could affect fish species of recreational or commercial importance. The Pampean lakes of central-eastern Argentina include several fish species of economic interest. Many of these lakes occur in the Salado River basin, extending on part of northern, center and eastern Buenos Aires Province.
The Brazilian Subregion of the Neotropical Region is one of the richest areas of fish diversity in the world (Ringuelet 1975; Böhlke et al. 1978). The Brazilian diversity dramatically diminishes south of the Río de la Plata River. Fish species richness of Salado River basin is 24 species (Barla 1991) but, according to Aurand (2000), only 14 species were recorded in the upper section. Among the predatory fishes present in this part of the basin, we studied three species: Oligosarcus jenynsii (Günther), Rhamdia sapo (Valenciennes), and Hoplias malabaricus malabaricus (Bloch). These species are piscivorous in different degree, at least in part of the life cycle (Destéfanis & Freyre 1972; Haro & Gutiérrez 1985; Oliveros & Rossi 1991; Aquino 1993; Hartz et al. 1996). It is possible that both Rhamdia sapo and Hoplias malabaricus integrate a particular trophic guild (sensu Aranha et al. 1998) of mainly piscivorous fishes, whereas Oligosarcus jenynsii integrate a trophic guild of a wider carnivorous diet.
This paper provides an analysis of the ecology of tertiary (and in part secondary) consumers in lakes of the Pampean region, as a part of a research project related to the control of eutrophication.
Study Area
Pampean lakes of Argentina range from several tens of meters in diameter to 2-3 km, rarely being deeper than 4 m. Lake basins are mostly below 100 m.a.s.l. They are usually mature or highly eutrophicated. Water level varies according to rains, tributaries and groundwater level. Changes in precipitation regime frequently produce floods or droughts. Predominance of evaporation in closed basins in the arid areas of the Pampean region provokes increases in solute concentration and rising of salinity, sometimes bringing about dystrophic states (Ringuelet 1968). Besides, hydrochemical properties, plankton composition, and general metabolism greatly change according to local modification of climate, geomorphology, and substratum. The Pampean lakes have different origins: transformation of coastal lagoons; deflation processes; sand dunes, loessic dunes, or shell accumulation forming dams; valleys filled with macrophytes; and tectonism (Frenguelli 1956; Ringuelet 1972). The important changes in the sea level during the late Pleistocene and Holocene (negative and positive) greatly influenced the valleys morphology (Tonni & Cione 1997). During the sea level drops river valleys probably rejuvenated.
Lakes studied herein (Mar Chiquita, Gómez, Carpincho, and Rocha) are interconnected and located in the upper part of Salado River in northwestern Buenos Aires Province. They integrate the "Sistema del río Salado y sus afluentes" (Frenguelli 1956), which runs SENW and is to the south of Paraná River course. The Salado River flows into in Bahía de Samborombón. The lakes are located between 34º27´-34º35´S and 60º54´-61º11´W, in Junín and Chacabuco departments (Figure 1). They seem to have originated in fluvial depressions and meanders and by shrinking of a large marshy area (Frenguelli 1956). Precipitation feed surface drainage and the underground water locally.
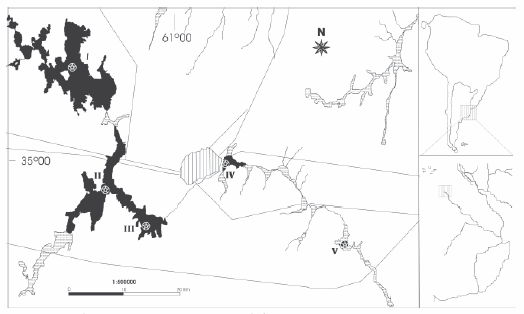
Figure 1. Sampling sites in the upper basin of Salado River, Buenos Aires Province, Argentina. I: Mar Chiquita lake, II: northern Gómez lake, III: eastern Gómez lake, IV: Carpincho lake, V: Rocha lake.
