










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.14 n.2 Córdoba jul./dic. 2004
TRABAJOS ORIGINALES
Vegetation recovery on a gas-pipeline track along an altitudinal gradient in the Argentinean Yungas forests
Agustina Malizia*, Natacha P Chacoff, Héctor R Grau & Alejandro D Brown
Lab. de Inv. Ecológicas de las Yungas, Univ. Nac. de Tucumán. Casilla de Correo 34 (4107), Yerba Buena, Tucumán, Argentina. Tel/fax 54-381-4253728.
*Laboratorio de Investigaciones Ecológicas de las Yungas, Universidad Nacional de Tucumán, Tucumán, Argentina.
Email: agustinamalizia@yahoo.com.ar
Email: liey@arnet.com.ar
Recibido: 28 de octubre de 2003;
Fin de arbitraje: 15 de diciembre de 2003;
Revisión recibida: 26 de diciembre de 2003;
Aceptado: 16 de febrero de 2004
ABSTRACT. After one growing season of recovery, vegetation cover and height, species richness, and life forms composition were surveyed in 15 sites located along a gas-pipeline track running through an altitudinal range from 400-2000 m in the subtropical mountains of north-western Argentina (23°S). Vegetation cover was negatively correlated with altitude but was generally high at all sites (> 60%) after one year. Total species richness and maximum vegetation height did not vary significantly with altitude. Cover of grasses, and cover and species richness of trees, small shrubs and climbers were negatively correlated with altitude. Herb species richness correlated positively with altitude. Large shrub richness and cover showed no statistical relationship with altitude. The relative cover of herbs and grass species richness did not vary along the altitudinal gradient. Overall, these results indicate that in the altitudinal range studied, vegetation recovery is relatively high after this type of disturbance, probably due to low dispersal limitations and to the availability of species well adapted to intense disturbances. Vegetation recovery after gas-pipeline construction or similar perturbations may lead to relatively fast ecological restoration in subtropical montane forest ecosystems.
Keywords: Disturbance; Environmental impact; Life forms; Revegetation; Subtropical montane forest.
RESUMEN. Recuperación de la vegetación sobre la traza de un gasoducto en un gradiente altitudinal de las Yungas argentinas: La cobertura, altura de la vegetación y riqueza de especie total y para diferentes formas de vida después de una estación de crecimiento fueron censados en 15 sitios ubicados sobre la traza de un gasoducto en un gradiente altitudinal entre 400-2000 m en las montañas del noroeste argentino (23°S). La cobertura de la vegetación disminuyó con la altitud, aunque fue alta en todos los sitios (> 60%). La riqueza total de especies y la altura máxima de la vegetación no variaron significativamente con la altitud. La cobertura de pastos y la cobertura y riqueza de especies de árboles, pequeños arbustos y lianas disminuyeron con la altitud, mientras que la riqueza de hierbas aumentó. Tanto la riqueza y cobertura de arbustos, como la cobertura de hierbas y la riqueza de pastos no mostraron variaciones. Si bien la cobertura o riqueza de la mayoría de las formas de vida disminuyeron con la altitud, los resultados de este estudio sugieren que en el gradiente altitudinal estudiado la recuperación de la vegetación es relativamente rápida en todo el gradiente luego de perturbaciones humanas de este tipo, probablemente por la baja limitación a la dispersión y por la disponibilidad de especies adaptadas a perturbaciones intensas. La recuperación de la vegetación luego de la construcción de gasoductos o perturbaciones similares puede permitir restauraciones ecológicas relativamente rápidas en bosques subtropicales de montaña.
Palabras clave: Disturbios; Impacto ambiental; Formas de vida; Revegetación; Selvas subtropicales de montaña.
