










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.14 no.1 Córdoba Jan./June 2004
TRABAJOS ORIGINALES
La respuesta funcional: una revisión y guía experimental
Valeria Fernández-Arhex* & Juan C Corley
Laboratorio de Ecología de Insectos Forestales, INTA EEA - Bariloche, S. C. de Bariloche, Río Negro, Argentina
*Laboratorio de Ecología de Insectos Forestales, INTA EEA - Bariloche, S. C. de Bariloche, CC 277 (8400) Río Negro, Argentina. Email: varhex@bariloche.inta.gov.ar
Recibido: 11 de junio de 2003;
Fin de arbitraje: 22 de agosto de 2003;
Revisión recibida: 28 de agosto de 2003;
Aceptado: 2 de septiembre de 2003
RESUMEN. La respuesta funcional de los depredadores expresa la influencia del comportamiento de los enemigos naturales como individuos sobre la dinámica poblacional. Sin embargo, existen numerosas limitaciones inherentes al proceso de estimación que afectan la confianza en los resultados y, en consecuencia, en las conclusiones biológicas. Estas limitaciones incluyen el diseño experimental, el método estadístico y el modelo de respuesta funcional usados para analizar los datos y estimar sus parámetros. Este trabajo proporciona una revisión del concepto de respuesta funcional y una guía experimental para su determinación. A lo largo del texto enfocamos en los parasitoides por ser éste el grupo clásico de los estudios de respuesta funcional y por su importancia para el control biológico de plagas. Describimos la respuesta funcional, analizamos sintéticamente los problemas descriptos para su estimación y resumimos y ejemplificamos el método de mayor consenso para su estimación. De nuestra revisión surgen las ventajas de usar experimentos de tiempo variable, pues permiten al individuo bajo estudio actuar de un modo natural, como así también las de usar densidades de presas que se asemejen a las observadas en la naturaleza. A su vez, los estudios deben estar complementados con detalladas observaciones del comportamiento del depredador. En el momento de determinar la forma de la respuesta funcional es conveniente utilizar la regresión logística, pues reduce el problema de diferenciar entre una curva hiperbólica (tipo II) de una sigmoidea (tipo III). Luego, para estimar los parámetros de la curva, la regresión no lineal ajustada por cuadrados mínimos es aplicable. Finalmente, ejemplificamos este protocolo modelo con el estudio de la respuesta funcional del parasitoide Ibalia leucospoides.
Palabras clave: Respuesta comportamental; Regresión logística; Denso-dependencia; Insectos.
ABSTRACT. Functional response: an overview and experimental guide: The functional response of predators is central to any description of the process of predation, and is probably the best example of the influence of the individual behaviour on the population dynamics of predator-prey interactions. The estimation of the functional response is useful because of its implications on stability, but probably more importantly because it allows appropriate estimates for predation parameters and the comparison between potential agents of biological control. There are, however, several limitations in its estimation that affect the quality of the results obtained from empirical studies and, consequently, set a limit to the biological conclusions derived from them. These involve all stages of the estimation process: the design and implementation of the experiments, the analysis of data and the model used to estimate functional response parameters. Our aim in this paper is to establish a base line for the empirical study of the functional response. Throughout the paper, we set our focus on parasitoids because these insects are a classical group in studies of functional response, and also because of their importance as agents of biological control of pests. We first describe the functional response and present an overview of the main problems arising in its estimation. Then, we summarise and exemplify the procedure that allows the best estimation of the functional response. Our report differs from past attempts in that it involves all stages of the estimation process. The best protocol involves variable time experiments, because they allow the studied animals to display behaviours under a more natural scenario. Along these lines, prey patch densities should mimic natural densities and individual behaviour should be observed throughout the assay. Also, other individual traits such as the abilities to switch prey and learn must be standardised. Functional response is, after all, a behavioural study and thus careful observations allow detecting inconsistencies and flaws of the experiment and data collection. With the data, a Logistic Regression Analysis is appropriate, because it allows distinguishing readily between type II and type III functional responses. Once the shape of the response is determined, appropriate model parameters may be estimated by a non-linear least squares procedure. We finally illustrate this protocol, providing analysis details in an appendix, with a study of the functional response of the parasitoid Ibalia leucospoides.
Keywords: Behavioural response; Logistic regression; Density-dependence; Insects.
