










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.14 n.1 Córdoba ene./jun. 2004
TRABAJOS ORIGINALES
Asociación de las hormigas granívoras Pogonomyrmex pronotalis, P. rastratus y P. inermis con caminos en el Monte central
Gabriela I Pirk1,2,*, Javier López de Casenave1,2 & Rodrigo G Pol2
1. Grupo de Investigación en Ecología de Comunidades de Desierto (Ecodes), Depto. Ecología, Genética y Evolución, FCEyN, Universidad de Buenos Aires, Buenos Aires, Argentina
2. Grupo de Investigación en Ecología de Comunidades de Desierto (Ecodes), UFyEV, IADIZA, CONICET, Mendoza, Argentina
*Ecodes. Depto. Ecología, Genética y Evolución, FCEyN, Univ. de Buenos Aires. Piso 4, Pab. 2, C. Universitaria, C1428EHA Buenos Aires. Email: pirk@bg.fcen.uba.ar
Recibido: 10 de julio de 2003;
Fin de arbitraje: 8 de agosto de 2003;
Revisión recibida: 11 de septiembre de 2003;
Aceptado: 17 de octubre de 2003
RESUMEN. Los caminos y sus bordes presentan características diferentes al ambiente original que los rodea, que pueden influir sobre las poblaciones de insectos. Para evaluar el grado de asociación de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis con los caminos en el Monte central, medimos la densidad de colonias en el algarrobal y en dos caminos, y estudiamos la dieta y la actividad de estas especies en cada ambiente. La densidad de colonias de Pogonomyrmex pronotalis fue mayor en los caminos, la de Pogonomyrmex rastratus fue menor, y similar en ambos ambientes, y la de Pogonomyrmex inermis fue muy baja en los caminos y aún menor en el algarrobal. La actividad de Pogonomyrmex pronotalis fue mayor en los caminos y superior a las de Pogonomyrmex rastratus y Pogonomyrmex inermis, que presentaron similares niveles de actividad en los dos ambientes. Las dietas de las tres especies fueron muy parecidas en los dos ambientes, predominando las semillas (80-100% de los ítems acarreados), particularmente las de gramíneas (más del 75% de las semillas). La riqueza específica de las semillas en la dieta no difirió entre ambientes para ninguna especie. La proporción de algunas semillas en la dieta difirió entre ambientes para Pogonomyrmex pronotalis. Solo Pogonomyrmex pronotalis pareció estar asociada a los caminos presentando una mayor densidad de colonias, probablemente por la presencia de sitios favorables para el establecimiento y supervivencia de los nidos, y una mayor actividad que podría estar relacionada con temperaturas óptimas para esta especie. Los resultados sugieren que la ubicación del nido podría condicionar sólo muy sutilmente la ecología trófica de estas hormigas.
Palabras clave: Abundancia; Dieta; Actividad forrajera; Sitios de nidificación; Semillas.
ABSTRACT. Association between granivorous ants Pogonomyrmex pronotalis, P. rastratus and P. inermis, and roads in the central Monte desert: Roads and roadsides represent a modified habitat markedly different from the original environment in which they are embedded. Conditions associated with roads can influence some insects´ population levels. In order to evaluate the degree of association between Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis, and roads in the central Monte desert we estimated colony density in the open woodland and in two roads. We also studied the activity and the diet of the three species in colonies located in both habitats. Pogonomyrmex pronotalis´ colony density was higher in the roads, Pogonomyrmex rastratus´ was lower and similar in both habitats and Pogonomyrmex inermis´ was very low in the roads and even lower in the open woodland. Pogonomyrmex pronotalis´ activity was higher in roads and also higher than Pogonomyrmex rastratus´ and Pogonomyrmex inermis´. These two species presented similar levels of activity in both habitats. Diets were similar for the three species in the roads and in the open woodland. Seeds accounted for 80-100% of the items carried to the nests, and consisted mainly of grass seeds (more than 75% of the seeds). Seed species richness in the diets did not differ between habitats for any ant species. The proportion of some seeds in Pogonomyrmex pronotalis´ diet varied between habitats. Pogonomyrmex pronotalis was the only species that showed an association with roads. Its higher density there is probably due to favourable sites for nesting and establishment of colonies and its higher activity levels could be related to more suitable temperatures for this species. The results show that the location of the nest could only subtly influence the trophic ecology of these ants.
Keywords: Abundance; Diet; Foraging activity; Nesting sites; Seeds.
INTRODUCCIÓN
Los caminos y sus bordes constituyen un ambiente modificado que presenta diferencias marcadas con el ambiente que los rodea. Por ejemplo, suelen mostrar cambios en la textura del sustrato y un aumento de la escorrentía superficial debido a la mayor compactación de la superficie del suelo. También presentan usualmente una mayor temperatura a nivel del suelo, como consecuencia de un incremento en la insolación. Además, los caminos suelen estar dominados por plantas poco representadas en el ambiente original y poseer una riqueza específica relativamente alta, con predominio de especies tolerantes a las perturbaciones y, muy comúnmente, de especies exóticas (Forman & Alexander 1998, Gelbarb & Harrison 2003). A estas características se suman otras perturbaciones de origen humano, como emisiones de gases por vehículos, luces, movimiento y ruidos. Todos estos factores pueden influir sobre las poblaciones de algunos insectos, en muchos casos positivamente. En algunas especies de hormigas, por ejemplo, los caminos favorecen la instalación del nido y el desarrollo de la colonia (DeMers 1993; Samways et al. 1997; Terranella et al. 1999) y, como consecuencia, constituyen "parches" de hábitat favorable en paisajes que, en su ausencia, les serían inhabitables (DeMers 1993). Más aún, en algunos casos pueden servirles como corredores de dispersión, permitiéndoles la expansión de su distribución geográfica hacia ambientes normalmente menos favorables (DeMers 1993; Farji-Brener 1996) y convirtiéndose en rutas de invasión de especies invasoras (Stiles & Jones 1998).