INTRODUCTION
Understanding and quantifying revegetation after human disturbances are important goals for basic research and ecological restoration purposes. Vegetation recovery affects important ecosystem properties such as soil erosion and fertility, and habitat quality (Borman & Likens 1979; Vitousek & White 1981; Peet 1992). Plant succession after human disturbances has been studied in Neotropical mountains in relation to traditional large-scale land uses such as grazing and agriculture. In those conditions, the main factors limiting early succession are dispersal limitation, competition with pre-existing grasses, and recurrent fires (Aide & Cavelier 1994; Sarmiento 1997; Holl et al. 2000; Wijdeven & Kuzee 2000; Zimmerman et al. 2000). While in several tropical mountains demographic and socio-economic factors are currently leading towards a reduction in grazing and agriculture, linear human disturbances such as roads and pipelines are increasingly widespread (Young 1994; Messerli & Ives 1997). Environmental impact assessment of these types of disturbances is becoming a common component of middle or large-scale development projects (Barrow 1997). However, impact assessments often lack basic information on dynamic processes such as vegetation recovery, which are needed to estimate ecosystem resilience and long-term ecological impacts.
In mountain ecosystems, roads or pipelines typically cover hundreds to thousands of meters of altitudinal range. Altitudinal gradients are recognized as a major control of vegetation structure in montane ecosystems. The changes in vegetation along elevational gradients are the consequence of the interaction among different factors, including different climatic effects on plant ecophysiology (Woodward 1987) and different disturbance regimes affecting vegetation (Harmon et al. 1984; Veblen et al. 1992). Given that revegetation rates depend on environmental conditions controlling establishment and growth, and on availability of propagules (Pickett et al. 1987), and that both factors vary with altitude, changes in vegetation recovery are also expected.
Studies on vegetation recovery after pipeline-related disturbances have been restricted to temperate and boreal ecosystems where the factors controlling revegetation (e.g., soil, climate, flora) are significantly different than those in Neotropical environments (Artz 1989; Zink et al. 1995; Paschke et al. 2000). We took advantage of a gas pipeline finished in 1999 in a Neotropical ecosystem of north-western Argentina to address two research objectives. The first was to quantify vegetation cover, height and species richness after one year of recovery. These variables are important for ecological restoration purposes since they influence soil erosion, habitat quality, and species diversity maintenance. We hypothesized that vegetation recovery of these variables decreases with altitude since vegetation growth is expected to decrease with decreased temperature along the altitudinal gradient. The second objective was to assess changes in cover and species richness of the different life forms. Different life forms are expected to respond differentially to the altitudinal gradient since they are associated with different resource-use strategies (Smith et al. 1997).
Study area
The study was conducted along the Nor-Andino gas-pipeline, which extends from Pichanal (Argentina) to Mejillones (Chile). The pipeline crosses approximately 60 km of montane forests located in the eastern slope of the Santa Victoria mountain range (23°14´S;64°40´W to 23°11´S;64°89´W, Figure 1). This area is part of the southern Yungas or Selva tucumano-boliviana, the southernmost extension of the tropical Andean montane forests (Cabrera & Willink 1980). The Nor-Andino gas pipeline is located in the core area of the upper Bermejo basin, considered the area of highest conservation priority of the Argentinean Yungas (Grau & Brown 2000; Brown et al. 2001). Climate is subtropical with a monsoonal rainfall regime (wet summers, dry winters). Mean annual temperature for the study area ranges from 14-21° C (Minetti 1999). Rainfall varies between 1000-2200 mm/year and peaks at intermediate altitudes, approximately 1000 m (Bianchi 1981).

Figure 1. (A) Relative location of the study area, (B) satellite image of the study transect, and (C) altitudinal profile showing the relative horizontal and vertical location of the sampling sites along the transect.
Figura 1. (A) Ubicación relativa del área de estudio, (B) imagen satelital de la transecta de estudio, y (C) ubicación relativa horizontal y vertical de los sitios a lo largo de la transecta.
Plant species composition, diversity and physiognomy change along the altitudinal range (400-2500 m). Three main forest types can be differentiated: (1) premontane lowland forests (400-700 m) dominated by Astronium urundeuva (Allemão) Engl., Phyllostylon rhamnoides (Poisson) Taubert, Parapiptadenia excelsa (Griseb.) Burkart, Anadenanthera colubrina (Vell.) Brenan, and Calycophylum mutiflorum Griseb.; (2) lower montane forests (700-1500 m) dominated by Blepharocalyx salicifolius (Kunth) O. Berg, Cinnamomum porphyrium (Griseb.) Kosterm., Ocotea puberula (Rich.) Nees, Ficus maroma Castellanos, and different species of Myrtaceae; and (3) upper temperate montane forests (1500-2500 m) with Cedrela lilloi C. DC., Juglans australis Griseb., Podocarpus parlatorei Pilg., and Alnus acuminata Kunth as the main tree species (Grau & Brown 2000).