Figura 1. Sitios de estudio en la cuenca superior del río Salado, Buenos Aires, Argentina. I: Laguna de Mar Chiquita, II: Laguna de Gómez Norte, III: Laguna de Gómez Este, IV: Laguna del Carpincho, V: Laguna de Rocha.
Mar Chiquita lake occurs upstream of the lakes chain (34º25´S; 61º10´W); its surface is 5280 ha, and its mean depth is about 1.5 m. It flows into by means of a channel to Gómez lake (34º37´S; 61º07´W). This latter has a surface of 3660 ha, a mean depth of about 1.1 m, and is located at 75 m.a.s.l. Carpincho lake (34º35´S; 60º54´W) is connected with Gómez lake by means of an artificial channel. Its surface is about 440 ha and its mean depth 1.2 m. Rocha lake, which is downstream (34º42´S; 60º43´W), near the city of Chacabuco, has a surface of about 400 ha and a mean depth of 1.4 m.
According to Köppen´s temperature zones, the Pampean area is located in the Temper ate zone (4-12 months between 10-20°C; Miller 1965). The lake area here studied is located in the Subhumid Subzone of the Plains Temperate Climatic Zone (Iglesias de Cuello 1981). During the late Cenozoic glaciations, most of the Pampean region had a colder and drier climate, similar to that of northeastern Patagonia (Tonni et al. 1999). Most (if not all) Pampean lakes were formed in the Late Pleistocene or Holocene.
METHODS
Fishes were obtained monthly, from March 2000 to March 2001, in four sampling sites: one in Mar Chiquita lake, two in Gómez lake, and another in Carpincho lake. Additional sampling was carried out in one site in Rocha lake during two summers (March 2000, January to March 2001; Figure 1). Sampling sites were mostly in the pelagic zone of the lakes.
Samples were obtained with three different fishing gears: trawling nets, gill nets, and fishing-lines. The coastal trawlnet (see characteristics in Nédélec 1975) had 15 m of floatline, a funnel of 2.2 m, and square mesh of 9.71 mm. The gill nets consist of two series, each with 10 nets with different square mesh (15, 21, 25, 30, 35, 40, 52, 60, 75, 80 mm). They were installed between 20-24 h. Temperature and conductivity were measured with digital thermometer and conductimeter. Samples obtained by means of trawling nets and fishing-lines were not included in global biocenologic analyses, but they were employed for the autecological ones.
Specimens were identified, counted, and weighted to the nearest 0.1 g. For each species, total, fork, standard, and cephalic lengths were measured. Sex was determined by internal examination of gonads. For each monthly sample the relative abundance in number and weight of the three predatory species was calculated.
For each species, sex ratio and length/weight ratio were calculated. A normal approximation of the Chi-square Test (Zar 1996) was employed to test differences among male-female proportions between sites in Oligosarcus jenynsii and Rhamdia sapo. From the length/weight ratio, the condition factor was calculated (Le Cren 1951), discriminating sex and sample sites. The condition factor data for Oligosarcus jenynsii and Rhamdia sapo were first tested for normality and homogeneity of variance using the Kolmogorov-Smirnov Goodness of Fit Procedure and the Bartlett´s Test, respectively (Zar 1996). The existence of significant differences between the four seasons of the year for the condition factor was established by a Single-factor Analysis of Variance. Not planned contrasts were performed for testing the differences between the four seasons on Oligosarcus jenynsii data. For assessing the repletion degree, stomachs were classified visually according to the food content: replete, full, almost full, almost empty, and empty.
RESULTS AND DISCUSSION
Biocenology
In Mar Chiquita, Gómez, and Carpincho lakes, the three predators constituted 11.43% of the numeric abundance and 10.77% of the biomass. Rocha lake was not included in this analysis because of the short sampling period involved. The proportion (in number) of piscivorous fishes per species was 88.51% (Oligosarcus jenynsii), 11.04% (Rhamdia sapo), and 0.45% (Hoplias malabaricus malabaricus). The proportion in biomass was 60.39% (Oligosarcus jenynsii), 33.24% (Rhamdia sapo), and 6.37% (Hoplias malabaricus malabaricus) (Figure 2). When Rocha lake was included, the biomass of Hoplias malabaricus malabaricus reached 11%.

