










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.14 n.1 Córdoba ene./jun. 2004
TRABAJOS ORIGINALES
Fluctuación estacional en la infestación de diversos hospedadores por la mosca del Mediterráneo, Ceratitis capitata (Diptera: Tephritidae), en la provincia de Buenos Aires
Diego F Segura*, M Teresa Vera & Jorge L Cladera
Instituto de Genética "E. A. Favret", INTA Castelar, Buenos Aires, Argentina
*Instituto de Genética "E. A. Favret", INTA Castelar. CC 25, Castelar (1712), Buenos Aires, Argentina. Email: dsegura@cnia.inta.gov.ar
Recibido: 2 de octubre de 2002;
Fin de arbitraje: 16 de enero de 2003;
Revisión recibida: 24 de abril de 2003;
Versión final recibida: 24 de junio de 2003;
Aceptado: 24 de junio de 2003
RESUMEN. Se realizó un relevamiento de 11 especies frutales hospedadoras de Ceratitis capitata presentes en un área experimental de San Pedro (Buenos Aires, Argentina), con una extensión aproximada de 115 ha. Se comparó la abundancia de la población de insectos adultos y el nivel de infestación de las especies frutales (inferido a partir del porcentaje de fruta infestada y del número de puparios/kg de fruta) a lo largo de una temporada cálida, y se analizó el efecto de algunas variables bióticas y abióticas sobre las fluctuaciones poblacionales. El estudio comprendió muestreos quincenales realizados entre noviembre de 1999 y mayo de 2000. En los mismos se recolectaron frutos maduros de 20 plantas tomadas al azar. Los periodos de fructificación de las especies presentes ocurrieron en forma escalonada a lo largo de la temporada reproductiva, asegurando a los individuos un suministro de recurso casi ininterrumpido. No obstante, la disponibilidad global de hospedadores no estuvo asociada con las fluctuaciones poblacionales. Probablemente el efecto restrictivo de las bajas temperaturas de invierno impide que la población alcance los niveles necesarios para explotar el recurso en forma proporcional a su disponibilidad. Se encontró un alto grado de asociación entre la abundancia de adultos y los niveles de infestación. En el área de estudio, el único factor climático que parece tener un efecto sobre estas variables es la temperatura. Las correlaciones de la abundancia de adultos y de los niveles de infestación con la temperatura del período precedente resultaron más altas que con las temperaturas del período corriente, indicando que este factor climático probablemente afecta en forma más intensa a los estadios inmaduros que a los adultos.
Palabras clave: Moscas de los frutos; Fluctuaciones poblacionales; Niveles de infestación.
ABSTRACT. Seasonal fluctuation on infestation of several hosts by the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae) in Buenos Aires Province: A survey of 11 host species of Ceratitis capitata was carried out in an experimental area of San Pedro (Buenos Aires, Argentina) with an approximate extension of 115 ha. The abundance of adult flies was compared with the infestation level (inferred from the percentage of infested fruit and from the number of puparia/kg of fruit) along the reproductive season. Also, the effect of some biotic and abiotic factors on the population fluctuations was analyzed. The study included 12 fortnightly samplings carried out between November 1999 and May 2000. In each sampling, mature fruits of 20 plants were taken at random. For each fruit, the number of puparia of Ceratitis capitata was recorded. The fruiting periods of the host species present in the area were staggered along the reproductive season, assuring to the flies a continuous supply of hosts. Nonetheless, population fluctuations were not related to the availability of hosts. The restrictive effect of low winter temperatures probably prevents population from reaching levels required to exploit the resource in proportion to its availability. There was a high association between the abundance of adult flies and infestation levels. In the study area, temperature was the only climatic factor showing significant association with the abundance of Ceratitis capitata and infestation levels. Correlations of the abundance of adult flies and of the infestation levels with the temperature of the previous period were higher than the correlation with the temperature of the current period, probably indicating that this climatic factor has a stronger effect on the immature stages than on the adults.
Keywords: Population fluctuations; Infestation levels.
INTRODUCCIÓN
La mosca del Mediterráneo, Ceratitis capitata (Wiedemann, 1824), es una de las plagas de los frutales más importante del mundo debido a su gran capacidad destructiva y a la gran diversidad de especies que puede utilizar como hospedadores. De hecho, más de 350 especies han sido listadas como hospedadores para esta plaga (Liquido et al. 1990). En Argentina genera pérdidas de hasta el 20% de la producción frutícola (Alvarado, INTA Castelar, com. pers.), pero el mayor daño económico es indirecto y se debe a la pérdida de mercados potenciales para las exportaciones debido a restricciones cuarentenarias (Malavasi et al. 1994). Existen aspectos básicos de la ecología de esta especie que, además de presentar interés científico, poseen interés aplicado, debido a su utilidad en programas de control o en la implementación de procedimientos cuarentenarios (Fletcher 1986). Un ejemplo son los estudios que buscan determinar cuáles son los factores bióticos y abióticos que afectan la abundancia de esta plaga, por cuanto permiten estimar los periodos más proclives a un aumento de los números poblacionales de la plaga por encima de los niveles de daño económico.
Entre los factores bióticos que afectan la abundancia, la relación entre la disponibilidad de especies frutales y las fluctuaciones poblacionales de Ceratitis capitata es uno de los más estudiados (Vergani 1961; Vargas et al. 1983; Harris & Lee 1986, 1987; Harris et al. 1993; Mazih & Debouzie 1996; Katsoyannos et al. 1998; Debouzie & Mazih 1999; Mavrikakis et al. 2000; Papadopoulos et al. 2001). Estos trabajos muestran que las fluctuaciones poblacionales están asociadas solo a algunos de los hospedadores presentes (hospedadores clave). Para tener un efecto significativo sobre las fluctuaciones poblacionales, un hospedador debe ser muy abundante y tener una alta capacidad de carga (sensu Lack 1954); i.e., poseer capacidad de soportar altos niveles de infestación (Vargas et al. 1983). Sin embargo, la presencia de diversas especies, incluyendo hospedadores no claves, en una secuencia de fructificación escalonada, también juega un papel importante. Cuando esta secuencia es interrumpida, la población desciende hasta niveles de abundancia muy bajos (Puzzi & Orlando 1965; Malavasi & Morgante 1981).