INTRODUCCIÓN
La influencia del comportamiento de los parasitoides sobre la dinámica de sus poblaciones y de la de sus presas posee su mejor ejemplo en la respuesta funcional. El término, usado originalmente por Solomon (1949), describe la relación entre el número de presas consumidas por un depredador en función de la densidad de la presa, en un espacio e intervalo de tiempo fijos. La respuesta funcional es central para cualquier descripción sobre parasitismo o depredación, precisamente porque el número de presas consumidas determina el desarrollo, supervivencia y reproducción de los depredadores o parasitoides (Oaten & Murdoch 1975). Sirve también para evaluar la potencialidad de agentes de control biológico de plagas (pero ver Fernández-Arhex & Corley 2003). Por ejemplo, permite conocer la densidad asintótica (de saturación) de presas, mas allá de la cual se incrementa la probabilidad de escape (Begon et al. 1995). También permite entender relaciones coevolutivas depredadorpresa e inferir acerca de los mecanismos etológicos básicos implícitos en dichas interacciones (Houck & Strauss 1985).
En su trabajo clásico, Holling (1959) describió tres tipos de respuesta funcional. En la respuesta de tipo I (Figura 1a) existe un aumento lineal de la tasa de ataque del depredador respecto a la densidad de la presa (con una pendiente igual a la eficiencia de búsqueda), hasta llegar a un punto a partir del cual la máxima tasa de ataque permanece constante. Este tipo de respuesta refleja uno de los aspectos más realistas de modelos clásicos como los de Thompson (1930) y Nicholson & Bailey (1935). En éstos, las interacciones huésped-parasitoide ocurren en ambientes estacionales y con una tasa de depredación denso-independiente (Hassell 2000).
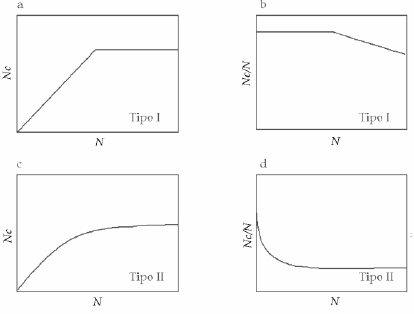

Figura 1. Los tres tipos de respuesta funcional más comunes. En a, c y e se muestra, para cada respuesta, la relación entre el número de presas consumidas por depredador (NC) y el número de presas ofrecidas al depredador (N), mientras que en b, d y f, se muestra la relación entre la proporción de presas consumidas (NC/N) y el número de presas ofrecidas al depredador (adaptado de Juliano 1993).
Figure 1. The three most common types of functional responses. In a, c, and e, the relationship between the number of prey consumed per predator (NC) and the number of prey offered (N) for each response are shown, whereas in b, c, and f, the relationship between the proportion of consumed prey (NC/N) and the number of prey offered is displayed (adapted from Juliano 1993).
En la respuesta funcional de tipo II (Figura 1c) aparece otro parámetro, además de la eficiencia de búsqueda, denominado "tiempo de manipulación" (Tm). Éste se define como el tiempo dedicado a perseguir, dominar, consumir y digerir las presas, y a prepararse para la siguiente búsqueda. Dado que el tiempo de búsqueda se reduce (debido al tiempo de manipulación), este tipo de respuesta resulta en un aumento desacelerado a medida que aumentan las presas consumidas, hasta llegar a una asíntota en la cual se expresa la máxima tasa de ataque (T/Tm). A esta densidad, el tiempo disponible por el depredador es usado para manipular la presa y el tiempo de búsqueda resulta entonces despreciable. En consecuencia, tiempos de manipulación largos conducen a bajas tasa de ataque y viceversa (Hassell 2000). Este tipo de relación resulta en un parasitismo densodependiente inverso, lo que implica que las presas, a altas densidades poblacionales, tendrán una menor probabilidad de ser atacadas que cuando están a densidades bajas. Teóricamente, la inclusión de este tipo de respuesta funcional en los modelos tipo Nicholson-Bailey desestabiliza al sistema (Hassell & May 1973; Walde & Murdoch 1988; Getz & Mills 1997). No obstante, como el tiempo de manipulación suele ser una fracción pequeña dentro del tiempo disponible, es probable que este efecto desestabilizador sea insignificante (Gullan & Cranston 2000). Para los parasitoides, tanto el tiempo de manipulación constante como la limitación por escasez de huevos conducen a respuestas de tipo II (Ivlev 1961).
Una curva sigmoidea es la que resulta en una respuesta de tipo III. En este caso, el número de presas consumidas por unidad de tiempo se acelera con el aumento de la densidad de presas, hasta que el tiempo de manipulación manipulación comienza a limitar su consumo (Figura 1e). Este tipo de respuesta produce, a bajas densidades de presa, una mortalidad denso-dependiente directa. Es por esta razón que solo la respuesta de tipo III es potencialmente estabilizadora de la dinámica presa-depredador y es importante destacar que esto sucede a bajas densidades de la presa (Hassell et al. 1977; Hassell 1978; Collins et al. 1981; Walde & Murdoch 1988; Chesson & Rosenzweig 1991; Berryman 1999; Bernstein 2000; Gullan & Cranston 2000). Una vez superado el punto de inflexión, la curva se parece a la descripta para la respuesta de tipo II.