At lower elevations (below 1200-1500 m) forests are semi-evergreen with 25-35 tree species per hectare (Grau & Brown 1995a). Closecanopy forests cover landscape, and climbers show the highest abundance and richness at this altitudinal level (A Malizia, pers. obs.). In the upper portion of the gradient, the vegetation is a mosaic of grasslands, shrublands and open-canopy forests. At the local scale (e.g., 1000 m2 transects), tree species richness decreases linearly with elevation (Morales 1996), whereas at the scale of 1 ha plots tree species richness shows relatively minor changes with elevation (Grau & Brown 1995a). In this region, no study has quantified altitudinal trends of climbers, grasses, shrubs and herbs.
The disturbance regime is dominated by treefall gaps, landslides and floods in the riparian areas at lower elevations, and by landslides and fire towards the upper elevations (HR Grau, in press). Several tree species regenerate in conditions of intense disturbances such as landslides (Grau & Brown 1995b).
The construction of the pipeline started in July 1998 and took approximately 16 months. To lay the pipeline, a 7-15 m wide road was opened, and all vegetation and topsoil were removed. The 56-cm diameter pipeline was buried at an average of 1.5 m of depth. Between September and October 1999, the road was abandoned progressively from lower to higher altitudes. Given that the road was abandoned at the end of the dry season when there is virtually no regeneration, we analyzed data assuming a simultaneous abandonment.
METHODS
Fifteen sampling sites were surveyed on the pipeline track along the studied altitudinal gradient (Figure 1). These sites were selected before the road was abandoned according to their accessibility to evaluate vegetation recovery. Data on vegetation recovery was collected in December 2000, one growing season after total vegetation removal.
At each sampling site, elevation and location were recorded using a Geographic Positioning System, and five transects were set perpendicular to the pipeline track. The first one was set on the pipeline track at least 20 m away from the access road, and transects were at least 10 m apart from each other. Each transect covered the total open belt (7-15 m). Along each transect, a vertical measuring stick was set every meter and a 5-cm radius was established around it. Thus, 10-cm diameter circular plots were established every meter along transects. In each plot, every plant inside the circle was identified to species and the life form type was recorded. Species were identified in the field or material was collected and identified using the literature (Cabrera 1978, 1983; Legname 1982; Gentry 1993; Jankowski et al. 2000) and reference collections from the Miguel Lillo Herbarium (Universidad Nacional de Tucumán, Argentina). Species nomenclature follows Zuloaga & Morrone (1999a, 1999b). Life forms were classified into six categories: herbs, grasses, climbers (herbaceous vines and woody lianas), trees, small shrubs, and large shrubs. Small shrubs were defined as plants with a woody base and herbaceous offshoots, while large shrubs have woody stems throughout. For practical purposes, the height of each individual was assigned to one of seven classes: 1.0-10.0 cm, 10.1-20.0 cm, 20.1-40.0 cm, 40.1- 60.0 cm, 60.1-100.0 cm, 100.1-200.0 cm, and 200.1-400.0 cm.