Para estudiar la interacción entre Ceratitis capitata y sus plantas hospedadoras en un área particular se debe realizar un estudio previo sobre las especies hospedadoras encontradas en ella (Malavasi et al. 1980; Malavasi & Morgante 1980; Boscán de Martinez 1993; Putruele 1996; de Coll, INTA Montecarlo, com. pública), así como un seguimiento de su fenología, especialmente de su periodo de fructificación (Aluja 1999). En general, también se analizan los niveles de infestación verificados en cada hospedador. La observación que se desprende de estos estudios es que una misma especie hospedadora parece no tener la misma susceptibilidad al ataque de Ceratitis capitata en distintos escenarios ecológicos. Pero estos niveles medios de infestación suelen resultar de un promedio de varias fechas distintas. Teniendo en cuenta que las hembras encuentran un panorama distinto a lo largo del tiempo, ya que la situación cambia continuamente a medida que la abundancia de los distintos hospedadores fluctúa, un promedio entre distintas fechas dificulta la interpretación de los resultados a nivel del hospedador.
Entre los factores abióticos que afectan las fluctuaciones poblacionales, la temperatura (fundamentalmente la invernal) es el principal en la región Mediterránea, particularmente en Grecia (Katsoyannos et al. 1998; Papadopoulos et al. 2001). En cambio, en zonas subtropicales la abundancia de Ceratitis capitata está principalmente afectada por las precipitaciones y la humedad relativa (Harris & Lee 1987). En estas zonas, la temperatura fluctúa dentro de los niveles óptimos y, por lo tanto, no representa un factor limitante para el establecimiento y la persistencia de la población (Bateman 1972). En Marruecos tanto la falta de humedad como la temperatura afectan las fluctuaciones poblacionales de Ceratitis capitata (Mazih & Debouzie 1996). En la Patagonia argentina, la abundancia y diversidad de hospedadores aseguran la supervivencia poblacional solo cuando las temperaturas resultan favorables (Rial, INTA Alto Valle, com. pública). En el noreste de la provincia de Buenos Aires, el factor climático más importante en relación con las fluctuaciones de la abundancia de Ceratitis capitata es la temperatura (Segade & Polack, INTA San Pedro, com. pública). Vergani (1961) sostiene que inviernos templados seguidos de veranos húmedos y cálidos aseguran un ciclo biológico corto y una proliferación abundante. En consecuencia, las provincias del noroeste y noreste argentino, que cuentan con este tipo de clima, resultan muy favorables para el desarrollo de la plaga. Sin embargo, hay que tener en cuenta que condiciones climáticas extremas (elevada temperatura ambiente, elevada o muy baja humedad y abundantes precipitaciones) también pueden imponer restricciones al crecimiento poblacional, y que estos factores extremos se presentan en varios lugares del noroeste y del noreste (Vera et al. 2002).
MÉTODOS
Área de estudio
El estudio se realizó en la Estación Experimental Agropecuaria del Instituto Nacional de Tecnología Agropecuaria (INTA) de San Pedro, al noreste de Buenos Aires. Dicha estación presenta una extensión aproximada de 115 ha en las cuales se desarrollan distintas actividades agrícolas. Cuenta con plantaciones experimentales de cítricos (naranja y mandarina), durazno y ciruela, dispuestas en distintos lotes. Dentro del mismo predio se encuentran otras especies frutales cultivadas en lotes de menor tamaño (higo, caqui, kiwi), así como plantas de falsa guayaba. El clima de la región es templado con dos períodos bien definidos: uno frío (desde mayo hasta septiembre) y otro cálido (desde noviembre hasta marzo). La temperatura promedio del período frío es de 12.0ºC y la del período cálido de 21.9ºC. La humedad relativa promedio anual es de 76% y las precipitaciones oscilan entre 900-1100 mm anuales, distribuidos principalmente en el período cálido (Torroba et al. 1968). En esta área se han registrado dos especies de moscas de los frutos: Ceratitis capitata y Anastrepha fraterculus (Wiedemann, 1830). Sin embargo, en la actualidad Anastrepha fraterculus se encuentra en niveles no detectables por el sistema de trampeo dispuesto en la zona (Segade & Polack, INTA San Pedro, com. pública). No se realiza control químico mediante uso de insecticidas en el área de estudio.
Muestreo
Se realizaron 12 muestreos entre los meses de noviembre de 1999 y mayo de 2000, separados entre sí por un lapso de 2 semanas. En cada muestreo se tomaron 20 unidades muestrales (i.e., plantas). El muestreo fue de tipo estratificado al azar; las unidades muestrales se dividieron en estratos homogéneos y, dentro de éstos, las unidades muestrales fueron seleccionadas al azar. Cada especie hospedadora fue considerada un estrato diferente. El número de estratos de los cuales se tomaron muestreas fue variable, puesto que dependía del número de especies hospedadoras que presentaran frutos maduros en cada muestreo. Para determinar el número de unidades muestrales dentro de cada estrato se realizó una asignación proporcional de muestras, dividiendo el número total de plantas a muestrear (n = 20) y asignando a cada estrato un número proporcional al total de plantas presentes en él.
En cada muestreo se recorrió previamente el área de estudio y se registró el estado fenológico de las plantas en cada estrato. Se seleccionaron todos aquellos estratos en que existían plantas con frutos susceptibles de ataque por moscas de la fruta y se asignó la cantidad de unidades muestrales en cada uno de ellos. Luego se procedió a seleccionar las unidades muestrales aleatoriamente dentro de cada estrato. En cada una de ellas se registró la cantidad de frutas que se encontraban en el suelo justo por debajo del área de cobertura. Se muestrearon 10 frutos por planta, tomando únicamente frutos desde la planta y cuidando que los frutos seleccionados no mostraran signos de que alguna larva hubiera abandonado el fruto. Las muestras fueron transportadas al INTA Castelar, ubicado aproximadamente a 170 km del área de estudio, donde fueron pesadas y colocadas en recipientes individuales, conteniendo un fondo de arena que es utilizado por las larvas como sustrato de pupación.