La abundancia de colonias de hormigas en o cerca de los caminos está determinada por las respuestas de cada especie a una serie de características de estos ambientes. Éstas incluyen variables microambientales como luz, temperatura, humedad y temperatura del suelo (Johnson 1992, 1998, 2000, 2001; Stiles & Jones 1998), aspectos edáficos y de microtopografía (Johnson 1992, 1998, 2000, 2001; DeMers 1993; Crist & Wiens 1996; Dugas 2001) y otros relacionados con la vegetación (Rissing 1988; Blom et al. 1991), la disponibilidad de recursos (Gordon 1992; Farji-Brener 1996) y la presencia o ausencia de otras especies, tanto potenciales competidoras como predadoras (Hölldobler & Wilson 1990; Gotelli 1996; Gordon 1999).
Además de la abundancia de colonias, otros aspectos de la ecología de las hormigas, como la actividad forrajera y la composición de la dieta, podrían estar afectados por las diferencias ambientales entre los caminos y su entorno. La temperatura es uno de los principales factores asociados a los patrones temporales de actividad de forrajeo (Whitford & Ettershank 1975; Porter & Tschinkel 1987) y el comportamiento de forrajeo en hormigas granívoras es sensible a la disponibilidad de semillas y a su disposición espacial (Crist & MacMahon 1992; Wilby & Shachak 2000). Estos factores suelen ser marcadamente diferentes en los caminos con respecto al ambiente original y pueden así condicionar el forrajeo y la dieta de estas especies.
Las hormigas del género Pogonomyrmex, conocidas vulgarmente como hormigas cosechadoras ("harvester ants"), son especies mayormente granívoras muy comunes en las zonas áridas y semiáridas de América (Taber 1998; MacMahon et al. 2000). La respuesta de estas especies a las perturbaciones es variable. Hay especies para las cuales las perturbaciones del suelo (e.g., las asociadas con una elevada erosión superficial o con la presencia de ganado) tienen un impacto negativo sobre su abundancia y su actividad de forrajeo (Anderson & Mull 1992; Gordon 1992; Crist & Wiens 1996). Pero varias de las especies del género parecen no estar afectadas por otras perturbaciones como el fuego (McCoy & Kaiser 1990; Zimmer & Parmenter 1998). En el caso de los caminos, las respuestas solo han sido estudiadas en North Dakota (EEUU), donde se observó que el rango de distribución geográfica de Pogonomyrmex occidentalis aumentó, favorecido por la presencia de zanjas a los costados de los caminos (DeMers 1993).
En comparación con las especies que habitan América del Norte, las especies sudamericanas del género Pogonomyrmex han sido escasamente estudiadas (Taber 1998; MacMahon et al. 2000; Marone et al. 2000). Tres de estas especies coexisten en la porción central del desierto del Monte, en Argentina: Pogonomyrmex inermis, Pogonomyrmex pronotalis y Pogonomyrmex rastratus (Claver & Fowler 1993; Marone et al. 2000). Más allá de algunos aspectos básicos, se ignoran las características del hábitat asociadas a la ecología de estas especies. En particular, el objetivo de este trabajo es evaluar el grado de asociación de Pogonomyrmex inermis, Pogonomyrmex pronotalis y Pogonomyrmex rastratus con los caminos en el Monte central. Para ello, se estudiaron la densidad de colonias, la actividad forrajera y la dieta de estas especies en la Reserva de la Biosfera de Ñacuñán, analizando las diferencias entre las colonias ubicadas a lo largo de caminos y aquellas presentes en el bosque abierto de algarrobo (Prosopis flexuosa), el ambiente original y predominante en la reserva.
ÁREA DE ESTUDIO
El estudio se llevó a cabo en la Reserva de la Biosfera de Ñacuñán (34º03´S; 67º54´O; provincia de Mendoza, Argentina). La reserva se encuentra a una latitud intermedia del desierto del Monte (Marone et al. 2000).
El clima de Ñacuñán es árido-semiárido, marcadamente estacional, con veranos cálidos y relativamente húmedos e inviernos fríos y secos. La temperatura promedio anual es de 15.6°C. La máxima y la mínima promedio anuales son 23.8°C y 7.6°C, respectivamente. La precipitación anual es, en promedio, de 334 mm (1972-1998), pero es muy variable interanualmente. El 78% de las precipitaciones anuales se produce en los meses de primavera y verano (octubre a marzo), restringiéndose la producción de semillas de la mayoría de las plantas a ese período. La mayor parte de la reserva está ocupada por el bosque abierto de Prosopis flexuosa (algarrobal). Losárboles (Prosopis flexuosa y Geoffroea decorticans) se encuentran dispersos en una matriz compuesta por arbustos (fundamentalmente Larrea divaricata), arbustos bajos (Lycium spp., Verbena aspera y otros) y gramíneas (Trichloris crinita, Pappophorum spp., Sporobolus cryptandrus, Aristida spp., Digitaria californica, Setaria leucopila). La abundancia y la cobertura de las dicotiledóneas herbáceas (e.g., Chenopodium papulosum, Phacelia artemisioides, Parthenium hysterophorus) son muy variables entre años (Marone 1991).