Figure 2. Relative abundance in number (N) and weight (W) of Oligosarcus jenynsii, Rhamdia sapo, and Hoplias malabaricus malabaricus in sampling sites in the upper basin of Salado River, Buenos Aires Province, Argentina, during the study period.
Figura 2. Abundancia relativa en número (N) y en peso (W) de Oligosarcus jenynsii, Rhamdia sapo y Hoplias malabaricus malabaricus en sitios de muestreo en la cuenca superior del río Salado, Buenos Aires, Argentina, durante el período de estudio.
When summer abundances in the five sites were compared, Hoplias malabaricus malabaricus was more abundant in Rocha lake, whereas Rhamdia sapo was less numerous and presented a lower biomass because it was represented only by juveniles (Figure 3). Hoplias malabaricus malabaricus is more abundant during the summer in this area (Ringuelet et al. 1967).
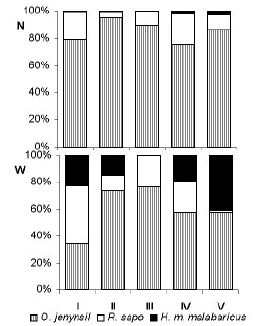
Figure 3. Relative abundance in number (N, above) and weight (W, below) of Oligosarcus jenynsii, Rhamdia sapo, and Hoplias malabaricus malabaricus in sampling sites in the upper basin of Salado River, Buenos Aires Province, Argentina, during summer. Site codes are the same as in Figure 1.
Figura 3. Abundancia relativa en número (N, arriba) y en peso (W, abajo) de Oligosarcus jenynsii, Rhamdia sapo y Hoplias malabaricus malabaricus en sitios de muestreo en la cuenca superior del río Salado, Buenos Aires, Argentina, durante el verano. Los códigos de los sitios son los mismos que en la Figura 1.
Autecology: Oligosarcus jenynsii
In all sites, 1419 individuals were caught. Standard length ranged from 57-202 mm, and weight ranged from 3.3-207.9 g. Maximum standard length recorded in other sites were 220 mm (Río Tercero reservoir, central Argentina; Iwaszkiw et al. 1983), 224 mm (San Roque reservoir, central Argentina; Haro & Gutiérrez 1985), 171.4 mm (El Cadillal reservoir, northern Argentina; Aquino 1993), and 264 mm (Lagoa Caconde, Río Grande do Sul, Brazil; Hartz et al. 1996).
Sex ratio.- Male numbers were higher in Mar Chiquita and Gómez lakes (Figure 4). Differences between Carpincho and Mar Chiquita, between northern and eastern Gómez, and between eastern Gómez and Rocha were significant (P < 0.05). In other sites, sex ratios were 69.3% for males and 30.7% for females (San Roque reservoir; Ocaña et al., unpubl. data), 66.3% for females, 28% for males and 5.7% for juveniles (El Cadillal reservoir; Aquino 1993), and 46.6% for males and for 53.4% females (Lagoa Caconde; Hartz et al. 1996).

Figure 4. Proportion of males, females, and immatures of Oligosarcus jenynsii, Rhamdia sapo, and Hoplias malabaricus malabaricus in sampling sites in the upper basin of Salado River, Buenos Aires Province, Argentina, during the study period. Site codes are the same as in Figure 1.
Figura 4. Proporción de machos, hembras e inmaduros de Oligosarcus jenynsii, Rhamdia sapo y Hoplias malabaricus malabaricus en sitios de muestreo en la cuenca superior del río Salado, Buenos Aires, Argentina, durante el período de estudio. Los códigos de los sitios son los mismos que en la Figura 1.