La arena contenida en los recipientes fue tamizada semanalmente para recuperar los puparios y se registró el número de puparios obtenidos. Éstos fueron colocados en recipientes individuales y mantenidos hasta la emergencia del adulto, para su posterior identificación. En aquellos casos en que no emergió el adulto, los puparios se preservaron en alcohol para su posterior identificación. Las muestras fueron descartadas luego de dos revisiones seguidas en las que no se produjeron puparios, siempre que hubieran sido revisadas al menos en tres ocasiones luego de su entrada.
Abundancia de adultos
Simultáneamente con el muestreo, el Laboratorio de Sanidad Vegetal de la EEA San Pedro realizó un seguimiento de la abundancia relativa de la población mediante un sistema de trampeo de adultos (Segade, INTA San Pedro, com. pers.). Se utilizaron trampas Jackson (cebadas con trimedlure, que atrae exclusivamente a machos) y trampas Portici (cebadas con un atrayente alimenticio líquido que atrae a ambos sexos). Las trampas fueron colocadas en la copa de plantas hospedadoras seleccionadas al azar dentro de la EEA. La frecuencia de revisión de las trampas fue distinta a la frecuencia de muestreo de frutos. Para estimar la abundancia de adultos en cada fecha de muestreo de frutos se calculó, para cada tipo de trampa en cada fecha de revisión, el número promedio de moscas caídas por trampa por día. Luego se estimó la abundancia de adultos en cada fecha de muestreo como la superficie por debajo de la curva de moscas por trampa por día, comprendida entre las fechas previa y actual de muestreo.
Análisis de datos
Para describir las fluctuaciones en la diversidad de hospedadores con frutos maduros se calculó el índice de diversidad específica de Shannon (Begon et al. 1990). El índice tiene en cuenta el número de especies presentes y la abundancia relativa de cada una de ellas. La abundancia relativa de cada hospedador fue obtenida a partir del peso estimado de fruta madura. Para esto se tuvo en cuenta el peso promedio de los frutos de cada especie y la producción total de frutos de cada especie, estimada a partir del número de árboles de cada especie y de la producción de cada árbol, obtenida del recuento de la fruta al pie de los árboles muestreados.
La abundancia poblacional de puparios fue obtenida a partir de la sumatoria (incluyendo todas las especies frutales) del producto entre el peso estimado de fruta madura de cada especie frutal y el número promedio de puparios/kg encontrado en cada especie frutal.
El nivel de infestación fue estimado a través del porcentaje de frutas infestadas (número de frutas infestadas/número de frutas muestreadas x 100) y del número de puparios/kg de fruta. En ambos casos se calculó el promedio poblacional para cada muestreo como el promedio aritmético de los valores para cada fruta (no fue necesario ponderar por la cantidad de plantas de cada especie porque se muestreó en forma proporcional a la misma).
La disponibilidad de cada hospedador en cada muestreo fue estimada a partir de la abundancia de cada especie (en kg de fruta producida) y de su calidad relativa como hospedador. La calidad de un hospedador fue estimada a partir de la relación entre la capacidad de carga (i.e., número máximo de puparios/kg) de ese hospedador y la capacidad de carga máxima registrada entre todos los hospedadores. De esta manera, la calidad toma valores entre 0 y 1, y carece de unidades. En todos los casos, la capacidad de carga se calculó como el percentil 90 de la distribución de valores del número de puparios/kg para frutas que tuvieron un número de puparios distinto de cero. La disponibilidad total del recurso se estimó, entonces, como la suma de las disponibilidades registradas en cada una de las especies presentes.
Para estudiar el grado de asociación entre puparios totales, nivel de infestación, adultos en trampa y diversas variables bióticas y abióticas, se realizó una serie de análisis de correlación. Las variables abióticas utilizadas fueron: temperatura máxima, promedio y mínima, temperatura del suelo a 5, 10 y 20 cm de profundidad, humedad relativa y precipitación. Para todas las variables, excepto para precipitación, se consideró como valor representativo del muestreo al promedio de los 14 días previos a éste (i.e., al período comprendido entre dos muestreos sucesivos). En el caso de la precipitación, se consideró la sumatoria de los valores diarios para el mismo período de tiempo. Se repitieron los mismos análisis, pero con los valores de las variables abióticas correspondientes al promedio o a la sumatoria (dependiendo del caso) de los 14 días previos al muestreo anterior (indicados con el subíndice t-1).
En el caso de los puparios totales, la única variable biótica utilizada fue la disponibilidad total de hospedadores. En los análisis de los niveles de infestación (% de fruta infestada y número de puparios/kg), las variables bióticas fueron: nivel de adultos en trampa (Portici y Jackson) y disponibilidad total de hospedadores. Para los niveles de adultos en trampa, las variables bióticas usadas fueron la abundancia de puparios en el mismo período de tiempo (puparios totales), en el período precedente (puparios totales t-1) y en el anterior a éste (puparios totales t-2).
RESULTADOS
Fenología de los hospedadores
Se encontraron 11 especies hospedadoras de Ceratitis capitata: caqui (Dyospiros kaki L. f.) (30 plantas), ciruela (Prunus insititia L.) (700 plantas), durazno (Prunus persica L.) (5063 plantas), falsa guayaba (Feijoa sellowiana O. Berg) (2 plantas), higo (Ficus carica L.) (106 plantas), kiwi (Actinidia chinensis Planch.) (17 plantas), mandarina (Citrus reticulata Blanco) (822 plantas), manzana (Malus domestica Borkh) (45 plantas), naranja (Citrus sinensis (L.) Osbeck) (830 plantas), nectarina (Prunus persica var. nucipersica L.) (72 plantas) y pera asiática (Pyrus pyrifolia (Burn. F.) Nakai) (74 plantas). Tres especies mostraron un periodo de fructificación interrumpido: naranja y nectarina (ausentes la primera fecha de diciembre), y mandarina (ausente en la segunda quincena de febrero y la primera de marzo) (Figura 1). Para los análisis subsiguientes, se decidió separar a la especie mandarina en dos variedades: mandarina 1 (noviembre-febrero) y mandarina 2 (marzo-mayo). Naranja y nectarina no mostraron datos que justificaran un análisis separado.