La reserva se encuentra atravesada por numerosos caminos que tienen un trazado lineal y que, en su mayoría, confluyen radialmente en la Estación Biológica de la reserva (Figura 1). Exceptuando la ruta provincial 153, asfaltada, que cruza la reserva de norte a sur, y el camino principal que lleva desde el pueblo de Ñacuñán hasta la estación biológica, la mayor parte corresponde a viejas picadas de prospección petrolífera, de tierra y con un ancho de aproximadamente 6 m, incluyendo sus bordes. Son transitadas por personas, a pie, a caballo o con vehículos, pero con muy escasa frecuencia. Estos caminos presentan una cobertura de plantas mucho menor que el algarrobal adyacente (la cobertura leñosa se retira periódicamente para mantenerlos transitables), encontrándose la superficie del suelo más expuesta al sol, al viento y a la lluvia, y con un mayor grado de compactación. La composición y la abundancia relativa de especies de plantas asociadas también parecen diferir entre los caminos y el algarrobal.
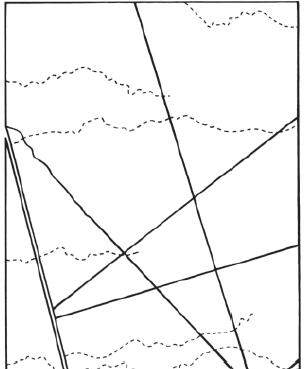

Figura 1. Mapa de la Reserva de Ñacuñán, Mendoza, Argentina. Las líneas negras corresponden a los principales caminos de la reserva, las líneas discontinuas a los cauces temporarios y la doble línea corresponde a la ruta 153 y a la vía del ferrocarril. Ñ: pueblo de Ñacuñán; EB: Estación Biológica.
Figure 1. Map of the Reserve of Ñacuñán, Mendoza, Argentina. Black lines represent the main roads, the broken lines are the temporary watercourses and the double line represents route 153 and the railway line. Ñ: town of Ñacuñán; EB: Biological Station.
MÉTODOS
Durante febrero de 2001 se estimó la densidad de colonias activas de Pogonomyrmex inermis, Pogonomyrmex pronotalis y Pogonomyrmex rastratus en el algarrobal y en caminos que lo atraviesan. En el algarrobal se contaron las colonias presentes en dos parcelas de 50×50 m. Ambas se encontraban a 50 m y 300 m de los dos caminos más cercanos y distaban entre sí unos 70 m. Para facilitar el conteo se subdividió cada parcela en cuadrados de 5×5 m, cada uno de los cuales fue relevado por la mañana y por la tarde, en los horarios de mayor actividad de estas hormigas (Pol 2001; Pol & Lopez de Casenave, en prensa). La densidad de colonias en los caminos se estimó contando las colonias presentes en dos transectas (una de 300×6 m y la otra de 345×6 m) ubicadas a lo largo de dos caminos dentro de la reserva. Estas también fueron relevadas en los horarios de mayor actividad de las hormigas.
Se analizó la dieta de Pogonomyrmex pronotalis y Pogonomyrmex rastratus (en febrero de 2001) y de Pogonomyrmex inermis (en febrero de 2002) en cinco colonias de cada especie en el algarrobal (cuatro en el caso de Pogonomyrmex rastratus) y en tres colonias de cada especie en uno de los caminos adyacentes. Se emplearon dos técnicas: (1) recolección manual de ítems acarreados, y (2) recolección con un dispositivo semiautomático de trampas de caída. La técnica de recolección manual consiste en la recolección de los ítems acarreados por las obreras que retornan al nido luego de su excursión de forrajeo. Se recolectaron ítems durante las horas de mayor actividad de la mañana y de la tarde, durante dos días. En cada una de las cuatro ocasiones de muestreo (i.e., dos horarios, dos días) se obtuvo la carga de, al menos, 10 obreras en cada colonia. La segunda técnica, basada en la propuesta por Crist & MacMahon (1991), consiste en capturar a las obreras que retornan acarreando ítems al nido mediante un dispositivo semiautomático. El dispositivo es una clausura circular de plástico de 33 cm de diá-metro y 10 cm de alto que se coloca rodeando la boca del nido. La clausura es transparente (por lo tanto, prácticamente no produce sombra sobre la boca del nido) y es suficientemente baja como para no obstaculizar el flujo de aire. Posee cuatro pares de aberturas (de 3×3 cm cada una y a 3 cm de altura), dispuestos a 90º. Cada par corresponde a una "entrada" y una "salida". Rampas de arena permiten a las obreras acceder a las aberturas del lado apropiado de la clausura. En el lado interno de cada "entrada" se ubica un recipiente cilíndrico enterrado (3 cm de diámetro, 5 cm de profundidad) que sobresale 3 cm de la superficie del suelo y que queda exactamente a la altura de la abertura de entrada. Cada recipiente tiene una tapa móvil que, al ser abierta, permite la captura por caída de las obreras que vuelven al nido. El dispositivo siempre fue colocado el día anterior al muestreo, para permitir que las hormigas se acostumbraran al uso de las "entradas y "salidas" y para que la instalación no perturbara la toma de las muestras. Muestreos piloto mostraron que las hormigas de las especies estudiadas son malas trepadoras (al igual que otras especies del mismo género; Gordon 1999) e incapaces de salir de los recipientes y de trepar por las paredes de la clausura. Los recipientes fueron abiertos durante una hora en los períodos de mayor actividad de la mañana y de la tarde, durante dos días. Al final de cada período de muestreo las hormigas de cada recipiente fueron devueltas a su nido.