Length/weight ratio.- This ratio, discriminated by sex, age and sample site, shows that individuals were well developed. In all cases, the power curve was significant at P < 0.01, and the slope was about 3 (Table 1). We calculated the length/weight ratio for specimens caught in Río Tercero reservoir, central Argentina, using standard length and weight data for 26 females published by Iwaszkiw et al. (1983). The slope was similar: 3.1606 (R2 = 0.9793).
Table 1. Power equations fitted to the length/weight ratios in males, females and immatures of Oligosarcus jenynsii (O.j.) and Rhamdia sapo (R.s.) in sampling sites in the upper basin of Salado River, Buenos Aires Province, Argentina. Site codes are the same as in Figure 1.
Tabla 1. Ecuaciones potenciales ajustadas a la relación longitud/peso para machos, hembras e inmaduros de Oligosarcus jenynsii y Rhamdia sapo en sitios de muestreo en la cuenca superior del río Salado, Buenos Aires, Argentina. Los códigos de los sitios son los mismos que en la Figura 1.

Condition factor.- It was about 1, varying slightly in the different sampling sites, and indicating that the individuals presented a normal development (Table 2). Values were highest in summer and autumn, both for males and females. Data did not deviate from the requirements of normality and homoscedasticity. Significant differences were found between summer and autumn (F = 6.2297, P < 0.05). In Lagoa Caconde, Hartz et al. (1996) reported higher values in autumn and winter.
Table 2. Seasonal values (mean ± SD) of the condition factor for males, females and immatures of Oligosarcus jenynsii, Rhamdia sapo and Hoplias malabaricus malabaricus in the upper basin of Salado River, Buenos Aires Province, Argentina.
Tabla 2. Valores estacionales (promedio ± DE) del factor de condición para machos, hembras e inmaduros de Oligosarcus jenynsii, Rhamdia sapo y Hoplias malabaricus malabaricus en la cuenca superior del río Salado, Buenos Aires, Argentina.

Repletion degree.- Food was found in 51% of the stomachs (Figure 5). Aquino (1993) reported the same proportion for El Cadillal reservoir. We related the stomach content to water temperature (Figure 6), but it appears that there were not significant changes throughout the year, although full stomachs were more frequent during spring and summer.
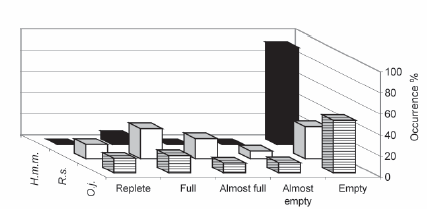
Figure 5. Degree of stomach repletion in Oligosarcus jenynsii (O.j.), Rhamdia sapo (R.s.), and Hoplias malabaricus malabaricus (H.m.m.) in the upper basin of Salado River, Buenos Aires Province, Argentina.
Figura 5. Grado de repleción estomacal en Oligosarcus jenynsii (O.j.), Rhamdia sapo (R.s.) y Hoplias malabaricus malabaricus (H.m.m.) en la cuenca superior del río Salado, Buenos Aires, Argentina.

Figure 6. Percentage of stomachs with food in Oligosarcus jenynsii, Rhamdia sapo and Hoplias malabaricus malabaricus, and water temperature in the upper basin of Salado River, Buenos Aires Province, Argentina, during the study period.
Figura 6. Porcentaje de estómagos con alimento en Oligosarcus jenynsii, Rhamdia sapo y Hoplias malabaricus malabaricus, y temperatura del agua en la cuenca superior del río Salado, Buenos Aires, Argentina, durante el período de estudio.
Autecology: Rhamdia sapo
In all sites, 178 individuals were caught. Standard length ranged from 43-421 mm, and weight ranged from 1.3-1925 g.
Sex ratio.- The proportion of immature individuals was higher in Carpincho and Rocha lakes (Figure 4). There were no significant differences between sampling sites.
Length/weight ratio.- In all cases, the power curve was significant at P < 0.01, and the slope was about 3 (Table 1). In Lagoa Paranoá (Pernambuco, Brazil), an analysis for a different species of the genus Rhamdia showed a slope of 3.05 (R2 = 0.947; Ribeiro Filho 1999).