Otros tipos de respuestas funcionales han sido descriptos en la literatura, como por ejemplo las respuestas con dos asíntotas, sin asíntota y en forma de cúpula (Casas & Hulliger 1994). Sin embargo, son muy escasos los ejemplos empíricos de estos modelos teóricos (Fernández-Arhex & Corley 2003).
Respuestas de tipo I se observan en general en animales filtradores; las de tipo II están generalmente ejemplificadas por depredadores invertebrados, incluidos los parasitoides, mientras que las de tipo III son más características de depredadores vertebrados, ya que este tipo de curvas surge típicamente por aprendizaje o por la posibilidad de cambiar una presa por otra ("switching"; Holling 1965; Berryman 1999; Gullan & Cranston 2000). Sin embargo, ciertos artrópodos depredadores y parasitoides poseen respuestas de tipo III (Hassell et al. 1977; pero ver Fernández-Arhex & Corley 2003).
En la estimación de la respuesta funcional, el mayor interés radica en conocer la forma general de la curva para el depredador en estudio, precisamente por sus potenciales efectos sobre la estabilidad del sistema (van Lenteren & Bakker 1976; Hassell et al. 1977). Otro aspecto importante es la estimación de sus parámetros. Determinar si los parámetros estimados de dos (o más) modelos son significativos es útil para comparar diferentes especies o edades de depredadores, por ejemplo, para determinar cuál de ellos es más efectivo en atacar una presa en particular (Thompson 1975) y para evaluar diferencias entre variaciones ambientales (O´Neil 1989). En términos evolutivos, la respuesta funcional puede ser útil para comparar diferentes especies o poblaciones de presas y encontrar evidencias de coevolución diferencial entre presa y depredador (Livdhal 1979; Houck & Strauss 1985; Juliano & Williams 1987). De este modo, tanto en términos básicos como aplicados, las conclusiones biológicas obtenidas del estudio de la respuesta funcional dependen fuertemente de su adecuada estimación.
LA DETERMINACIÓN DE LA RESPUESTA FUNCIONAL
Consideraciones generales
Tres aspectos son centrales para la estimación de la respuesta funcional: el diseño experimental, el análisis estadístico y el modelo a utilizar. Los tres han sido polémicos por diversas razones (van Lenteren & Bakker 1976; Livdhal & Stiven 1983; Houck & Strauss 1985; Juliano 1993; Manly & Jamienson 1999). En las secciones siguientes analizamos resumidamente estos tres aspectos, enfocando en los estudios hechos con parasitoides y haciendo referencia a aquellos puntos en discusión. En una sección final presentamos y luego ejemplificamos el método con mayor consenso para obtener la estimación más adecuada de la respuesta funcional.
El diseño experimental.- El diseño del experimento es un cuello de botella en la determinación de la respuesta funcional. Dos aspectos son relevantes: determinar las densidades de presas a usar y establecer la duración del experimento (Walde & Murdoch 1988; Bernstein 2000). Uno de los sesgos de algunos experimentos se refiere a las limitaciones en la oferta de bajas densidades de presa. En este caso, respuestas sigmoideas (Tipo III) pueden verse oscurecidas, y se hace difícil poder distinguir entre una curva hiperbólica de una sigmoidal. Es precisamente a bajas densidades donde las respuestas de tipo II y de tipo III difieren, y esta diferencia tiene importantes consecuencias sobre la interacción. Para muchos vertebrados depredadores que poseen la habilidad de concentrarse en un tipo particular de presa cuando la densidad de esa presa va aumentando, especialmente cuando dicha presa es preferida sobre otras, el problema de distinguir entre respuestas de tipo II y III se ha resuelto mediante el agregado de un excedente de una presa alternativa, ofrecida junto con la presa predilecta en todas las densidades. De este modo, el depredador va a tener la opción de alternar de presa en el momento en que la presa preferida deje de aumentar y no utilizará todo su tiempo en su búsqueda (van Lenteren & Bakker 1976). No obstante, para parasitoides, en su mayoría especialistas, el ofrecimiento de bajas densidades de huésped se ve muchas veces afectado debido a la necesidad del investigador de inducir una respuesta positiva frente al estímulo.
También existe debate sobre la duración del experimento: si se deben realizar en un tiempo fijo (preestablecido por el experimentador) o en un tiempo variable (no preestablecido). Los primeros han sido criticados debido a que esto puede no reflejar el verdadero comportamiento del parasitoide en su ambiente natural. Experimentos a tiempo fijo producen generalmente respuestas de tipo II (Griffiths 1969; Collins et al. 1981; Hertlein & Thorarinsson 1987). Si bien en este tipo de experimentos es raro que se observe el comportamiento del parasitoide, una curva hiperbólica se atribuye a un aumento en el tiempo de manipulación o a una limitación en el número de huevos. En cambio, los mecanismos que producen una respuesta de tipo III a tiempo fijo se asocian con un decrecimiento en el tiempo de manipulación o a un incremento en el tiempo de búsqueda, en respuesta a un aumento en la densidad del huésped. La importancia relativa de ambos aspectos no está clara. Venturia canescens (Gravenhorst, 1829) (Hymenoptera: Ichneumonidae) es un ejemplo en el cual el tiempo de manipulación decrece a medida que aumenta la densidad de huéspedes a la vez que su tiempo de búsqueda aumenta (Cook & Hubbard 1977).