Los elementos acarreados por las obreras fueron identificados y clasificados en seis categorías: semillas, flores, frutos, otras estructuras vegetales (restos de hojas, hojas enteras, restos de tallos), invertebrados (individuos enteros y porciones) y otros ítems (heces de vertebrados, musgos y líquenes, suelo). Las semillas fueron determinadas a nivel de especie o género usando colecciones de referencia y una guía de las semillas presentes en Ñacuñán (Peralta & Rossi 1997). Para estimar la dieta se calculó el porcentaje representado por cada tipo de ítem en cada colonia y luego se promediaron los porcentajes de las colonias de cada especie en cada ambiente.
La actividad forrajera de cada colonia se estimó con la tasa de captura registrada en las trampas de caída de los dispositivos semiautomáticos y fue definida como el número de obreras halladas en las trampas por unidad de tiempo. Como en el caso de la dieta, se promediaron los valores de las colonias de cada especie de hormiga en cada ambiente.
Se utilizó la Prueba de Mann-Whitney de dos colas para evaluar la diferencia entre ambientes en la riqueza específica de la dieta, el porcentaje que representan las semillas de cada especie en la dieta y la actividad de las colonias. Esta prueba no pudo ser usada con los datos de Pogonomyrmex rastratus por el bajo número de réplicas (colonias) empleado.
RESULTADOS
No se encontraron colonias de Pogonomyrmex inermis ni de Pogonomyrmex pronotalis en las parcelas ubicadas en el algarrobal (Figura 2). Sin embargo, sabemos que existen colonias en dicho ambiente (véase más abajo). Estos resultados sugieren que la densidad de colonias de estas especies en el algarrobal es muy baja, por debajo del nivel de detección de este método. En contraste, la densidad promedio para Pogonomyrmex rastratus fue de 16 colonias/ha en el algarrobal. La densidad de colonias de Pogonomyrmex pronotalis en los caminos fue mucho mayor que la de las otras dos especies en ese ambiente. La densidad de colonias de Pogonomyrmex rastratus fue similar en el algarrobal y en los caminos. Lo mismo sucedió con Pogonomyrmex inermis, aunque en este caso las densidades fueron mucho más bajas.

Figura 2. Densidad de colonias (promedio + EE) de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis en caminos (n = 2 caminos) y en el algarrobal (n = 2 parcelas) en la Reserva de Ñacuñán, Mendoza, Argentina.
Figure 2. Colony density (average + SE) of Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis in roads (n = 2 roads) and in the open woodland ("algarrobal", n = 2 areas) in the Reserve of Ñacuñán, Mendoza, Argentina.
Las dietas de Pogonomyrmex inermis, Pogonomyrmex pronotalis y Pogonomyrmex rastratus en las colonias del camino presentaron características generales muy similares a las del algarrobal (Tabla 1). Los resultados obtenidos con el dispositivo semiautomático mostraron que las semillas predominaron en todos los casos (90-95%), excepto para Pogonomyrmex inermis en los caminos (Tabla 1a). Sin embargo, en este caso el menor porcentaje de semillas se debió a que una colonia particular presentó una alta proporción de estructuras vegetales y otros ítems, a diferencia de las demás (lo cual quedó reflejado en el término de error; Tabla 1a). Los restos vegetales fueron siempre el segundo componente de la dieta, pero su proporción nunca superó el 10%. Con la técnica manual las semillas también preponderaron, en este caso con 96-100% (Tabla 1b). En cuanto a la riqueza específica de las semillas presentes en la dieta, no se detectaron diferencias significativas entre ambientes para ninguna de las tres especies de hormigas (Tabla 2).
Tabla 1. Dieta de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis en caminos y en el algarrobal en la Reserva de Ñacuñán, Mendoza, Argentina. Se muestran los promedios (± EE) de los porcentajes de cada tipo de ítem acarreado sobre el total de cada colonia, estimados usando un dispositivo semiautomático (a) y una técnica manual (b). También se indican el tamaño de muestra (número de colonias) y el número total de ítems recolectados.
Table 1. Diet of Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis in the roads and in the open woodland ("algarrobal") in the Reserve of Ñacuñán, Mendoza, Argentina. The average (± SE) of the percentages of each type of item over the total number of items carried to each nest, estimated by means of a semi-automated collecting device (a) and by hand collection (b) are shown. Sample size (number of colonies) and total number of items collected are also shown.