Condition factor.- Data did not deviate from the requirements of normality and homoscedasticity. No significant differences F = 1.82419, P > 0.05) were found between seasons. In both males and females, condition factor was about 1 (Table 2), indicating that individuals presented a normal development. It was slightly lower in immature individuals during winter.
Repletion degree.- Food was found in 69% of the stomachs (Figure 5). Ribeiro Filho (1999) reported food in 76.5% of the stomachs in Lagoa Paranoá. According to this author, the local species of the genus Rhamdia feeds continuously, what results in a higher rate of stomachs with food. In the studied Pampean lakes, 100% of the stomachs presented food during autumn (Figure 6). This autumn was relatively warm.
Autecology: Hoplias malabaricus malabaricus
In all sites, only 13 individuals were caught. Standard length ranged from 235-435 mm, and weight ranged from 325.7-2275 g.
Sex ratio.- There were more males in Rocha lake, where there was a higher catch (Figure 4).
Length/weight ratio.- This rate was not significant due to the scarce number of individuals caught, excepting males when the five sites were considered altogether. In this case, the rate is Weight = 5 10-6. Lenght 3.26, and it is significant at P < 0.05. The fit obtained for females was Weight = 4 10-5. Lenght 2.92, although it was not significant. In San Miguel del Monte lake (Buenos Aires Province), the same subspecies showed a length/weight ratio of Weight = 8.1019 10-6. Lenght 3.1908 (Dománico 1998). The slope was 2.87 (R2 = 0.87) in Cabra Corral reservoir (Salta Province; Barros et al., unpubl. data), 2.97 (R2 = 0.92) in Brotas reservoir (Pernambuco, Brazil; Ribeiro Filho 1999), and 3.19 (R2 = 0.98) in Lagoa Paranoá (Brazil; Ribeiro Filho 1999).
Condition factor.- In both males and females, condition factor was about 1 (Table 2), indicating that individuals presented a normal development.
Repletion degree.- Food was found in only one stomach (7.7%; Figure 5). In rivers and lakes of Santa Fe Province, Oliveros & Rossi (1991) found 47% of stomachs with food, taking all sizes. Ribeiro Filho (1999) reported food in 88% of the stomachs in Pernambuco (Brazil), and 69.9% in Lagoa Paranoá. He considered that these figures were unexpected, because Hoplias malabaricus malabaricus is an ambush predator that waits quietly under cover until suitable prey comes close (Ringuelet et al. 1967). Besides, Parma de Croux (1983) mentions that adults of Hoplias malabaricus malabaricus diminish predatory activity and metabolic rate. Ramírez (1963) reported that Hoplias malabaricus malabaricus strongly reduces its feeding in winter. Moreover, Ringuelet et al. (1967) suggest that this fish is dormant during the colder months. In the studied Pampean lakes, the sole stomach with food was full, and the specimen was caught at the end of the winter, when temperature was steeply rising (Figure 6). The absence of food in most of the samples could be related to the lack of juveniles of this species, and to the peculiar predatory habit and metabolism of adults.
The analyzed parameters suggest a normal development of the three species, and are in agreement with data from other environments. Adult individuals of these species eat mainly fish and, secondarily, crustaceans and other relatively small animals (Ringuelet et al. 1967; Haro & Gutiérrez 1985; Oliveros & Rossi 1991; Aquino 1993; Hartz et al. 1996; Ribeiro Filho 1999). Notwithstanding the different morphology, size, feeding strategies and habitat, their trophic niches are probably overlapping. The coexistence of these species could be guaranteed possibly by the different feeding strategies. The percentage of available high-quality energy transferred from one trophic level to another varies from 2-30%, depending on the type of species involved and the ecosystem in which the transfer takes place (Miller 1990). An average of about 10% of the high-quality chemical energy available at one trophic level is typically transferred and stored in useful form as chemical energy in the bodies of the organisms at the next level.