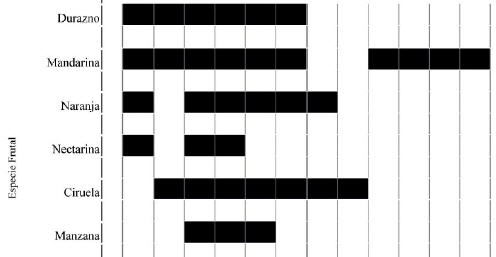

Figura 1. Fechas de muestreo (entre noviembre de 1999 y mayo de 2000) en las cuales las distintas especies hospedadoras presentaron fruta susceptible de ataque por moscas de los frutos en el noreste de la provincia de Buenos Aires.
Figure 1. Sampling dates (November 1999 to May 2000) in which the different host species bore mature fruits, susceptible of attacks by fruit flies in north-eastern Buenos Aires Province.
Las variaciones en la composición de especies con fruta madura estuvieron acompañadas por fluctuaciones en la abundancia relativa de cada especie. Estos cambios determinaron notables fluctuaciones en la diversidad de fruta disponible a lo largo del tiempo (Figura 2). Hubo muestreos en los que la diversidad fue especialmente baja (muestreos 4, 8, 11 y 12) o alta (muestreo 6). Las caídas en la diversidad específica estuvieron asociadas con la preponderancia de tres especies: durazno (muestreos 1-5), caqui (muestreos 7 y 8) y mandarina 2 (muestreos 9-12), que aparecieron como especies dominantes en distintos momentos (Figura 2). Al mismo tiempo, el valor relativamente alto del índice en el muestreo 6 se correspondió con un número de especies relativamente elevado ("otros" incluye en esa fecha a las especies mandarina 1, naranja, ciruela, higo y pera asiática) y a una abundancia similar entre las mismas (i.e., no hubo una especie dominante).
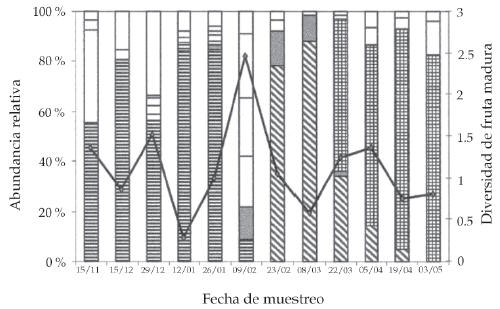
Figura 2. Abundancia relativa de las distintas especies hospedadoras (barras) y diversidad de fruta madura (índice de Shannon) (línea continua), para cada fecha de muestreo, en el noreste de la provincia de Buenos Aires. La categoría "otros" incluye al resto de los hospedadores presentes y se muestra desagregada para poder explicar las fluctuaciones en el índice. Las especies son: durazno (líneas horizontales), caqui (líneas oblicuas), higo (gris), mandarina 2 (cuadriculado), otras (blanco).
Figure 2. Relative abundance of the different host species (bars) and diversity of mature fruit (Shannon index) (continuous line) for each sampling date in north-eastern Buenos Aires Province. The category "others" includes the rest of the hosts, and it was disaggregated in order to explain the fluctuations in the index. Host species are peach (horizontal lines), persimmon (oblique lines), fig (grey), tangerine 2 (squared), and others (white).
Moscas de la fruta encontradas
En todas las especies frutales muestreadas se encontraron puparios de Ceratitis capitata, mientras que solo se recuperaron puparios de Anastrepha fraterculus en durazno, falsa guayaba, caqui y mandarina. La incidencia de Anastrepha fraterculus en el área de estudio fue muy baja: se recuperaron 13 puparios de esta especie contra 18017 de Ceratitis capitata y no existieron registros de la primera en los relevamientos realizados a través de trampas. Debido a la baja representación de Anastrepha fraterculus, los análisis se realizaron únicamente para Ceratitis capitata.
Variación temporal de la población
El número total de puparios mostró inicialmente valores muy bajos, aumentando bruscamente hasta alcanzar un máximo aproximado de 3.106 puparios en enero (Figura 3). En el muestreo siguiente, la producción global de puparios cayó y, a partir de esa fecha, se produjo un aumento leve hasta un nuevo máximo en marzo (aproximadamente 2.105 puparios), a partir del cual cayó definitivamente. La población de adultos se mantuvo inicialmente baja y creció rápidamente a partir de enero hasta alcanzar un máximo en febrero, luego del cual la población disminuyó hasta niveles muy bajos, aumentando levemente hacia mayo (Figura 4). El porcentaje de fruta infestada mostró valores bajos durante noviembre y diciembre, aumentando rápidamente a partir de enero hasta alcanzar un máximo en la primera quincena de marzo (Figura 5). Luego se produjo una caída muy brusca y, finalmente, un leve aumento hacia el final de la temporada. El número de puparios/kg mostró una dinámica semejante: un aumento gradual hasta un máximo en febrero, luego una caída rápida y, finalmente, un pequeño aumento (Figura 5).

Figura 3. Número total de puparios, para cada fecha de muestreo, en el noreste de la provincia de Buenos Aires. En el recuadro se muestra el detalle del segundo máximo poblacional encontrado.
Figure 3. Total number of puparia for each sampling date in north-eastern Buenos Aires Province. The box inside the figure shows the detail of the second population peak found.