Tabla 2. Riqueza específica de semillas en la dieta de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis en caminos y en el algarrobal en la Reserva de Ñacuñán, Mendoza, Argentina, combinando los resultados obtenidos con el dispositivo semiautomático y la técnica manual. Se muestran los promedios (± EE) del número de especies acarreadas a cada nido y, entre paréntesis, el total de semillas. El tamaño de muestra es el mismo que en la Tabla 1a. ns: no significativo (P > 0.05); Prueba de Mann-Whitney.
Table 2. Species richness of seeds in the diet of Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis in the roads and in the open woodland ("algarrobal") of Ñacuñán, Mendoza, Argentina. Results are a combination of the semi-automated collecting device and hand collection samplings. The average (± SE) of the number of seed species carried to each nest is shown and the total number of seeds is between brackets. Sample size is the same as in Table 1a. ns: not significant (P > 0.05); Mann-Whitney Test.

Tanto en los caminos como en el algarrobal, las semillas de gramíneas constituyeron la mayor parte de la dieta granívora de las tres especies de hormigas (nunca menos del 75%; Figura 3). La relación entre gramíneas y no gramíneas no varió en la dieta de Pogonomyrmex inermis y Pogonomyrmex rastratus entre ambientes, pero sí en la de Pogonomyrmex pronotalis, con una menor proporción de gramíneas en los caminos (76%) que en el algarrobal (97%). Aristida spp., Trichloris crinita, Pappophorum spp. y Digitaria californica fueron las especies más acarreadas por Pogonomyrmex pronotalis y Pogonomyrmex rastratus en los dos ambientes y por Pogonomyrmex inermis en el algarrobal. En los caminos, Pogonomyrmex inermis acarreó mayoritariamente Digitaria californica, Aristida spp., Stipa ichu y Sporobolus cryptandrus.


Figura 3. Dieta granívora de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis en caminos (barras negras) y en el algarrobal (barras blancas) en la Reserva de Ñacuñán (Mendoza, Argentina) estimada usando un dispositivo semiautomático (a-c) y una técnica manual (d-f). Se muestran los promedios (+ EE) de los porcentajes de cada especie o grupo de especies de plantas sobre el total de semillas para cada ambiente. El tamaño de muestra es el mismo que en la Tabla 1. Ar: Aristida spp., Tr: Trichloris crinita, Pa: Pappophorum spp., Di: Digitaria californica, Ch: Chloris castilloana, Se: Setaria leucopila, Sp: Sporobolus cryptandrus, St: Stipa ichu, Ne: Neobouteloua barbata, Dp: Diplachne dubia, Dc: Dicotiledóneas. *: P < 0.05; ns: no significativo; Prueba de Mann-Whitney.
Figure 3. Granivorous diet of Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis in roads (black bars) and in the open woodland ("algarrobal", white bars) in the Reserve of Ñacuñán (Mendoza, Argentina), estimated by means of a semi-automated device (a-c) and by hand collection (d-f). Averages (+ SE) of the percentage of each species or group of species of seeds carried to each nest over the total number of seeds are shown for each type of habitat. Sample size is the same as in Table 1. See species codes in the legend above. *: P < 0.05; ns: not significant; Mann-Whitney Test.
Considerando en conjunto los resultados de las dos técnicas de estimación de la dieta utilizadas, el porcentaje que representaron las semillas de algunas especies en la dieta de Pogonomyrmex pronotalis fue significativamente diferente entre ambientes (Figura 3). Aristida spp. fue acarreada en mayor proporción en las colonias del algarrobal, mientras que Pappophorum spp., Sporobolus cryptandrus y las dicotiledóneas mostraron una mayor proporción en los caminos. Por su parte, aunque no pudo ser probado estadísticamente, Pogonomyrmex rastratus pareció acarrear un mayor porcentaje de Aristida spp. en el algarrobal y una proporción mayor de Pappophorum spp. y Digitaria californica en las colonias de los caminos. En la dieta de Pogonomyrmex inermis no se registraron diferencias significativas en la proporción de las especies entre ambientes (Figura 3).
La actividad forrajera de las colonias de las tres especies fue similar, con excepción de la de Pogonomyrmex pronotalis en los caminos, en donde fue tres veces mayor (Figura 4).

Figura 4. Actividad forrajera (promedio + EE) de colonias de Pogonomyrmex pronotalis, Pogonomyrmex rastratus y Pogonomyrmex inermis en caminos y en el algarrobal, en la Reserva de Ñacuñán, Mendoza, Argentina. El tamaño de muestra es el mismo que en la Tabla 1a. *: P < 0.05; ns: no significativo; Prueba de Mann-Whitney.
Figure 4. Foraging activity (average + EE) of Pogonomyrmex pronotalis, Pogonomyrmex rastratus and Pogonomyrmex inermis in colonies in roads and in the open woodland ("algarrobal") in Ñacuñán, Mendoza, Argentina. Sample size is the same as in Table 1a. *: P < 0.05; ns: not significant; Mann- Whitney Test.