In order to estimate maximum vegetation height, we only considered the 10 tallest individuals per site. As plant height was assigned to classes, we used the mean value of its height class (e.g., if the individual was in the 200.1-400.0 cm class, we used 300 cm as its height). Finally, we computed the average of the 10 tallest values obtained by this procedure. Total percentage of cover and percentage of cover of each life form was computed as the number of circular plots with at least one individual taller than 1 cm divided by the total number of plots per site. Species richness is expected to change with the area of analysis. To estimate species richness in a comparative way among sites including transects of different length, we used the number of species included in the first seven plots of each transect, which was the number of plots of the shortest transect of our study. Thus, total species richness and species richness per life form correspond to the number of species found in an area of 350 cm2 per site (70 cm2 per transect, five transects per site). To assess the representativeness of this sample size as an index of species richness per site we used the rarefaction solution (Gotelli & Entsminger 2001). The method uses a specified number of individuals (which corresponds to the minimum abundance of individuals across all sites), to randomly select individuals from the complete set of data of each site, and generates new samples for which species richness is calculated. The process is repeated many times (1000 iterations) computing the mean and variance of species richness. In 9 of 15 sites, our estimated species richness index where included in the 95% confidence intervals of the rarefied species richness values, 5 were one or two species out of the intervals, and one was 4 species deviated. Moreover, the slope of the relationship between these rarefied species richness values and the altitudinal gradient was identical to the one reported in our results. Therefore we assumed that the sample size that we selected as an index of species richness is accurate. For comparative purposes and to avoid confusion in the interpretation of the results of different life forms, we used the same sample size for species richness of each life form and total species richness, although we did not apply rarefaction to species richness per life form because in many cases (e.g., climbers, trees, and small and large shrubs) the minimum abundance value was equal to 1. Finally, absolute frequency of species per site was reported.
Given that many variables did not meet the requirements of parametric Correlation Analysis (normality and homoscedasticity), Kendall´s Non-Parametric Correlation Analysis was used to assess the statistical significance of the variable-altitude relationships. The Slope Coefficient (b) of a Linear Regression Analysis is also presented for descriptive purposes. Descriptions of the statistical methods can be found in Sokal & Rohlf (1995).
RESULTS
We recorded a total of 116 species along the altitudinal gradient: 53 herbs, 21 trees, 17 large shrubs, 11 grasses, 10 climbers, and 4 small shrubs.
Maximum vegetation height and percent vegetation cover had a negative relationship with altitude. Maximum vegetation height decreased at a rate of about 70 cm in 1600 m, but the correlation was not statistically significant due to a relatively high inter-site variability (Figure 2A). At all sites vegetation cover after one growing season was greater than 60%, and decreased approximately 30% from 400 to 2000 m (Figure 2B). There was no statistical relationship between species richness and altitude (Figure 2C).

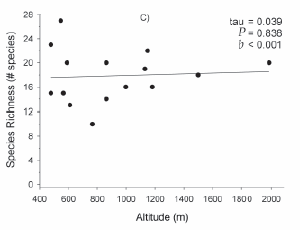
Figure 2. (A) Maximum vegetation height, (B) vegetation cover, and (C) total species richness (species/350 cm2) along the altitudinal gradient after one year of regeneration. Results of Non-Parametric Kendall´s Correlation Analysis are presented together with the slope parameter (b) of a Linear Regression.
Figura 2. (A) Altura máxima de la vegetación, (B) cobertura de la vegetación, y (C) riqueza total de especies (especies/350 cm2) a lo largo del gradiente altitudinal después de una estación de crecimiento. Se presentan los valores del Análisis de Correlación no Paramétrica de Kendall junto a los valores de la pendiente (b) de una Regresión Lineal.
Cover and richness of tree saplings, small shrubs and climbers showed a decreasing trend, although the trend for climber richness was only marginally significant (Figure 3A, 3B and 3D). Grass cover showed a steep decrease with altitude, reaching the highest values (approximately 60%) at low altitudes (Figure 3F). There was no relationship between grass species richness and altitude (Figure 3F), and herbs were the only life form that had an increase in richness with altitude (Figure 3E). There was no relationship between altitude and herb cover (Figure 3E), large shrub cover, and large shrub richness (Figure 3C).


Figure 3. Species richness (species/350 cm2) and vegetation cover of tree saplings (A), small shrubs (B), large shrubs (C), climbers (D), herbs (E), and grasses (F) along the altitudinal gradient after one year of regeneration. Results of Non-Parametric Kendall´s Correlation Analysis are presented together with the slope parameter (b) of a Linear Regression.
Figura 3. Riqueza de especies (especies/350 cm2) y cobertura de la vegetación correspondiente a árboles (A), arbustos pequeños (B), arbustos grandes (C), lianas (D), hierbas (E) y pastos (F) a lo largo del gradiente altitudinal después de una estación de crecimiento. Se presentan los valores del Análisis de Correlación no Paramétrica de Kendall junto a los valores de la pendiente (b) de una Regresión Lineal.