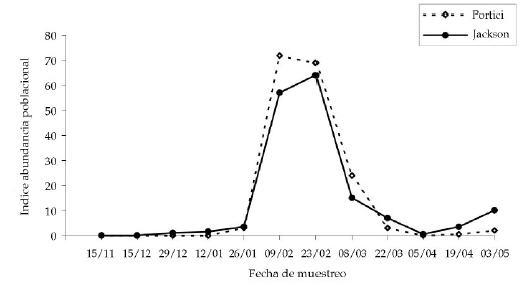
Figura 4. Índice de abundancia de adultos en trampas tipo Portici y Jackson (Segade, com. pers.), para cada fecha de muestreo, en el noreste de la provincia de Buenos Aires.
Figure 4. Abundance of adults in Portici and Jackson traps (Segade, pers. comm.) for each sampling date in north-eastern Buenos Aires Province.

Figura 5. Porcentaje de fruta infestada y número de puparios/kg, para cada fecha de muestreo, en el noreste de la provincia de Buenos Aires. En ambos casos se presenta el promedio (± EE) entre todas las unidades muestrales tomadas en cada muestreo.
Figure 5. Percentage of infested fruit and number of puparia/kg of fruit for each sampling date in north-eastern Buenos Aires Province. Averages (± SE) among all the sampling units are shown.
Niveles de infestación por hospedador
Tanto el porcentaje de fruta infestada como el número de puparios/kg mostraron una gran heterogeneidad entre las especies frutales en los valores máximos alcanzados a lo largo del periodo de muestreo (Tabla 1).
Tabla 1. Valores máximos del porcentaje de frutas infestadas y del número de puparios/kg de fruta registrado para cada especie hospedadora en el noreste de la provincia de Buenos Aires. Entre paréntesis se indica la fecha de muestreo en que ocurrió el máximo.
Table 1. Maximum values of percentage of infested fruit and number of puparia/kg of fruit for each host in north-eastern Buenos Aires Province. Sampling date in which that maximum happened is shown between parentheses.

Las diferencias entre especies hospedadoras en cuanto al porcentaje de frutas infestadas y al número de puparios/kg no siempre se tradujeron en diferencias semejantes en la contribución de cada hospedador al número de puparios totales, puesto que la abundancia de cada hospedador fue variable (Figura 6). Entre diciembre y enero, el durazno fue el hospedador que más puparios aportó al número total de puparios; luego pasó a tomar su lugar el caqui y, en menor grado, el higo. Al final, quedaron como principales fuentes de puparios la falsa guayaba, el caqui y la mandarina 2.

Figura 6. Contribución relativa de las distintas especies hospedadoras a la producción total de puparios, para cada fecha de muestreo, en el noreste de la provincia de Buenos Aires.
Figure 6. Relative contribution of the different host species to the total production of puparia for each sampling date in north-eastern Buenos Aires Province.
Análisis de Correlación
La abundancia de adultos, estimada mediante trampas Portici, se correlacionó significativamente con la temperatura mínima t-1 y las tres medidas de temperatura del suelo t-1, mientras que la abundancia de adultos estimada mediante trampas Jackson se correlacionó con la temperatura del suelo t-1 a 10 cm y 20 cm de profundidad (Tabla 2). Asimismo, ambas medidas de abundancia de adultos mostraron una correlación significativa con la abundancia de puparios t-1 y la abundancia de puparios t-2. El porcentaje de fruta infestada mostró correlaciones significativas con la temperatura mínima, con la temperatura mínima t-1 , con la temperatura del suelo y con la temperatura del suelo t-1 a las tres profundidades. El porcentaje de fruta infestada también estuvo correlacionado con la abundancia de adultos estimada por ambos métodos. El número de puparios/kg presentó asociaciones significativas con la temperatura mínima t-1 y la temperatura del suelo t-1 para las tres profundidades, y con la abundancia de adultos estimada a partir de los dos tipos de trampas (Tabla 2). No se encontraron correlaciones significativas entre el número total de puparios y las variables bióticas y abióticas analizadas.
Tabla 2. Valores del Coeficiente de Correlación que fueron significativos (P < 0.05) entre los índices relativos de densidad poblacional y diversas variables abióticas, puparios totales e índices de caídas en trampa (Portici y Jackson). Todos los coeficientes significativos fueron positivos. ns: no significativo; -: combinaciones de variables no analizadas.
Table 2. Significant values (P < 0.05) of the Correlation Coefficient between population relative density indices and abiotic variables, total number of puparia or trap catches indexes (Portici and Jackson). All significant coefficient values were positive. ns: not significant; -: pair of variables not analyzed.

DISCUSIÓN
Este estudio permitió describir el patrón de maduración y abundancia de las especies hospedadoras de Ceratitis capitata presentes en un área que, por su alta diversidad de especies frutales, resulta particularmente interesante para el estudio de las interacciones entre esta plaga y sus hospedadores.
En esta área se encontraron once especies hospedadoras, cuyos periodos de fructificación ocurren en forma escalonada durante el período cálido que va desde noviembre hasta marzo. Ésta puede ser considerada como la temporada reproductiva para esta población, dado que el número total de puparios y la población de adultos se encuentran en niveles bajos (y en algunos casos no detectables) en los frutos que se muestrean antes y después de este periodo. La abundancia relativa de frutos maduros de cada especie fue heterogénea, pero se observó una clara predominancia de alguna de ellas (inicialmente durazno, luego caqui y finalmente mandarina 2). El escalonamiento en la maduración de especies hospedadoras le asegura a los individuos de Ceratitis capitata un suministro de recurso casi ininterrumpido. A lo largo de todo el año, el único momento en que se interrumpe la disponibilidad de fruta madura ocurre entre la caída de mandarina 2 y la maduración de otras especies de cítricos, como las naranjas de invierno que en el año del estudio ocurrió aproximadamente un mes después del último muestreo (y que no fueron incluidas). La existencia de hospedadores durante la mayor parte del año hace que esta área sea adecuada para la proliferación de la plaga durante la época reproductiva y para su supervivencia durante el invierno. De hecho, aún cuando las temperaturas bajas imponen condiciones restrictivas para los adultos y las pupas, Mavrikakis et al. (2000) han encontrado larvas y huevos de Ceratitis capitata en cítricos durante el invierno en ambientes templados, sugiriendo que la presencia de cítricos durante el invierno le permitiría a esta población perdurar en el área de estudio, quedando en niveles demasiado bajos como para ser encontrada.