DISCUSIÓN
Las hormigas del género Pogonomyrmex estudiadas presentaron distintos grados de asociación con los caminos. Pogonomyrmex pronotalis parece verse beneficiada por las condiciones de este ambiente particular; presentó allí una densidad de colonias muy alta y, sumado a esto, dichas colonias fueron mucho más activas que las del algarrobal. En general, una mayor actividad forrajera suele estar asociada con una mayor abundancia de obreras. Si así fuera, la combinación de mayores densidades de colonias y mayores niveles de actividad indicaría una diferencia marcada en la abundancia de estas hormigas en los caminos con respecto al algarrobal adyacente. De esta manera, este ambiente podría constituir un hábitat importante para la instalación o para la supervivencia de los nidos en esta especie y, eventualmente, podría estar asociado a expansiones de su distribución, como ha sido observado en Pogonomyrmex occidentalis en América del Norte (DeMers 1993). Sin embargo, esta hipótesis requiere de un análisis geográfico más minucioso. A diferencia de la especie anterior, la densidad de colonias de Pogonomyrmex rastratus y su nivel de actividad forrajera no fueron diferentes entre los caminos y el algarrobal. Finalmente, Pogonomyrmex inermis, al igual que Pogonomyrmex rastratus, no presentó variaciones entre ambientes, aunque en este caso su escasa abundancia impide una aseveración concluyente. La evaluación de las asociaciones con el hábitat de Pogonomyrmex inermis requiere de más estudios, ya que su abundancia es temporalmente muy variable en Ñacuñán: durante fines de la década del 80 su presencia era habitual (Claver & Fowler 1993), mientras que en los años previos a este estudio (1995-2000) era muy escasa (J Lopez de Casenave, obs. pers.) y en los años posteriores su abundancia aumentó otra vez, registrándose densidades de colonias mucho más altas (e.g., 50 colonias/ha en uno de los caminos en febrero de 2003; GI Pirk & J Lopez de Casenave, datos no publ.).
La diferencia en la densidad de colonias de Pogonomyrmex pronotalis entre caminos y algarrobal puede estar asociada a sus preferencias por nidificar en un determinado microhábitat. Los nidos de Pogonomyrmex pronotalis dentro del algarrobal están ubicados en sitios con alta proporción de suelo desnudo y escasa cantidad de mantillo y cobertura de plantas (Pol 2001; J Lopez de Casenave et al., datos no publ.), características que, precisamente, son típicas de los caminos en contraposición al algarrobal que los rodea. Esto podría indicar que en el algarrobal los sitios favorables para el establecimiento de sus colonias no son muy comunes, encontrando en los caminos las condiciones adecuadas y, por consiguiente, mostrando una asociación estrecha con ellos. Por el contrario, Pogonomyrmex rastratus, que no presenta diferencias entre ambientes, tiene un microhábitat de nidificación distinto al de Pogonomyrmex pronotalis en el algarrobal (Pol 2001), ubicando sus nidos tanto en zonas con mayor cobertura de plantas y de mantillo como en áreas de suelo desnudo (aunque generalmente debajo o cerca de la base de una gramínea). La mayor versatilidad de Pogonomyrmex rastratus en sus requerimientos de nidificación le permitiría establecerse en diferentes microambientes, traduciéndose esto en una ocupación relativamente homogénea del hábitat (i.e., tanto en el algarrobal como en los caminos y sus bordes).
No se conoce con certeza qué factor o factores inciden en la utilización de áreas abiertas para la ubicación de los nidos por parte de Pogonomyrmex pronotalis, pero es probable que tanto la humedad como la temperatura estén involucrados. Los requerimientos de humedad difieren entre especies de este género (Johnson 1992, 1998, 2001; DeMers 1993) y no parece improbable que Pogonomyrmex pronotalis necesite suelos más secos como aquellos que son característicos de las áreas de menor humedad dentro del algarrobal y, en especial, de los caminos. La temperatura, por su parte, afecta a las hormigas tanto directamente (e.g., en su tasa de consumo de oxígeno o de pérdida de agua) como indirectamente (e.g., en el costo del transporte de semillas), de manera tal que la actividad de las hormigas suele estar restringida a períodos en los cuales las condiciones térmicas les permiten mantener su temperatura corporal dentro de rangos fisiológicamente tolerables (Whitford & Ettershank 1975; Porter & Tschinkel 1987; Marsh 1988; Lighton & Feener 1989). Sugestivamente, los niveles máximos de actividad forrajera de Pogonomyrmex pronotalis ocurren a temperaturas del suelo mayores que los de Pogonomyrmex rastratus (35-45°C vs. 30-40°C, respectivamente; Pol & Lopez de Casenave, en prensa). Las áreas sin cobertura, como los caminos, alcanzan temperaturas superiores a aquellas cubiertas por vegetación (e.g., el suelo desnudo posee un promedio de hasta 21ºC más que bajo árboles y arbustos durante febrero en Ñacuñán; J Lopez de Casenave & FA Milesi, datos no publ.), y esto podría favorecer la actividad de Pogonomyrmex pronotalis y estar en relación directa con su preferencia por este tipo de ambiente. Cabe destacar que la diferencia en las densidades observadas entre ambientes también podría deberse a otros factores, como por ejemplo a diferencias en la tasa de supervivencia de las colonias entre los distintos ambientes, aunque este aspecto aún no se ha estudiado en estas especies.
