










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.15 n.1 Córdoba ene./jun. 2005
TRABAJOS ORIGINALES
Intercambio gaseoso en dos especies de plantas alto andinas de Chile central: efecto de la asociación a plantas en cojín
Marco A Molina-Montenegro 1*, Ernesto I Badano1, Patricia A Inostroza2 & Lohengrin A Cavieres1
1 Grupo de Investigación en Ecología, Biogeografía y Sistemática (ECOBIOSIS), Departamento de Botánica, Universidad de Concepción, Concepción, Chile
2 Laboratorio de Fisiología Vegetal, Departamento de Botánica, Universidad de Concepción, Concepción, Chile
* Grupo de Investigación en Ecología, Biogeografía y Sistemática (ECOBIOSIS), Depto. de Botánica, Univ. de Concepción, Concepción, Chile. Email: marcmoli@udec.cl
Recibido: 16 de abril de 2003;
Fin de arbitraje: 29 de julio de 2003;
Revisión recibida: 14 de agosto de 2003;
Segunda revisión recibida: 21 de mayo de 2004;
Aceptado: 10 de junio de 2004
RESUMEN. Se ha propuesto que en ambientes extremos como los sistemas de alta montaña, las interacciones entre las especies tenderían a ser del tipo positivas y aumentarían tanto en intensidad como en frecuencia a medida que el ambiente se torna más limitante para la adquisición de recursos. En este trabajo, se estudiaron las modificaciones microclimáticas que realizan los cojines de la especie Laretia acaulis y el efecto de ésta sobre la respuesta fisiológica de dos especies de la comunidad de alta montaña situada a 2800 m.s.n.m. en Los Andes de Chile central: Taraxacum officinale y Euphorbia collina, las cuales crecen frecuentemente entre y fuera de los cojines respectivamente. Se realizaron mediciones de intercambio gaseoso en plantas creciendo sobre el cojín y en espacios abiertos con el objeto de evidenciar los efectos provocados por las modificaciones microclimaticas que realizan los cojines. Se evidenció que los individuos de T. officinale que crecen entre los cojines presentaban una mayor tasa de fotosíntesis neta que sus coespecificos que crecen en los espacios abiertos. E. collina no mostró diferencia en la tasa de fotosíntesis neta de los individuos presentes tanto entre como fuera de los cojines. El efecto de asociación con una nodriza (e.g. plantas en cojín) para especies de gran biomasa como E. collina sería menos beneficioso debido a la competencia por los recursos y el espacio, lo cual explicaría que esta especie se registrara frecuentemente en los espacios abiertos.
Palabras clave: Tasa de fotosintesis neta; Andes; Facilitación; Ecofisiología; Efecto nodriza.
ABSTRACT. Gas exchange in two high andean plant species of central Chile: effect of the association with cushion plants: It has been proposed that on stressful environments as high mountain habitats, interactions between species of plants would tend to be of the positive type, increasing in intensity and frequency when resources in the environment become more limited. In this work, we studied the microclimatic modifications produced by cushions of Laretia acaulis species and its effects on the physiological performance of two associated species of the high mountain community located at 2800 m.a.s.l. in Los Andes of central Chile: Taraxacum officinale and Euphorbia collina, which frequently grow within and outside the cushions respectively. Gas exchange measures were taken in order to assess the effects caused by the microclimatic modifications done by cushions plants. We demonstrated that individuals of T. officinale that grow within cushions displayed, unlike their co-specifics in open spaces, a greater net photosynthesis rate. E. collina did not show differences in the rate of net photosynthesis between individuals present within and outside cushions. Only T. officinale displayed a greater rate of photosynthesis in the microsite where it is frequently distributed (within cushions). The effect of association with a nurse (e.g. cushion plants) for large biomass species, such as E. collina, could be less beneficial due to the competition for resources and space; this would explain why E. collina is more frequently found in open spaces.
Keywords: Net photosynthesis rates; Andes; Facilitation; Ecophysiology; Nurse effect.
INTRODUCCIÓN
Las interacciones positivas son aquellas relaciones interespecíficas por las cuales una o más especies se ven beneficiadas sin perjuicio para la otra (Hunter & Aarssen 1988). Este tipo de interacciones han sido descritas principalmente en ambientes estresantes como las zonas áridas (Pugnaire et al. 1996), zonas de marismas (Bertness & Yeh 1994; Callaway & Pennings 2000) y de alta montaña (Núñez et al. 1999; Molina-Montenegro et al. 2000; Cavieres et al. 2002).