Las fluctuaciones encontradas en la producción global de puparios muestran dos valores máximos. El primero resulta de la combinación de una alta abundancia de durazno y un grado de infestación también alto, mientras que el segundo resulta de los niveles de infestación y de la abundancia de caqui y, en menor grado, de higo. Es evidente que las fluctuaciones en la población de adultos son muy semejantes a las mostradas por el número de puparios totales, con una asociación significativa desfasada en el tiempo. Esto puede deberse a que cuando los individuos fueron muestreados en el campo estaban en estadio de huevo o larva (y no pupa), y transcurren aproximadamente 20 días desde que los estadios inmaduros alcanzan el estadio adulto (Putruele 1998).
Si bien era de esperar que la disponibilidad de hospedadores afectara el número de puparios, el Análisis de Correlación no detectó una asociación significativa. Esto implica que la disponibilidad de hospedadores no se traduce en una contribución neta al número de puparios, puesto que esta contribución depende de la proporción de recurso que están utilizando los individuos. De hecho, al realizar el mismo análisis pero omitiendo los cuatro primeros muestreos (en los que parece haber una alta asincronía) se encontró una asociación positiva y significativa entre la disponibilidad de hospedadores y el número de puparios (r = 0.999). Evidentemente, el efecto de las condiciones climáticas o de algún recurso en estado limitante (que no es la disponibilidad de hospedadores) están impidiendo que esta población explote en forma completa el recurso hospedador que se le presenta en los primeros muestreos.
Los máximos alcanzados por el número total de puparios se deben principalmente a unos pocos hospedadores, que actuarían como "hospedadores clave" de este sistema (i.e., hospedadores responsables de elevar las curvas poblacionales de las moscas). En este estudio uno de los hospedadores clave fue el durazno. Estudios previos realizados en zonas con características ecológicas semejantes al área de estudio también reconocen la importancia de esta especie como principal contribuyente a la población de pupas (Vergani 1961; Abbiati et al. 1989). El higo y el caqui también se mostraron como hospedadores clave en este sistema, aunque su menor abundancia hizo que su impacto sobre la población sea menor que el del durazno.
Las fluctuaciones observadas en los niveles de infestación responden principalmente a dos factores relacionados con los hospedadores: capacidad de carga (número máximo registrado de puparios/kg de fruta) y abundancia relativa. Los valores máximos de infestación están relacionados con la alta capacidad de carga y la abundancia relativa elevada del caqui y, en menor grado, del higo. Aun cuando el durazno muestra una alta capacidad de carga y abundancia, en promedio no muestra niveles altos de infestación. Esto probablemente se debe a que en los meses de noviembre, diciembre y enero, momento en que hay mayor abundancia de durazno maduro (susceptible al ataque por Ceratitis capitata), el nivel poblacional de adultos deCeratitis capitata es bajo. En las fechas de muestreo intermedias, la población de adultos alcanzó niveles más altos y la abundancia de hospedadores fue menor; entonces, se alcanzan niveles de infestación mayores, los que se verifican en los hospedadores con mayor capacidad de carga (en este caso el higo y el caqui). Asimismo, en el caso de la mandarina 2 se esperaría que los altos niveles de infestación encontrados en los muestreos previos a su maduración repercutieran fuertemente sobre este hospedador, sobre todo una vez que la abundancia de caqui e higo disminuye. Sin embargo, esto no fue lo que se observó, y la disminución en la población de adultos parece ser la responsable de ello. Esta falta de correlación entre los niveles de infestación y la disponibilidad global de hospedadores, y la asociación positiva que habría entre la infestación y los niveles de la población de adultos, se vieron reflejados en los resultados de las correlaciones (Coeficiente de Correlación no significativo en el primer caso y significativo en el segundo).
El nivel de adultos en trampa mostró una fuerte correlación con la temperatura mínima del período precedente, pero no con la temperatura mínima del intervalo de tiempo corriente. Teniendo en cuenta que la duración de los estadios inmaduros estimada para una temperatura promedio como la del área de estudio fluctúa entre 18-20 días (Putruele 1998), esta correlación podría estar indicando que la temperatura mínima se asocia al nivel de adultos debido a que afecta los estadios inmaduros (aumentando la mortalidad y el tiempo de desarrollo) que determinan, al menos en parte, el número de adultos encontrados en el siguiente muestreo. En este sentido, el impacto de la temperatura mínima sobre la población de adultos es indirecto. Las temperaturas registradas en el suelo (utilizado por las larvas como sustrato de pupación) mostraron una asociación significativa con los niveles de adultos del período subsiguiente, sugiriendo un efecto diferido sobre la abundancia de los adultos. Además, las asociaciones positivas entre la temperatura mínima y la temperatura del suelo y los niveles de infestación serían una consecuencia del efecto de la temperatura sobre los niveles de adultos, pues, como se discutió anteriormente, los adultos serían responsables, al menos en parte, de los niveles de infestación hallados (Figura 7).

Figura 7. Efecto propuesto de la temperatura sobre las fluctuaciones en la abundancia de adultos y los niveles de infestación (porcentaje de fruta infestada y numero de puparios/kg de fruta) durante el periodo de muestreo en el noreste de la provincia de Buenos Aires. Las fechas enteras señalan un efecto directo y las punteadas un efecto indirecto entre las variables; t0 y t1 representan periodos de muestreo sucesivos. El efecto de la temperatura sobre los estadios inmaduros viene dado por un aumento de la mortalidad por estrés térmico y porque las bajas temperaturas alargan el periodo de desarrollo (y así las larvas y pupas quedan expuestas a enemigos naturales por más tiempo).
Figure 7. Proposed effect of temperature on adult abundance fluctuations and on infestation levels (percentage of infested fruit and number of puparia/kg of fruit) during the sampling period in north-eastern Buenos Aires Province. Full arrows indicate direct influences of one variable on another, and dashed arrows indicate indirect effects between variables; t0 and t1 represent successive sampling dates. The effect of temperature upon immature stages comes from an increase in mortality caused by thermal stress and because low temperatures extend the duration of these stages, and the larvae and pupa remain exposed to their natural enemies.