Los experimentos a tiempo fijo indican también que la distribución espacial de los huéspedes puede afectar la tasa de parasitismo (Madden & Pimentel 1965; Cheke 1974). Por ejemplo, el parasitismo por Encarsia formosa (Gahan, 1924) (Hymenoptera: Aphelinidae) es más alto cuando los huéspedes están agregados que cuando están distribuidos al azar (Burnett 1958a). A su vez, se observó que un aumento en el tamaño del parche (e.g., la distancia entre los huéspedes) puede cambiar el patrón de parasitismo desde denso-independiente (respuesta de tipo I) a denso-dependiente directo (respuesta de tipo III) (Burnett 1958b). La distribución espacial de los huéspedes puede inducir la migración de los parasitoides en busca de mejores parches de huéspedes (van Lenteren & Bakker 1978). En experimentos a tiempo variable es posible contrarrestar este problema al evaluar el comportamiento de agregación del parasitoide, es decir, la tendencia del mismo a permanecer más tiempo en áreas con mayor disponibilidad de huéspedes (Walde & Murdoch 1988).
Los experimentos a tiempo variable tienden a producir respuestas tanto de tipo I como de tipo III (Oaten 1977; Morrison 1986; Hertlein & Thorarinsson 1987). Esto es porque, por ejemplo, un parasitoide que busca durante un tiempo constante en cada parche, va a residir más tiempo en aquel parche cuya densidad de huéspedes sea mayor y, a su vez, pasará más tiempo manipulando a los huéspedes. De este modo, el tiempo total de residencia en cada parche aumenta linealmente con el número de huéspedes presentes, caracterizando una respuesta de tipo I. Hertlein & Thorarinsson (1987) demostraron que, incluso aunque el tiempo total aumente con la densidad del parche, el tiempo de búsqueda no varía y la curva resultante es una de tipo I. Una alternativa es que el parasitoide sólo aumente su búsqueda en áreas donde la recompensa es mayor (Hassell & May 1974; Murdoch & Oaten 1975). En este caso, tanto el tiempo de búsqueda como el tiempo total se incrementan con el número de presas en cada parche. Entonces el parasitismo puede ser denso-dependiente y el tiempo total en el parche se acelerará con el número de huéspedes. Un ejemplo de esto lo constituye la avispa Trichogramma spp. (Hymenoptera: Trichogrammatidae), quien presenta una respuesta de tipo III precisamente porque busca más en parches donde hay mayor densidad de huéspedes (Morrison 1986). Van Lenteren & Bakker (1976) demostraron que si aumenta el tiempo para abandonar el parche ("giving up time") luego de que un parasitoide encuentra a su huésped, su respuesta funcional también será de tipo III. En estudios comparativos en los cuales se incluyeron experimentos a tiempo fijo y variable, la curva de tipo II que resultó a tiempo fijo cambió a tipo III o I cuando se realizó el mismo experimento pero con tiempo variable (van Lenteren & Bakker 1978; Hertlein & Thorarinsson 1987).
El método estadístico.- Para poder distinguir, a partir de los datos experimentales, entre una respuesta de tipo II (hiperbólica) y una de tipo III (sigmoidea), se han utilizado diferentes métodos estadísticos. La regresión lineal por cuadrados mínimos es el método más común, en el cual se involucra al número de presas consumidas o huéspedes atacados (NC) y a la densidad de la presa o densidad del huésped (N) (Livdhal & Stiven 1983; Williams & Juliano 1985; Trexler et al. 1988). También han sido usadas las regresiones no lineales por cuadrados mínimos para la estimación y comparación de parámetros (Juliano & Williams 1987). Trexler et al. (1988) demostraron, en un trabajo muy importante, que la mejor manera de analizar la forma de la respuesta funcional y para distinguir entre una curva de tipo II o de tipo III es mediante la regresión logística, ajustando una ecuación polinomial de la proporción de presas consumidas (NC/N) vs. el número de presas ofrecidas (N). La regresión logística consiste en ajustar una función que predice cómo responde la proporción de huéspedes atacados para una (o más) variables continuas. En este caso, la variable continua es N y la variable dependiente (Y) es la probabilidad de que el huésped expuesto sea atacado (de tipo binario). Si la proporción de presas consumidas inicialmente se incrementa con el número de presas ofrecidas, se identifica una respuesta de tipo III (Figura 1f). Si, por el contrario, la proporción de presas consumidas decrece monotónicamente, se identifica una respuesta de tipo II (Figura 1d; ver Juliano 1993 para más detalles). Una vez identificada la forma general de la curva, se utiliza el modelo correspondiente a dicha respuesta (ver a continuación) para estimar los parámetros de la respuesta funcional utilizando, como método estadístico, una regresión no lineal ajustada por cuadrados mínimos del número de presas consumidas vs. el número de presas ofrecidas (Juliano 1993).