Los mecanismos por los cuales pueden actuar los procesos de facilitación tales como disminución de temperaturas extremas, mayor disponibilidad de agua y mayor concentración de nutrientes han sido descritos con anterioridad por diversos autores (Hager & Faggi 1990; Kikvidze 1996; Callaway & Pugnaire 1999; Núñez et al. 1999). Sin embargo, sólo en los últimos años se ha comenzado a estudiar los efectos de estos mecanismos en términos fisiológicos: asimilación de CO2, eficiencia fotosintética, eficiencia en el uso del agua y variación en los contenidos de pigmentos (Niinements et al. 1999; Loik & Holl 2001).
Ante una situación de estrés, es la fotosíntesis el proceso fisiológico con mayor repercusión sobre el aumento de biomasa de una planta. Las interacciones positivas podrían influir sobre este proceso, permitiendo un balance de carbono positivo (Kikvidze 1996). Los factores de estrés pueden actuar de manera directa o indirecta sobre la fotosíntesis, dependiendo de qué factor de estrés se trate y de la ruta metabólica sobre la que esté actuando (Lambers et al. 1998).
Se ha documentado que el estrés por déficit de agua provoca la oclusión de los estomas por parte de las células guardianas, ocasionando una disminución en la conductancia estomática y, en muchas especies, un déficit de CO2 para la fotosíntesis (Lambers et al. 1998). Se ha demostrado que algunas especies suelen tener un balance negativo en la captación de carbono, que se hace más marcado en periodos de sequía debido a que la fotosíntesis es muy baja en presencia de estrés hídrico (Körner 1999; Nardini et al. 1999).
Los ambientes de alta montaña se caracterizan por las bajas temperaturas del aire y el corto período favorable para el establecimiento de plántulas (Smith & Young 1987). Estas características, sumadas a los fuertes vientos y a la baja disponibilidad de agua, disminuyen la tasa de fotosíntesis limitando la adquisición de recursos para las plantas (Sakai & Larcher 1987; Körner 1999).
Las plantas en cojín son una de las formas de vida mejor adaptadas a las extremas condiciones de los ambientes de alta montaña (Alliende & Hoffmann 1985; Pysek & Lyska 1991). Su particular arquitectura achaparrada modifica los patrones microclimáticos que se generan entre y bajo su dosel (Körner & De Moraes 1979; Körner & Cochrane 1983). Por ejemplo, Hager & Faggi (1990) han demostrado que las plantas en cojín disminuyen la velocidad del viento entre su dosel, lo que disminuiría la evapotranspiración y la pérdida de calor por convección permitiendo, de esta manera, que tanto la temperatura como la humedad sean mayores entre los cojines que fuera de ellos. Por otro lado, Cavieres et al. (1998) demostraron que en una comunidad alto andina en los Andes de Chile, existiría un mayor porcentaje de humedad bajo plantas en cojín que en el suelo aledaño a ellas. De manera similar Nuñez et al. (1999) demostraron en una comunidad de altamontaña, que la concentración de nutrientes en el suelo bajo plantas en cojín es mayor que en los espacios abiertos. De esta forma, las plantas en cojín al proporcionar micrositios más adecuados para la adquisición de recursos, actuarían como nodrizas para el resto de las especies de la comunidad (Callaghan 1987).
El presente trabajo tiene como objetivo comparar el desempeño fisiológico de dos especies de plantas de altamontaña: Taraxacum officinale (Asteraceae) y Euphorbia collina (Euphorbiaceae), que crecen tanto entre los cojines de la especie Laretia acaulis (Apiaceae) como en el suelo aledaño a ellos, a los 2800 m.s.n.m. en los Andes de Chile central. Debido a las modificaciones microclimáticas que realizan los cojines, sería esperable que los individuos de ambas especies que crecen en asociación con la nodriza presenten una mayor tasa de fotosíntesis neta que los individuos co-específicos presentes en un microhábitat más estresante como lo son los espacios abiertos.