Among the grasses, Cynodon dactylon was very frequent at lower altitude sites while Paspalum dilatatum was very frequent along the entire gradient. Asteraceae was the most diverse herb family recorded. Most species of Asteraceae were very frequent at upper portions of the gradient, with the exception of Conyza bonariensis, which was more frequent at lower altitudes. Other herb species such as Commelina diffusa (Commelinanceae) were also frequent towards higher sites, while Kyllinga vagianata (Cyperaceae) was common the entire gradient throughout. The most abundant small shrub was Solanum chaetophorum (Solanaceae), which showed high frequencies at lower altitude sites. Tree, climber and large shrub species showed no clear frequency patterns along the gradient.
The proportion of cover by herbs and large shrubs increased with elevation, while the other four life forms tended to decrease (Figure 4).

Figure 4. Relative cover of the six life forms along the altitudinal gradient.
Figura 4. Cobertura relativa de las seis formas de vida a lo largo del gradiente altitudinal.
DISCUSSION
Even though vegetation recovery was very high in all sites (> 60% cover), there was a significant negative relationship with altitude as we hypothesized. These are high values in comparison with vegetation recovery after similar disturbances in temperate ecosystems (Artz 1989; Zink et al. 1995; Paschke et al. 2000). In addition, these cover values are the result of colonization by a diverse set of life forms, implying a faster recovery of functional diversity. This pattern of rapid recovery can be attributed to the short distance to seed sources due to the linear morphology of the pipeline track. Although seed arrival is often a major limitation for forest recovery in Neotropical mountains (e.g., Aide & Cavelier 1994; Myster & Sarmiento 1998), the relatively short distance to potential seed sources along the gas-pipeline track seems to allow early seed arrival without significant limitations. Given the high inter-site variation, recovery of vegetation height and richness did not show a clear relationship with the altitudinal gradient.
Differences in recovery of the different life forms were generally consistent with the ecology of those life forms along the altitudinal gradient, in which two groups of responses can be identified both for recovery of species richness and vegetation cover. In the case of species richness, life forms that dominate the landscape towards the lower portion of the gradient (trees and climbers), as well as small shrubs, showed a decreasing trend with elevation, as expected from the change from forest to open woodlands. Herbaceous plants consistently increased with altitude. Recovery of vegetation cover showed a similar pattern with the only difference that herb cover was relatively constant along the gradient and grass cover showed a decreasing trend with altitude, contrary to what one should expect from the vegetation gradient, in which grasses are the dominant life form towards the upper portion of the gradient. These results suggest that, while in the lower portion of the gradient grasses behave as pioneer plants that will be progressively displaced by the dominant life forms (primarily trees), in the upper portion of the gradient grasses could be considered late successional species that will replace herbs through time.
Two main conclusions emerge. First, vegetation recovery of plant cover is consistent with observed worldwide vegetation patterns along altitudinal gradients (Woodward 1987). Since temperature decreases with altitude, growth rates and regeneration are lower. Such a pattern is clearer for life forms typical of less extreme environments: trees and climbers. Second, despite the intense local disturbance produced by the pipeline construction, post-disturbance recovery is rapid. Vegetation ground cover, a main control of soil erosion, exceeds 60% at all elevations after only one growing season. This can be largely related to the fact that, although the disturbance was intense, the disturbed area is narrow, and consequently seed dispersal is not a major limitation for recovery. In abandoned pastures the two main factors limiting vegetation restoration are availability of propagules and competition with preexisting grasses (Guevara et al. 1986; Aide & Cavelier 1994; Zimmerman et al. 2000). The rapid regeneration in our study was mainly due to the absence of these factors. In our study, seed input was not limited due to the proximity to seed sources, and competition with preexisting vegetation did not occur since the disturbance removed all vegetation. In addition, the disturbance regime is intense in these ecosystems, where landslides are common components of forest dynamics (Grau & Brown 1995a; HR Grau, in press). Landslides may be promoting the availability of disturbance-adapted species, making patterns of diversity recovery consistent with models of small disturbances (Miller 1982) or disturbance-adapted vegetation (Denslow 1985). The only exception to this pattern seems to be grasses, which behaved as pioneer/early colonizers at lower elevations, and lagged behind herbs at higher elevations, where herbs dominated the undisturbed landscape.