El número total de puparios no mostró asociación con las variables climáticas analizadas, indicando que, en la comparación entre estadios inmaduros, la larva es menos afectada por la temperatura que la pupa. Esto concuerda con los resultados obtenidos por Mavrikakis et al. (2000), de cuyo trabajo se desprende que los frutos actuarían como un refugio debido a su capacidad de amortiguar las fluctuaciones en la temperatura ambiente, sobre todo cuando ésta toma valores muy bajos.
No se detectó en el área de estudio una asociación entre los niveles de adultos, de puparios y de infestación con las variables hídricas; el único factor climático que parece tener un efecto sobre esta población de Ceratitis capitata, al menos durante el periodo analizado, es la temperatura. Esto concuerda con trabajos realizados en áreas próximas al sitio de estudio (Segade & Polack, INTA San Pedro, com. pública) y con el patrón predicho para esta localidad (datos no publ.). Estas asociaciones sugieren que el área de estudio presenta un patrón similar al descripto en ambientes templados de la región mediterránea, donde el principal factor climático que estaría asociado a las fluctuaciones poblacionales es la temperatura relativamente baja de invierno (Katsoyannos et al. 1998; Papadopoulos et al. 2001).
El estudio comparado de las fluctuaciones en la población de adultos y de los estadios inmaduros con los valores instantáneos y desfasado en el tiempo de las variables climáticas brindó un panorama más informativo. En este caso, los análisis de correlación parecen indicar que el estado más sensible a la temperatura es el pupal, ya sea en forma directa, afectando su supervivencia, o diferida, a través de una disminución en la actividad de los adultos debida al estrés térmico sufrido en el estado pupal (Messenger & Flitters 1954; Meats 1989).
La naturaleza de la relación existente entre las moscas de los frutos y sus plantas hospedadoras, y el efecto que tienen las variables ambientales sobre esta relación son de fundamental importancia para comprender el proceso de expansión del número de hospedadores y para diseñar tácticas de detección y manejo efectivas (Prokopy, University of Massachussets, com. pública). A partir de este trabajo se pueden sacar algunas conclusiones que pueden aportar al manejo de la plaga en el sistema estudiado. Entre ellas, se puede proponer el cultivo de variedades más tempranas de durazno (que estarían más protegidas del daño causado por esta plaga por madurar cuando la plaga no se ha recuperado del efecto restrictivo de las bajas temperaturas de invierno) y la implementación de un sistema de control que integre tratamientos químicos realizados en momentos clave (e.g., en los momentos previos a los picos poblacionales) y en lugares clave (e.g., en las plantaciones de higo y caqui), con métodos de control mecánico como enterrar los frutos caídos, para evitar que las larvas aún presentes en ellos completen su desarrollo. La importancia de implementar medidas de control contra esta plaga se desprende claramente de los niveles de infestación, en los cuales puede verse que en un sistema sin control (como el estudiado) se pueden alcanzar pérdidas de hasta un 70% de la producción. Asimismo, el conocimiento de los principales factores climáticos asociados a las fluctuaciones poblacionales es importante para estimar cuáles son las condiciones ambientales que favorecen un aumento poblacional de la plaga hasta densidades superiores a los niveles de daño económico (Mavrikakis et al. 2000). No obstante, este trabajo tiene limitaciones que deberían ser subsanadas con otras observaciones. Entre estas deben incluirse: (1) el seguimiento de la población y de los niveles de infestación durante una cierta cantidad de años, para confirmar las tendencias encontradas y determinar la importancia de ciertos eventos estocásticos sobre este sistema; y (2) estudios de laboratorio en los que se evalúe no solo el grado de infestación alcanzado por ciertos hospedadores sino la elección de los individuos entre distintas especies, así como las tácticas que adoptan cuando los hospedadores no tienen la misma abundancia.
AGRADECIMIENTOS
Los autores desean agradecer a todo el personal de la EEA INTA San Pedro, en especial al Lic. G. Segade y al Ing. Agr. A. Polack por su constante apoyo en el área de estudio y por los valiosos aportes realizados a este trabajo, entre los cuales se encuentran los datos de adultos en trampa. Asimismo, deseamos agradecer al Ing. Agr. R. Rodríguez por permitirnos acceder a las bases de datos climáticos del Instituto de Clima y Agua, INTA Castelar, a la Dra. M. Busch por su colaboración en el diseño de este trabajo, por sus observaciones y por la lectura crítica de una versión previa de este manuscrito, y a la Ing. Agr. N. Kahn por el asesoramiento estadístico. Al Proyecto FONCyT PID 615 por el apoyo financiero.