El modelo de la respuesta funcional.- Numerosos modelos mecanísticos y fenomenológicos han sido utilizados para describir las respuestas funcionales (Juliano 1993). Aquí resumimos los principales, siendo el centro del debate en esta sección las respuestas de tipo II y III.
El modelo original fue descripto por Holling (1966), quien representó la respuesta funcional de tipo II utilizando la conocida "ecuación de los discos" :
| NC = (aNT) / (1 + aNTm) | (1) |
Una respuesta funcional de tipo III, que no permite el agotamiento ("depletion") de las presas, implica, por el contrario, que la tasa de ataque (a) varía en función de la densidad de la presa (N) (Hassell 1978):
| a = (d + bN) / (1 + cN) | (2) |
donde b, c y d son constantes. Sustituyendo la ecuación (2) en la (1), se obtiene la relación sigmoidea que postuló Hassell (1978) para la respuesta de tipo III:
| NC=(dNT + bN2T) / (1+cN+dNTm+bN2Tm) | (3) |
Royama (1971) y Rogers (1972) propusieron un modelo alternativo, conocido como "ecuación del depredador al azar", que tiene las mismas restricciones que el modelo mencionado anteriormente, pero que permite el agotamiento de la presa. La ecuación (4) integra la tasa instantánea de depredación sobre la duración total del experimento (T), suponiendo un decrecimiento exponencial en la densidad de la presa. Resulta apropiado en experimentos en los cuales no se observa continuamente a los predadores (Houck & Strauss 1985; Juliano 1993):
| NC = N {1 - exp [a (TmNC - T)]} | (4) |
Sustituyendo la ecuación (2) en la (4), la respuesta de tipo III es la siguiente (Hassell et al. 1977; Hassell 1978):
| NC=N{1-exp[(d+bN)(T NC-T)/(1+ cN)]} | (5) |
Guía práctica para estimar la respuesta funcional
En esta sección pretendemos ofrecer una guía para determinar el tipo de respuesta funcional, con el fin de estandarizar la forma de estudio y su futura comparación. La misma ha sido descompuesta en sus tres aspectos claves: el diseño experimental, el análisis estadístico y la estimación de los parámetros.
Diseño experimental.- Es aconsejable utilizar diseños de tiempo variable. Éstos permiten interpretar el comportamiento del individuo bajo estudio de un modo más natural y aplicar reglas de uso de parches que efectivamente utiliza en condiciones de campo. Por su parte, se debe intentar usar densidades que reproduzcan las observadas en la naturaleza y evitar exagerar las densidades para inducir la respuesta. Otros dos aspectos importantes son considerar los efectos del agotamiento y, para depredadores generalistas y oportunistas, los efectos de las presas alternativas. Los estudios deben estar complementados con detalladas observaciones del comportamiento del depredador. La respuesta funcional estudia, en definitiva, un aspecto del comportamiento y, como tal, sigue reglas propias de ese nivel. La observación cautelosa permite comprender el proceso y detectar inconsistencias. Finalmente, tanto en el caso de parasitoides como en el de depredadores no debe olvidarse que el aprendizaje puede ser un fenómeno común y, consecuentemente, debe estandarizarse esta variable.
Análisis estadístico.- Este punto vale tanto para depredadores como para parasitoides. Para conocer la forma general de la respuesta funcional, es claramente aconsejable utilizar la regresión logística (Juliano 1993) de la proporción de presas consumidas vs. el número de presas ofrecidas. De esta manera, se evita el problema de discriminar, a bajas densidades de presa, entre una curva hiperbólica y una sigmoidea.
Estimación de los parámetros.- En este punto se aconseja utilizar la regresión no lineal, ajustada por cuadrados mínimos, del número de presas consumidas vs. el número de presas ofrecidas, utilizando la ecuación correspondiente para cada tipo de respuesta funcional según resulte la regresión logística.