We suggest that Hoplias malabaricus malabaricus is a top-predator in the lakes here studied. The low quality and quantity of energy that reaches this trophic level possibly restrict this species (bottom-up effect). However, it could be limited by the possible competition with Rhamdia sapo. This competition is suggested by (1) the distribution of catfish sizes, and (2) the fact that in these upper Salado River basin lakes Hoplias malabaricus malabaricus population is much less abundant than that of Rhamdia sapo, which is opposite of what is observed in the lower Salado River basin lakes (Barla 1990, 1991). Zoogeographical features may likely explain this. There are biodiversity differences, which decrease toward the west (Ringuelet 1975).
The possible overlap of trophic niches should be studied through qualitative and quantitative analyses of diet, to determine if presence and abundance of the different piscivorous species are constrained by interspecific competition, resource availability, or other environmental factors.
ACKNOWLEDGEMENTS
We acknowledge to Laboratorio de Sistemas de Producción Acuática (Facultad de Agronomía, Universidad de Buenos Aires), where we carried out the research, with partial support from the Agencia Nacional de Investigaciones Científicas y Tecnológicas (Grant PICT 04698 to R Quirós and MJ Barla), and the Consejo Nacional de Investigaciones Científicas y Técnicas. We are also grateful to the Municipality of Junín that provided assistance for the fieldwork. We thank to Laboratorio de Sistemas de Producción Acuática for the use of data obtained during the development of the project (especially to A Renella for temperature and conductivity data) as well as to other people that participated in the project. We also thank AL Cione for assistance in the preparation of a former draft of the manuscript, and R Sarandón for his help with some statistical topics.
REFERENCES
AQUINO, AE. 1993. Hábitos alimentarios de Oligosarcus jenynsii (Pisces, Characidae) en el embalse El Cadillal (Tucumán, Argentina). Acta Zoologica Lilloana 42:267-277. [ Links ]
ARANHA, JMR; D TAKEUTI & TM YOSHIMURA. 1998. Habitat use and food partitioning of the fishes in a coastal stream of Atlantic Forest, Brazil. Trop. Biol. 46:951-959. [ Links ]
AURAND, S. 2000. Estudio comparativo de las comunidades de peces de las lagunas de Gómez y del Carpincho (verano 1998-1999). Tesis de grado, Facultad de Agronomía, Universidad de Buenos Aires, Buenos Aires. [ Links ]
BARLA, MJ. 1990. Estructura ecológica de la comunidad íctica de una laguna pampásica. Tesis doctoral, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba. [ Links ]
BARLA, MJ. 1991. Species composition, richness and diversity of fish assemblages in different habitats of a Pampean lake (Argentina). Ann. Limnol. 27:163-173. [ Links ]
BECHARA, JA. 1993. El papel de los peces en el control de la estructura de las comunidades bénticas de los ecosistemas lóticos. Pp. 143-158 in: A Boltovskoy & HL López (eds). Conferencias de Limnología. Instituto de Limnología "Dr. Raúl A. Ringuelet". La Plata. [ Links ]
BÖHLKE, JE; SH WEITZMAN & NA MENEZES. 1978. Estado atual da sistematica de peixes da agua doce da America do Sul. Amazonica 8:657-677. [ Links ]
DESTÉFANIS, S & L FREYRE. 1972. Relaciones tróficas de los peces de la laguna Chascomús con un intento de referenciación ecológica y tratamiento bioestadístico del espectro trófico. Acta Zoologica Lilloana 29:17-33. [ Links ]