Overall, the rapid vegetation recovery appears to be driven by efficient dispersal, lack of competition with pre-existing vegetation, low compaction of the soil and capacity of local plant communities to colonize severely disturbed sites. This suggests that intense linear disturbances, such as pipeline or similar constructions (e.g., short lasting, low traffic roads with low soil compaction), could be managed and restored with native vegetation in a relative short time period. The first year after the disturbance seems to play a significant role in colonization and potentially on successional pathways making management decisions during this period particularly relevant. At higher elevations, revegetation is slower and restoration may need more aid, for example by planting life forms that are important components of the landscape, such as grasses.
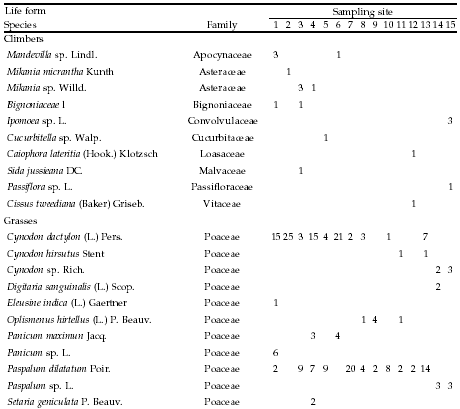
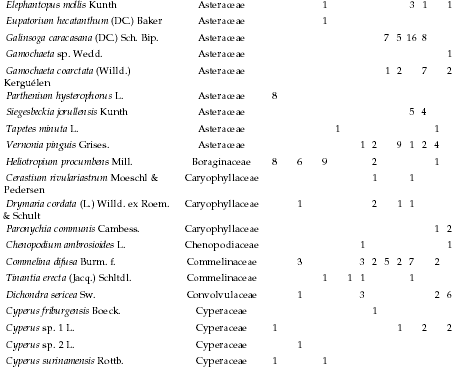
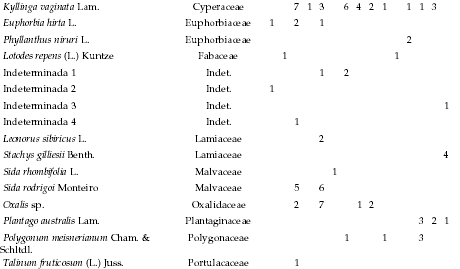
Appendix
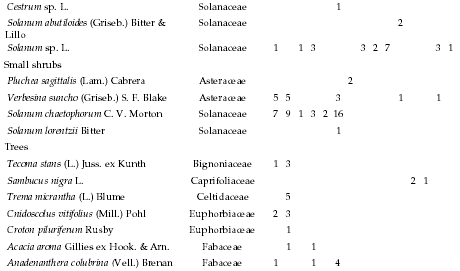
Absolute frequency of occurrence for species recorded along the altitudinal gradient, listed by life form, per sampling site. Each site consisted of 35 plots (five 7-m transects). 1: 478 m, 2: 478 m, 3: 546 m, 4: 565 m, 5: 590 m, 6: 610 m, 7: 765 m, 8: 863 m, 9: 863 m, 10: 998 m, 11: 1130 m, 12: 1148 m, 13: 1182 m, 14: 1500 m, and 15: 1985 m.
Frecuencia absoluta de la presencia en los sitios de muestreo de las especies identificadas a lo largo del gradiente altitudinal, listadas por forma de vida. Cada sitio incluyó 35 parcelas (cinco transectas de 7 m). 1: 478 m, 2: 478 m, 3: 546 m, 4: 565 m, 5: 590 m, 6: 610 m, 7: 765 m, 8: 863 m, 9: 863 m, 10: 998 m, 11: 1130 m, 12: 1148 m, 13: 1182 m, 14: 1500 m, and 15: 1985 m.



ACKNOWLEDGEMENTS
To Karina Buzza and Yamila Sasal for helping at several stages of the fieldwork, to Alberto Slanis for identifying the botanical specimens, and to Alfredo Grau, Jorge Rabinovich, Mitchell Aide, Pablo Jayat and three anonymous reviewers for helpful comments on the manuscript. This project was supported by Nor-Andino S.A.