Ejemplo de estimación de respuesta funcional
Ibalia leucospoides (Hochenwarth, 1785) (Hymenoptera: Ibaliidae) es un parasitoide de la avispa plaga Sirex noctilio (Fabricius, 1793) (Hymenoptera: Siricidae). El experimento consistió en colocar parasitoides "naive" (hembras de menos de 48 h de vida) en jaulas conteniendo una troza de pino (longitud: 0.5 m, diámetro: 9.5-11 cm) con agujeros de oviposición del huésped, a temperatura ambiente (26.2 ± 0.2 °C) y bajo luz natural. Los parasitoides que se utilizaron fueron previamente alimentados (con solución de sacarosa 30%) y dejados a fecundar durante 24-48 h. Las densidades del huésped que se usaron (i.e., los tratamientos) fueron 2, 4, 6, 10 y 14 agujeros de oviposición por troza. Cada tratamiento fue replicado 10 veces, cambiando tanto el parasitoide como la troza, para evitar pseudoreplicación. El ensayo terminó cuando el parasitoide abandonó la troza y no retornó pasados 5 min (tiempo variable).
Las preguntas que interesan para este ejemplo fueron: (1) ¿cuál es la forma de la respuesta funcional de Ibalia leucospoides? y (2) ¿cuáles son los parámetros que mejor describen la función de esa respuesta? Por lo tanto, las variables de respuesta estudiadas fueron: (1) el número de agujeros (i.e., huéspedes) visitados y atacados, y (2) el tiempo de permanencia sobre la troza.
El primer paso en el análisis de datos consistió en ajustar una regresión logística, para poder determinar la forma de la curva (Juliano 1993). La estructura de entrada de los datos y el modelo utilizado se muestran en el Apéndice. El procedimiento básico consiste en evaluar sucesivos modelos polinomiales (ver ecuación en Apéndice) y sus parámetros desde una ecuación cúbica a una lineal. En este ejemplo, tanto la ecuación cúbica como la cuadrática resultaron no significativas; por lo tanto, se ajustó una ecuación lineal (Tabla 1). El parámetro β0 no es particularmente informativo, pero el lineal ( β1), que resultó negativo y significativo, indica que la respuesta funcional de Ibalia leucospoides es de tipo II. Entonces, la proporción de agujeros atacados decrece monotónicamente a medida que aumenta la cantidad de agujeros ofrecidos.
Tabla 1. Resultado del Análisis de Regresión Logística de la proporción de agujeros de oviposición del huésped atacados por el parasitoide Ibalia leucospoides vs. el número inicial de agujeros de oviposición ofrecidos.
Table 1. Results of a Logistic Regression Analysis of the proportion of host oviposition drills attacked by the parasitoid Ibalia leucospoides vs. number of drills offered.

A continuación se estimaron los parámetros para la ecuación de tipo II, mediante una regresión no lineal ajustada por cuadrados mínimos. En este ejemplo, la función general que se utilizó fue la ecuación (4) que involucra la estimación de dos parámetros: a y Tm. Para la realización de la regresión no lineal se requieren por cada réplica solamente un par de observaciones que son los valores correspondientes a NC y N. Debido a que T es variable fue incluido como una función específica, basada en una distribución del tiempo total de permanencia en la troza por cada parasitoide estudiado (Fernández-Arhex et al., datos no publ.).
La regresión no lineal, como otros procedimientos no lineales, es iterativa y, por lo tanto, requiere de estimaciones iniciales de los parámetros y, en muchos casos, este es un elemento clave. En los modelos de respuesta funcional, el Tm puede ser aproximado por T NC a altas densidades de N; la estimación inicial de los otros parámetros resulta menos obvia y se realiza por aproximación. En el modelo de la respuesta de tipo III, además de Tm hay que estimar tres parámetros adicionales (b, c y d) usados para calcular a (ver ecuación 2). En este modelo (ver Juliano 1993), la estimación inicial del parámetro d parte de una estimación inicial del orden de 0, debido a que en la mayoría de los casos el valor de este parámetro es bajo. A su vez, para los parámetros b y c es mejor seleccionar varias estimaciones iniciales variando entre dos órdenes de magnitud. Hay que tener en cuenta que los resultados deben tener un sentido biológico. Por ejemplo, no pueden existir Tm negativos y este parámetro junto con b deben ser mayores a 0, así como positivos los parámetros c y d.
Los resultados de la regresión no lineal para el modelo de tipo II indicaron que los parámetros resultaron significativos con un intervalo de confianza de 95% que no incluye al 0, en donde la tasa de ataque promedio (± EE) resultó ser 0.0004 ± 0.0003 seg-1 con un tiempo de manipulación de 10 ± 3 min (Figura 2).

Figura 2. Respuesta funcional de tipo II del parasitoide Ibalia leucospoides.
Figure 2. Type II functional response of the parasitoid Ibalia leucospoides.
APÉNDICE
Estructura de entrada de datos para realizar un Análisis de Regresión Logística, usando el programa estadístico SAS (SAS Institute Inc 1999). En este caso se modeló una ecuación lineal.
data a;
input tronco agujtot agujatac;
proporc=agujatac/agujtot;
datalines;
1 2 1
2 2 2
3 2 2
...