Las diferencias en la dieta de las especies estudiadas entre el algarrobal y los caminos indican que la ubicación del nido podría condicionar sólo muy sutilmente la ecología trófica de estas hormigas. Las características generales de la dieta no variaron entre ambientes, pues las semillas (en especial las de gramíneas) constituyeron siempre el grueso del material acarreado, observándose diferencias solo en el porcentaje de algunas semillas en la dieta de Pogonomyrmex pronotalis. Si la disponibilidad de semillas fuera un factor importante en la determinación de la dieta y si las diferencias en la vegetación entre los caminos y el algarrobal tuvieran un correlato en la abundancia de semillas, se esperaría de estas hormigas un ajuste en su ecología trófica para enfrentarse a las condiciones en los caminos como el observado en Pogonomyrmex pronotalis. Sin embargo, el patrón general registrado en estas especies indica que las diferencias en la dieta entre ambientes son más bien la excepción. Esta situación puede deberse, entre otros factores, a que la distancia de forrajeo de estas hormigas (más de 30 m; RG Pol & J Lopez de Casenave, datos no publ.) hace improbable que se encuentren limitadas a la disponibilidad de semillas ofrecida por los caminos y sus bordes asociados, cuyo ancho promedio es de unos 6 m.
Si bien la remoción de semillas por hormigas granívoras no suele superar el 10% de la producción total local (Pulliam & Brand 1975; Whitford 1978), las hormigas pueden tener un impacto importante sobre las especies preferidas, removiendo en ocasiones hasta el 100% de las semillas preferidas disponibles (Crist & MacMahon 1992). Esta remoción selectiva puede causar cambios en la estructura de la comunidad de plantas al alterar la abundancia relativa de las especies o la distribución espacial de las plantas. En el caso de Pogonomyrmex pronotalis, este efecto podría estar potenciado por las diferencias en su abundancia y en los niveles de actividad entre los caminos y el algarrobal, de manera tal que puede esperarse un impacto significativamente mayor sobre las especies más removidas por esta especie, en o cerca de los caminos. La importancia de este efecto de los consumidores sobre el recurso (i.e., efecto "top-down"), y en particular su relación con el efecto del recurso sobre los consumidores (i.e., efecto "bottom-up"), solamente podrá ser dilucidada con estudios más detallados de las variaciones espacio-temporales de la dieta de estas hormigas y de la dinámica de las gramíneas.
AGRADECIMIENTOS
Agradecemos especialmente a F. A. Milesi, S. Claver y a cuatro revisores anónimos por la lectura crítica del manuscrito. L. Marone y V. R. Cueto leyeron una versión preliminar de este trabajo. F. A. Milesi, A. Ribichich y la familia Faión nos ofrecieron generosamente su ayuda en distintas ocasiones. El estudio fue financiado por la Agencia de Promoción Científica y Tecnológica (PICT 01-03187). Este trabajo es la contribución número 21 del Grupo de Investigación en Ecología de Comunidades de Desierto (Ecodes) de IADIZA y FCEyN (UBA).
BIBLIOGRAFÍA
ANDERSON, CJ & JF MULL. 1992. Emigration of a colony of Pogonomyrmex occidentalis Cresson (Hymenoptera: Formicidae) in Southwestern Wyoming. J. Kansas Entomol. Soc. 65:456-458. [ Links ]
BLOM, PE; WH CLARK & JB JOHNSON. 1991. Colony densities of the seed harvesting ant Pogonomyrmex salinus (Hymenoptera: Formicidae) in seven plant communities in Idaho National Engineering Laboratory. Journal of the Idaho Academy of Sciences 27:28-36. [ Links ]
CLAVER, S & HG FOWLER. 1993. The ant fauna (Hymenoptera, Formicidae) of the Ñacuñán Biosphere Reserve. Naturalia 18:189-193. [ Links ]
CRIST, TO & JA MACMAHON. 1991. Foraging patterns of Pogonomyrmex occidentalis (Hymenoptera: Formicidae) in a shrub-steppe ecosystem: the roles of temperature, trunk trails, and seed resources. Environ. Entomol. 20:265-275. [ Links ]
CRIST, TO & JA MACMAHON. 1992. Harvester ant foraging and shrub-steppe seeds: interactions of seed resources and seed use. Ecology 73:1768-1779. [ Links ]
CRIST, TO & JA WIENS. 1996. The distribution of ant colonies in a semiarid landscape: Implications for community and ecosystem processes. Oikos 76:301-311. [ Links ]
DEMERS, MN. 1993. Roadside ditches as corridors for range expansion of the western harvester ant (Pogonomyrmex occidentalis Cresson). Landscape Ecol. 8:93-102. [ Links ]
DUGAS, DP. 2001. The influence of arroyo edges on Pogonomyrmex rugosus nest distribution in the Chihuahuan Desert, New Mexico. J. Arid Environ. 47:153-159. [ Links ]
FARJI-BRENER, AG. 1996. Posibles vías de expansión de la hormiga cortadora de hojas Acromyrmex lobicornis hacia la Patagonia. Ecología Austral 6:144-150. [ Links ]