REFERENCES
AIDE, TM & J CAVELIER. 1994. Barriers to lowland tropical forest restoration in the Sierra Nevada de Santa Marta, Colombia. Restor. Ecol. 2:219-229. [ Links ]
ARTZ, MC. 1989. Impact of linear corridors on perennial vegetation in the east Mojave desert: implications for environmental management and planning. Nat. Areas J. 9:117-129. [ Links ]
BARROW, CJ. 1997. Environmental and social impact assessment. Arnold. Londres. [ Links ]
BIANCHI, AR. 1981. Las precipitaciones del noroeste argentino. Instituto Nacional de Tecnología Agropecuaria. Salta. [ Links ]
BORMAN, FH & GE LIKENS. 1979. Pattern and process in a forested ecosystem. Springer-Verlag. New York. [ Links ]
BROWN, AD; HR GRAU; A GRAU & L MALIZIA. 2001. Argentina. Pp. 623-660 in: M Kappelle & AD Brown (eds). Bosques nublados del Neotrópico. Instituto Nacional de Biodiversidad. San Jose. [ Links ]
CABRERA, AL. 1978. Flora de la Provincia de Jujuy. Parte X: Compositae. Instituto Nacional de Tecnología Agropecuaria. Buenos Aires. [ Links ]
CABRERA, AL. 1983. Flora de la Provincia de Jujuy. Parte VIII: Clethraceas a Solanaceas. Instituto Nacional de Tecnología Agropecuaria. Buenos Aires. [ Links ]
CABRERA, AL & A WILLINK. 1980. Biogeografía de America Latina. 2nd edn. OEA. Washington DC. [ Links ]
DENSLOW, JS. 1985. Disturbance-mediated coexistence of species. Pp. 63-82 in: STA Pickett & PS White (eds). The ecology of natural disturbances and patch dynamics. Academic press. Orlando. [ Links ]
GENTRY, AH. 1993. A Field guide to the families and genera of woody plants of northwest South America (Colombia, Ecuador, Peru) – with supplementary notes on herbaceous taxa. Conservation International. Washington DC. [ Links ]
GOTELLI, NJ & GL ENTSMINGER. 2001. ECOSIM: null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. URL: http:// homepages.together.net/~gentsmin/ecosim.htm. [ Links ]
GRAU, A & AD BROWN. 2000. Development threats to biodiversity and opportunities for conservation in the mountain ranges of the upper Bermejo river basin, NW Argentina and SW Bolivia. Ambio 29:445-450. [ Links ]
GRAU, HR. In press. Disturbios y sucesión en el gradiente altitudinal de las yungas argentinas. In: J Frangi & MF Arturi (eds). Manejo y conservación de los bosques argentinos. Universidad Nacional de La Plata. Buenos Aires. [ Links ]
GRAU, HR & AD BROWN. 1995a. Patterns of tree species diversity in altitudinal and latitudinal gradients in the Argentinean subtropical montane forests. Pp. 295-300 in: S Churchill; E Forero; J Luteyn & E Balslev (eds). Neotropical montane forests biodiversity and conservation. New York Botanical Garden Press. New York. [ Links ]
GRAU, HR & AD BROWN. 1995b. Los deslizamientos de ladera como condicionantes de la estructura y composición de la selva subtropical de montaña. Pp. 79-84 in: AD Brown & HR Grau (eds). Investigación, conservación y desarrollo en selvas subtropicales de montaña. Laboratorio de Investigaciones Ecológicas de las Yungas. Tucumán. [ Links ]
GUEVARA, S; SE PURATA & E VAN DER MAAREL. 1986. The role of remnant trees in tropical forest succession. Vegetatio 66:77-84. [ Links ]
HARMON, ME; SP BRATTON & PS WHITE. 1984. Disturbance and vegetation response in relation to environmental gradients in the Great Smoky Mountains. Vegetatio 55:129-139. [ Links ]