38 14 7
39 14 5
40 14 4
proc plot;
plot proporc*agujatac;
run;
proc genmod;
model agujatac / agujtot = agujtot / dist=bin link=logit;
run;
tronco=el número del experimento
agujtot=el número de agujeros ofrecidos por tronco.
agujatac=el número de agujeros atacados visitados por el parasitoide.
proc plot=es la sentencia que se utiliza para realizar un gráfico
proc genmod= es la sentencia que se utiliza para realizar un modelo generalizado.
link=logit=es la sentencia para que se realice una regresión logística.
dist=bin= indica que se utiliza una distribución binomial.
Ecuación usada para la regresión logística: conviene comenzar el análisis utilizando una ecuación cúbica, ya que es un modelo complejo que sirve para describir la mayoría de los datos experimentales.
Log (π/1-π) = β0+ β1X+ β2X2+ β3X3
donde β0 es el parámetro constante o intercepto, β1 es el parámetro lineal, β2 es el parámetro cuadrático, β3 es el parámetro cúbico y X es la densidad de agujeros de oviposición ofrecidos por troza.
AGRADECIMIENTOS
Los autores agradecen a C. Gittins por el apoyo recibido para la realización de este trabajo. Cuatro revisores han hecho aportes muy valiosos. Este estudio es parte de un proyecto financiado por el Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) (PIP 529/98). El autor y el coautor son becario e investigador del CONICET, respectivamente.
BIBLIOGRAFÍA
BEGON, M; JL HARPER & CR TOWNSEND. 1995. Ecología: individuos, poblaciones y comunidades. Ed. Omega. Barcelona. [ Links ]
BERNSTEIN, C. 2000. Host-parasitoid models: the story of successful failure. Pp. 41-57 en: M Hochberg & A Ives (eds). Population biology of host-parasitoid interactions. Princeton University Press. Princeton. [ Links ]
BERRYMAN, AA. 1999. The theoretical foundations of biological control. Pp. 3-21 en: BA Hawkins & HV Cornell (eds). Theoretical approaches to biological control. Cambridge University Press. Cambridge. [ Links ]
BURNETT, T. 1958a. Effect of host distribution on the reproduction of Encarsia formosa Gahan (Hymenoptera: Chalcidoidea). Can. Entomol. 90:179-191. [ Links ]
BURNETT, T. 1958b. Effect of area of search on reproduction of Encarsia formosa Gahan (Hymenoptera: Chalcidoidea). Can. Entomol. 90:225-229. [ Links ]
CASAS, J & B HULLINGER. 1994. Statistical analysis of functional response experiments. Biocontrol Sci. Techn. 4:133-145. [ Links ]
CHEKE, RA. 1974. Experiments on the effect of host spatial distribution on the numerical response of parasitoids. J. Anim. Ecol. 43:107-114. [ Links ]
CHESSON, P & M ROSENZWEIG. 1991. Behaviour, heterogeneity, and the dynamics of interacting species. Ecology 72:1187-1195. [ Links ]
COLLINS, MD; SA WARD & AFG DIXON. 1981. Handling time and the functional response of Aphelinus thomsoni, a predator and parasite of the aphid Drepanosiphum platanoidis. J. Anim. Ecol. 50:479-487. [ Links ]
COOK, RM & SF HUBBARD. 1977. Adaptative searching strategies in insect parasites. J. Anim. Ecol. 46:115-125. [ Links ]
FERNÁNDEZ-ARHEX, V & JC CORLEY. 2003. The functional response of parasitoids and its implications for biological control. Biocontrol Sci. Techn. 13:403-413. [ Links ]
GETZ, WM & NJ MILLS. 1997. Host-parasitoid coexistence and egg-limited encounter rates. Am. Nat. 148:333-347. [ Links ]
GRIFFITHS, KJ. 1969. The importance of coincidence in the functional and numerical responses of two parasites of the European pine sawfly, Neodiprion sertifer. Can. Entomol. 101:673-713. [ Links ]
GULLAN, PJ & PS CRANSTON. 2000. The insects: an outline of Entomology. 2da edn. Blackwell Science. Carleton. [ Links ]
HASSELL, MP. 1978. The dynamics of arthropod predator-prey systems. Princeton University Press, Princeton. [ Links ]
HASSELL, MP. 2000. The spatial and temporal dynamics of host-parasitoid interactions. Oxford University Press. Oxford. [ Links ]
HASSELL, MP; JH LAWTON & JR BEDDINGTON. 1977. Sigmoid functional responses by invertebrate predators and parasitoids. J. Anim. Ecol. 46:249-262. [ Links ]
HASSELL, MP & RM MAY. 1973. Stability in host-parasite models. J. Anim. Ecol. 42:693-726. [ Links ]
HASSELL, MP & RM MAY. 1974. Aggregation in predators and insect parasites and its effect on stability. J. Anim. Ecol. 43:567-594. [ Links ]