DOMÁNICO, AA. 1998. Age and growth of Hoplias malabaricus malabaricus (Bloch, 1794) from San Miguel del Monte Lagoon (Argentine) and comparison with other lentic environments of pampasic plains. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" 8:31-41. [ Links ]
FRENGUELLI, J. 1956. Rasgos generales de la hidrografía de la provincia de Buenos Aires. Publicaciones del LEMIT 62:1-19. [ Links ]
HARO, JG & M GUTIÉRREZ. 1985. Alimentación de Oligosarcus jenynsii (Günther) (Pisces, Characidae) en el Lago San Roque (Córdoba, Argentina). Revista de la Asociación de Ciencias Naturales del Litoral 16:227-235. [ Links ]
HARTZ, SM; A MARTINS & G BARBIERI. 1996. Dinamica da alimentacao e dieta de Oligosarcus jenynsii (Gunther, 1864) na lagoa Caconde, Río Grande do Sul, Brasil (Teleostei, Characidae). Boletim do Instituto de Pesca 23:21-29. [ Links ]
IGLESIAS DE CUELLO, A. 1981. Tipos de clima. Pp. 193-200 in: E Chiozza & R Figueira (eds). Atlas total de la República Argentina. Centro Editor de América Latina. Buenos Aires. [ Links ]
IWASZKIW, JM; L FREYRE & ED SENDRA. 1983. Estudio de la maduración, época de desove y fecundidad del dientudo Oligosarcus jenynsii (Pisces Characidae) del Embalse Río Tercero, Córdoba, Argentina. Limnobios 2:518-525. [ Links ]
LE CREN, ED. 1951. The length weigth relationship and seasonal cycle in gonad and condition in perch (Perca fluviatilis). J. Anim. Ecol. 20:201-219. [ Links ]
MILLER, AA. 1965. Climatology. Methuen. Londres. [ Links ]
MILLER JR, GT. 1990. Resource conservation and management. Wadsworth. Belmont. [ Links ]
NÉDÉLEC, C. 1975. Catalogue of small-scale fishing gear. Fishery Industries Division, FAO. West Byfleet. [ Links ]
OLIVEROS, OB & LM ROSSI. 1991. Ecología trófica de Hoplias malabaricus malabaricus (Pisces, Erythrinidae). Revista de la Asociacón de Ciencias Naturales del Litoral 22:55-68. [ Links ]
PARMA DE CROUX, MJ. 1983. Metabolismo de rutina de Hoplias malabaricus (Bloch, 1974) (Osteichthyes, Erythrinidae). Rev. Bras. Zool. 1:217-222. [ Links ]
RAMÍREZ, FC. 1963. Contribución al conocimiento de la Tararira (Hoplias malabaricus malabaricus) de la Laguna de Chascomús (Prov. de Buenos Aires). Notas del Museo de La Plata, Zoología 20:141-158. [ Links ]
RIBEIRO FILHO, RA. 1999. Caracterizaçao de dietas alimentares de peixes predadores em açudes de Pernambuco e no lago Paranoa (Brasilia-DF). Tesis, Departamento de Pesca, Universidade Federal Rural de Pernambuco, Pernambuco. [ Links ]
RINGUELET, R. 1968. Tipología de las lagunas de la Provincia de Buenos Aires. La Limnología regional y los tipos lagunares. Physis 28:65-76. [ Links ]
RINGUELET, R. 1972. Ecología y biocenología del hábitat lagunar o lago de tercer orden de la Región Neotrópica Templada (Pampasia Sudoriental de la Argentina). Physis 31:55-76. [ Links ]
RINGUELET, R. 1975. Zoogeografía y Ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur 2:1-122. [ Links ]
RINGUELET, R; R ARAMBURU & AA DE ARAMBURU. 1967. Los peces argentinos de agua dulce. Comisión de Investigaciones Científicas de la Provincia de Buenos Aires. La Plata. [ Links ]
TONNI, EP & AL CIONE. 1997. Did the Argentine Pampean ecosystem exist in the Pleistocene? Current Research in the Pleistocene 14:131-133. [ Links ]
TONNI, EP; AL CIONE & A FIGINI. 1999. Predominance of arid climates indicated by mammals in the pampas of Argentina during the late Pleistocene and Holocene. Palaeogeogr. Palaeocl. 147:257-281. [ Links ]
ZAR, JH. 1996. Biostatistical analysis. 3rd edn. Prentice Hall. Upper Saddle River. [ Links ]