HOLL, KD; ME LOIK; EH LIN & IA SAMUELS. 2000. Tropical montane forest restoration in Costa Rica: overcoming barriers to dispersal and establishment. Restor. Ecol. 8:339-349. [ Links ]
JANKOWSKI, LS; D BAZZANO; M TOURN & G ROITMAN. 2000. Plantas trepadoras, nativas y exóticas. Literature of Latin America. Buenos Aires. [ Links ]
LEGNAME, PR. 1982. Árboles indígenas del noroeste argentino. Opera Lilloana 34:1-155. [ Links ]
MESSERLI, B & JD IVES. 1997. Mountains of the world, a global priority. Parthenon Pub. Cichester & París. [ Links ]
MILLER, TE. 1982. Community diversity and interactions between the size and frequency of disturbance. Am. Nat. 120:533-536. [ Links ]
MINETTI, JL. 1999. Atlas climático del noroeste argentino. Fundación Carl C: Zon Caldenius, Sede NOAA, Universidad Nacional de Tucumán. Tucumán. [ Links ]
MORALES, JM. 1996. Comunidades arbóreas no saturadas en las Yungas de Argentina. Ecología Austral 6:17-22. [ Links ]
MYSTER, RW & FO SARMIENTO. 1998. Seed inputs to microsite patch recovery on two tropandean landslide in Ecuador. Restor. Ecol. 6:35-43. [ Links ]
PASCHKE, KW; J DELEO & R EDENTE. 2000. Revegetation of roadcut slopes in Mesa Verde National Park, U.S.A. Restor. Ecol. 8:276-282. [ Links ]
PEET, RK. 1992. Community structure and ecosystem function. Pp. 234-248 in: DC Glenn Levin; RK Peet & TT Veblen (eds). Plant succession, theory and prediction. Chapman & Hall. Londres. [ Links ]
PICKETT, STA; SL COLLINS & JJ ARMESTO. 1987. Models, mechanisms and pathways of succession. Bot. Rev. 53:335-371. [ Links ]
SARMIENTO, FO. 1997. Landscape regeneration by seeds and successional pathways to restore fragile tropandean slopelands. Mt. Res. Dev. 17:239-252. [ Links ]
SMITH, TM; HH SHUGART & FI WOODWARD. 1997. Plant functional types. Their relevance to ecosystem properties and global change. Cambridge University Press. Cambridge. [ Links ]
SOKAL, RR & FJ ROHLF. 1995. Biometry. The principles and practice of statistics in the biological sciences. Freeman. New York. [ Links ]
VEBLEN, TT; T KITZBERGER & A LARA. 1992. Disturbance and forest dynamics along a transect from Andean rainforest to Patagonian shrubland. J. Veg. Sci. 3:507-520. [ Links ]
VITOUSEK, PM & PS WHITE. 1981. Process studies in succession. Pp. 267-276 in: DC West; HH Shugart & DB Botkin (eds). Forest succession: concepts and applications. Springer Verlag. New York. [ Links ]
WIJDEVEN, SMJ & ME KUZEE. 2000. Seed availability as a limiting factor in forest recovery processes in Costa Rica. Restor. Ecol. 8:414-424. [ Links ]
WOODWARD, FI. 1987. Climate and plant distribution. Cambridge University Press. Cambridge. [ Links ]
YOUNG, K. 1994. Roads and the environmental degradation of tropical montane forests. Conserv. Biol. 8:972-976. [ Links ]
ZIMMERMAN, JK; JB PASCARELLA & TM AIDE. 2000. Barriers to forest regeneration in an abandoned pasture in Puerto Rico. Restor. Ecol. 8:350-360. [ Links ]
ZINK, TA; MF ALLEN; B HEINDLI-TENHUNEN & EB ALLEN. 1995. The effect of a disturbance corridor on an ecological reserve. Restor. Ecol. 3:304-310. [ Links ]
ZULOAGA, FO & O MORRONE. 1999a. Catálogo de las plantas vasculares de la República Argentina. I. Missouri Botanical Garden Press. Missouri. [ Links ]
ZULOAGA, FO & O MORRONE. 1999b. Catálogo de las plantas vasculares de la Repuública Argentina. II. Missouri Botanical Garden Press. Missouri. [ Links ]