HERTLEIN, MB & K THORARINSSON. 1987. Variable patch times and the functional response of Leptopilina boulardi (Hymenoptera: Eucoilidae). Environ. Entomol. 16:593-598. [ Links ]
HOLLING, CS. 1959. Some characteristics of simple types of predation and parasitism. Can. Entomol. 91:385-398. [ Links ]
HOLLING, CS. 1965. The functional response of predators to prey density and its role in mimicry and population regulation. Mem. Entomol. Soc. Can. 45:3-60. [ Links ]
HOLLING, CS. 1966. The functional response of invertebrate predators to prey density. Mem. Entomol. Soc. Can. 48:1-86. [ Links ]
HOUCK, MA & RE STRAUSS. 1985. The comparative study of functional responses: experimental design and statistical interpretation. Can. Entomol. 117:617-629. [ Links ]
IVLEV, VS. 1961. Experimental ecology of the feeding of fishes. Yale University Press. New Haven. [ Links ]
JULIANO, S.A. 1993. Nonlinear curve fitting: predation and functional response curves. Pp. 159-182 en: SM Scheiner & J Gurevitch (eds). Design and analysis of ecological experiments. Chapman & Hall, Nueva York. [ Links ]
JULIANO, SA & FM WILLIAMS. 1987. A comparison of methods for estimating the functional response parameters of the random predator equation. J. Anim. Ecol. 56:641-653. [ Links ]
van LENTEREN, JC & K BAKKER. 1976. Functional responses in invertebrates. Neth. J. Zool. 26:567-572. [ Links ]
van LENTEREN, JC & K BAKKER. 1978. Behavioural aspects of the functional responses of a parasite (Pseudeucoila bochei Weld) to its host (Drosophila melanogaster). Neth. J. Zool. 28:213-233. [ Links ]
LIVDHAL, TP. 1979. Evolution and handling time: the functional response of a predator to the density of sympatric and allopatric strains of prey. Evolution 33:765-768. [ Links ]
LIVDHAL, TP & AE STIVEN. 1983. Statistical difficulties in the analysis of predator functional response data. Can. Entomol. 115:1365-1370. [ Links ]
MADDEN, JL & D PIMENTEL. 1965. Density and spatial relationships between a wasp parasite and its housefly host. Can. Entomol. 97:1031-1037. [ Links ]
MANLY, BFJ & CD JAMIENSON. 1999. Functional response and parallel curve analysis. Oikos 85:523-528. [ Links ]
MORRISON, G. 1986. "Searching time aggregation" and density dependent parasitism in a laboratory host-parasitoid interaction. Oecologia 68:298-303. [ Links ]
MURDOCH, WW & A OATEN. 1975. Predation and population stability. Adv. Ecol. Res. 9:1-131. [ Links ]
NICHOLSON, AJ & VA BAILEY. 1935. The balance of animal populations. Part 1. P. Zool. Soc. Lond. 3:551-598. [ Links ]
OATEN, A. 1977. Optimal foraging in patches: a case for stochasticity. Theor. Popul. Biol. 12:263-285. [ Links ]
OATEN, A & WW MURDOCH. 1975. Functional response and stability in predator-prey systems. Am. Nat. 109:289-298. [ Links ]
O´NEIL, RJ. 1989.Comparison of laboratory and field measurements of the functional response of Podisus maculaventris (Heteroptera: Pentatomidae). J. Kansas Entomol. Soc. 62:148-155. [ Links ]
ROGERS, D. 1972. Random search and insect population models. J. Anim. Ecol. 41:369-383. [ Links ]
ROYAMA, T. 1971. A comparative study of models for predation and parasitism. Res. Popul. Ecol. 1:1-91. [ Links ]
SAS INSTITUTE INC. 1999. SAS® system for Microsoft® Windows®, v. 8.0 (TSMO). SAS Institue Inc. Cary. [ Links ]
SOLOMON, JE. 1949. The natural control of animal populations. J. Anim. Ecol. 18:1-35. [ Links ]
THOMPSON, DJ. 1975. Towards a predator-prey model incorporating age structure: effects of predator and prey size on the predation of Daphnia magna by Ischnura elegans. J. Anim. Ecol. 44:907-916. [ Links ]
THOMPSON, WR. 1930. The utility of mathematical methods in relation to work on biological control. Ann. Appl. Biol. 17:641-648. [ Links ]
TREXLER, JC; CE MCCULLOGH & J TRAVIS. 1988. How can the functional response best be determined? Oecologia 76:206-214. [ Links ]
WALDE, SJ & WW MURDOCH. 1988. Spatial density dependence in parasitoids. Annu. Rev. Entomol. 33:441-446. [ Links ]
WILLIAMS, FM & SA JULIANO. 1985. Further difficulties in the analysis of functional response experiments and a resolution. Can. Entomol. 117:631-640. [ Links ]