FORMAN, RT & LE ALEXANDER. 1998. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 29:207-231. [ Links ]
GELBARD, JL & S HARRISON. 2003. Roadless habitats as refuges for native grasslands: interactions with soil, aspect and grazing. Ecol. Appl. 13:404-415. [ Links ]
GORDON, DM. 1992. Nest relocation in harvester ants. Ann. Entomol. Soc. Am. 85:44-47. [ Links ]
GORDON, DM. 1999. Ants at work: how an insect society is organized. W. W. Norton & Co. New York. [ Links ]
GOTELLI, NJ. 1996. Ant community structure: effects of the predatory ant lions. Ecology 77:630-638. [ Links ]
HÖLLDOBLER, B & EO WILSON. 1990. The ants. Harvard University Press. Cambridge. [ Links ]
JOHNSON, RA. 1992. Soil texture as an influence on the distribution of desert seed-harvester ants Pogonomyrmex rugosus and Messor pergandei. Oecologia 89:118-124. [ Links ]
JOHNSON, RA. 1998. Foundress survival and brood production in the desert seed-harvester ants Pogonomyrmex rugosus and P. barbatus (Hymenoptera, Formicidae). Insect. Soc. 45:255-266. [ Links ]
JOHNSON, RA. 2000. Habitat segregation based on soil texture and body size in the seed harvester ants Pogonomyrmex rugosus and P. barbatus. Ecol. Entomol. 25:403-412. [ Links ]
JOHNSON, RA. 2001. Biogeography and community structure of North American seed-harvester ants. Annu. Rev. Entomol. 46:1-29. [ Links ]
LIGHTON, JRB & DH FEENER, JR. 1989. Water loss rate and cuticular permeability in foragers of the desert ant Pogonomyrmex rugosus. Physiol. Zool. 62:1232-1256. [ Links ]
MACMAHON, JA; JF MULL & TO CRIST. 2000. Harvester ants (Pogonomyrmex spp.): their community and ecosystem influences. Annu. Rev. Ecol. Syst. 31:265-291. [ Links ]
MARONE, L. 1991. Habitat features affecting bird spatial distribution in the Monte desert, Argentina. Ecología Austral 1:77-86. [ Links ]
MARONE, L; J LOPEZ DE CASENAVE & VR CUETO. 2000. Granivory in southern South American deserts: conceptual issues and current evidence. BioScience 50:123-132. [ Links ]
MARSH, AC. 1988. Activity patterns of some Namibia desert ants. J. Arid Environ. 14:31-73. [ Links ]
MCCOY, ED & BW KAISER. 1990. Changes in foraging activity of the southern harvester ants Pogonomyrmex badius (Latreille) in response to fire. Am. Midl. Nat. 123:112-123. [ Links ]
PERALTA, IE & BE ROSSI. 1997. Guía para el reconocimiento de especies del banco de semillas de la Reserva de la Biósfera de Ñacuñán (Mendoza, Argentina). Boletín de Extensión Científica, IADIZA 3:1-24. [ Links ]
POL, RG. 2001. Patrones de actividad diaria y estacional de las hormigas granívoras Pogonomyrmex rastratus y Pogonomyrmex pronotalis en la porción central del desierto del Monte. Tesis de licenciatura, Universidad Nacional de San Luis. [ Links ]
POL, R & J LOPEZ DE CASENAVE. En prensa. Activity patterns of harvester ants Pogonomyrmex pronotalis and Pogonomyrmex rastratus in the central Monte desert, Argentina. J. Insect Behav. [ Links ]
PORTER, SD & WR TSCHINKEL. 1987. Foraging in Solenopsis invicta (Hymenoptera: Formicidae): effects of weather and season. Environ. Entomol. 16:802-808. [ Links ]
PULLIAM, HR & MR BRAND. 1975. The production and utilization of seeds in plains grasslands of southeastern Arizona. Ecology 56:1158-1166. [ Links ]
RISSING, SW. 1988. Seed-harvester association with shrubs: competition for water in the Mohave Desert? Ecology 69:809-813. [ Links ]
SAMWAYS, MJ; R OSBORN & F CARLIEL. 1997. Effect of highway on ant (Hymenoptera: Formicidae) species composition and abundance, with a recommendation for roadside verge width. Biodivers. Conserv. 6:903-913. [ Links ]
STILES, JH & RH JONES. 1998. Distribution of the red imported fire ant, Solenopsis invicta, in road and powerline habitats. Landscape Ecol. 13:335-346. [ Links ]
TABER, SW. 1998. The world of the harvester ants. Texas A & M University Press, College Station. [ Links ]
TERRANELLA, AC; L GANZ & JJ EBERSOLE. 1999. Western harvester ants prefer nest sites near roads and trails. Southwest. Nat. 44:382-384. [ Links ]
WHITFORD, WG. 1978. Foraging in seed harvester ants Pogonomyrmex spp. Ecology 59:185-189. [ Links ]
WHITFORD, WG & G ETTERSHANK. 1975. Factors affecting foraging activity in Chihuahuan desert harvester ants. Environ. Entomol. 4:689-696. [ Links ]
WILBY, A & M SHACHAK. 2000. Harvester ant response to spatial and temporal heterogeneity in seed availability: pattern in the process of granivory. Oecologia 125:495-503. [ Links ]
ZIMMER, K & RR PARMENTER. 1998. Harvester ants and fire in a desert grassland: ecological responses of Pogonomyrmex rugosus (Hymenoptera: Formicidae) to experimental wildfires in central New Mexico. Environ. Entomol. 27:282- 287. [ Links ]