BIBLIOGRAFÍA
ABBIATI, N; O ARREGOCÉ; D CARPIO; O LEMA; G ORTÍZ; MTG PUTRUELE ET AL. 1989. Avances en las investigaciones sobre moscas de las frutas en el litoral del río Uruguay. Boletín Técnico, Proyecto "Control integrado de moscas de la fruta", FAO. [ Links ]
ALUJA, M. 1999. Fruit fly (Diptera: Tephritidae) research in Latin America: myths, realities and dreams. An. Soc. Entomol. Brasil 28:565-594. [ Links ]
BATEMAN, MA. 1972. The ecology of fruit flies. Annu. Rev. Entomol. 17:493-518. [ Links ]
BEGON, M; JL HARPER & CR TOWNSEND. 1990. Ecology. Individuals, populations and communities. 2da edn. Blackwell. Cambridge. [ Links ]
BOSCÁN DE MARTÍNEZ, N. 1993. Niveles de infestación de moscas de la fruta de los géneros Anastrepha y Ceratitis en mango en Venezuela. Agron. Trop. 43:319-326. [ Links ]
DEBOUZIE, D & A MAZIH. 1999. Argan (Sapotaceae) trees as reservoirs for Mediterranean fruit fly (Diptera: Tephritidae) in Morocco. Environ. Entomol. 28:53-60. [ Links ]
FLETCHER, BS. 1986. Some issues in fruit fly ecology. Pp. 117-133 en: M Mangel; JR Carey & RE Plant (eds). Pest control: operations and systems analysis in fruit flies management. Springer-Verlag. Berlin y Heidelberg. [ Links ]
HARRIS, EJ & CYL LEE. 1986. Seasonal and annual occurrence of Mediterranean fruit flies (Diptera: Tephritidae) in Makaha and Waianae Valleys, Oahu, Hawaii. Environ. Entomol. 15:507-512. [ Links ]
HARRIS, EJ & CYL LEE. 1987. Seasonal and annual distribution of the Mediterranean fruit fly (Diptera: Tephritidae) in Honolulu and suburban areas of Oahu, Hawaii. Environ. Entomol. 16:1273-1282. [ Links ]
HARRIS, EJ; RI VARGAS & JE GILMORE. 1993. Seasonality in occurrence and distribution of Mediterranean fruit fly (Diptera: Tephritidae) in Upland and Lowland areas on Kauai, Hawaii. Environ. Entomol. 22:404-410. [ Links ]
KATSOYANNOS, BI; NA KOULOUSSIS & JR CAREY. 1998. Seasonal and annual occurrence of Mediterranean fruit flies (Diptera: Tephritidae) on Chios Island, Greece: differences between two neighboring citrus orchards. Ann. Entomol. Soc. Am. 91:43-51. [ Links ]
LACK, D. 1954. The natural regulation of animal numbers. Clarendon. Oxford. [ Links ]
LIQUIDO, NJ; LA SHINODA & RT CUNNINGHAM. 1990. Host plants of the Mediterranean fruit fly (Diptera: Tephritidae): an annotated world review. Misc. Publ. Entomol. Soc. Am. 77:1-52. [ Links ]
MALAVASI, A & JS MORGANTE. 1980. Biologia de "Moscas-das-frutas" (Diptera, Tephritidae). II: Indices de infestação em diferentes hospedeiros e localidades. Rev. Bras. Biol. 40:17-24. [ Links ]
MALAVASI, A & JS MORGANTE. 1981. Adult and larval population fluctuation of Anastrepha fraterculus and its relationship to host availability. Environ. Entomol. 10:275-278. [ Links ]
MALAVASI, A; JS MORGANTE & RA ZUCCHI. 1980. Biologia de "Moscas-das-frutas" (Diptera, Tephritidae). I: Lista de hospedeiros e ocorrência. Rev. Bras. Biol. 40:9-16. [ Links ]
MALAVASI, A; GG ROHWER & DS CAMPBELL. 1994. Fruit Fly Free Areas: strategies to develop them. Pp.165-180 en: CO Calkin (ed.). Fruit flies and the sterile insect technique. CRC Press. Boca Raton. [ Links ]
MAVRIKAKIS, PG; AP ECONOMOPOULOS & JR CAREY. 2000. Continuous winter reproduction and growth of the Mediterranean fruit fly (Diptera: Tephritidae) in Heraklion, Crete, Southern Greece. Environ. Entomol. 29:1180-1187. [ Links ]
MAZIH, A & D DEBOUZIE. 1996. Infestation rate of argan fruit (Argania spinosa) by the Mediterranean fruit fly (Ceratitis capitata) in relation to phenology and maturation of the fruit. Entomol. Exp. Appl. 81:31-38. [ Links ]
MEATS, A. 1989. Abiotic mortality factor- temperature. Pp. 229-239 en: AS Robinson & G Hooper (eds). Fruit flies: their biology, natural enemies and control. Elsevier. Amsterdam. [ Links ]
MESSENGER, PS & NE FLITTERS. 1954. Bioclimatic studies of three species of fruit flies in Hawaii. J. Econ. Entomol. 47:756-765. [ Links ]
PAPADOPOULOS, NT; BI KATSOYANNOS; JR CAREY & NA KOULOUSSIS. 2001. Seasonal and annual occurrence of the Mediterranean fruit fly (Diptera: Tephritidae) in Northern Greece. Ann. Entomol. Soc. Am. 94:41-50. [ Links ]
PUTRUELE, MTG. 1996. Hosts for Ceratitis capitata and Anastrepha fraterculus in the Northeastern Province of Entre Ríos, Argentina. Pp. 343-345 en: BA McPheron & GJ Steck (eds). Fruit fly pests: a world assessment of their biology and management. St. Lucie. Delray Beach. [ Links ]
PUTRUELE, MTG. 1998. Estudios bioecológicos de Ceratitis capitata Wiedemann (Diptera: Tephritidae): Influencia de la temperatura sobre el estado larvario, dinámica poblacional y validación de un modelo epidemiológico para predecir su fenología. Tesis Doctoral, Universidad Politécnica de Valencia, Valencia. [ Links ]
PUZZI, D & A ORLANDO. 1965. Estudios sobre a ecologia das "moscas-das-frutas" (Trypetidae) no Estado de São Paulo, visando o controle racional da praga. Arq. Inst. Biol. S. Paulo 32:9- 22. [ Links ]
TORROBA, CA; AT AMMA; A MITIDIERI & RE GAMIETEA. 1968. El cultivo del duraznero en el área de influencia de la Estación Experimental San Pedro. IDIA 220:1-43. [ Links ]
VARGAS, RI; EJ HARRIS & T NISHIDA. 1983. Distribution and seasonal occurrence of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) on the island of Kauai in the Hawaii Island. Environ. Entomol. 12:303-310. [ Links ]
VERA, MT; R RODRÍGUEZ; DF SEGURA; JL CLADERA & RW SUTHERST. 2002. Potential geographical distribution of the Mediterranean fruit fly,Ceratitis capitata (Diptera: Tephritidae), with emphasis on Argentina and Australia. Environ. Entomol. 31:1009-1022. [ Links ]
VERGANI, AR. 1961. La mosca del Mediterráneo. Boletín de divulgación, INTA. Concordia. [ Links ]