MATERIALES Y MÉTODOS
Sitio de estudio
Este trabajo se realizó en la localidad de La Parva, situada en los Andes de Chile central. El sitio de estudio se ubica a los 2800 m.s.n.m. (33º19´S; 70º16´O), a 70 Km al este de la ciudad de Santiago, sobre una ladera de exposicion S-E (120º con respecto al norte) con una pendiente de 35º y un sustrato que presenta un 29.0% de arcilla y 1.8% de roca (Cavieres & Arroyo 2000). En general, el clima de Chile central es de tipo Mediterráneo con estacionalidad en las temperaturas y precipitaciones, y una moderada amplitud térmica entre el período invernal y la época estival (di Castri & Hajek 1976).
La localidad de La Parva presenta precipitaciones medias anuales de 400-900 mm, siendo éstas, principalmente nivales y concentradas en los meses de invierno (junio-julio-agosto). Las temperaturas mínimas promedio alcanzan los - 4.1ºC en el mes de junio, y las mínimas absolutas llegan a los -12.5ºC en el mes de agosto. La temperatura máxima promedio es de 15.5ºC y la máxima absoluta se presenta con 29.0ºC, ambas registradas en el mes de octubre (Cavieres & Arroyo 1999). El sitio de estudio presenta una vegetación dominada por la especie en cojín Laretia acaulis (Cavieres & Arroyo 2000), y también están presentes en la comunidad especies como Thlaspi magellanicum, Euphorbia collina, Perezia carthamoides, Cerastium arvense y Taraxacum officinale.
Especies estudiadas
Laretia acaulis, (Cav.) Gillies & Hook (Apiaceae): habita en las altas cordilleras de Coquimbo a Talca, entre los 2400-3200 m.s.n.m. Es una planta perenne agrupada, de crecimiento muy lento, dispuesta en céspedes densos. Posee hojas resinosas, arrosetadas de 1.5-2.5 cm de largo. Flores amarillas, pequeñas, agrupadas en umbelas de 6-10. Floración de diciembre a enero.
Taraxacum officinale, Weber (Asteraceae): esta especie es una maleza cosmopolita de origen europeo. Es una planta herbacea perenne, rizomatosa, sin tallo, con hojas dispuestas en roseta, oblongas, pinnatisectas, con segmentos enteros o dentados, glabras o algo híspidas. Habita en laderas de cerros, zonas de cultivos y sectores urbanos.
Euphorbia collina, Phil. (Euphorbiaceae): planta perenne, glabra. Tallos de 15-30 cm de altura, ascendentes, ramificados, que contienen látex blanco. Hojas remanentes en la base, elípticas a obovado-espatuladas. Habita principalmente en la cordillera chilena, desde Coquimbo hasta Linares, con presencia también en la Patagonia Argentina (Hoffmann et al. 1998).
Análisis de aleatorización
Con el objeto de comparar la abundancia tanto de T. officinale como de E. collina, se analizaron en los dos micrositios las frecuencias observadas de las especies creciendo entre los cojines y los espacios abiertos con frecuencias generadas por azar (Slade & Hall 1999; Badano et al. 2002; Cavieres et al. 2002). Debido a que estos análisis requieren que el esfuerzo de muestreo sea el mismo en ambas posiciones a comparar, se consideraron por separado las muestras pareadas (entre-fuera) tomadas para cada especie. Las distribuciones de frecuencias al azar se generaron con el software Resampling Stats (Kikvidze et al. 2001; Badano et al. 2002), considerando 1000 remuestreos aleatorios de las frecuencias observadas de cada especie en las muestras. Posteriormente, se calculó la probabilidad con que la distribución de frecuencia observada para cada individuo podría atribuirse a una distribución al azar. Sería esperable que los individuos tanto de T. officinale como de E. collina se distribuyan 50% en cada micrositio, sin embargo la mayor frecuencia de ocurrencia de una especie en un micrositio dado podría estar explicada por el mayor desempeño fisiologico obtenido en el mismo.
Caracterización microclimática de los sustratos
Para analizar las modificaciones microclimáticas que induce L. acaulis con respecto a los espacios abiertos, se midió el potencial hídrico matricial del suelo y la temperatura del sustrato. El potencial hídrico se midió a 10 cm de profundidad con un tensiómetro 2725 Series Jet Fill Tensiometer bajo tres cojines al azar de L. acaulis que tuvieran un diámetro superior a 20 cm. Las mediciones de temperatura se realizaron a 1 cm de profundidad con un termómetro digital Digital Thermometer 871A entre tres cojines al azar con intervalos de dos horas entre las 08:00 y 20:00 horas. A fin de tener información comparable, por cada muestra de humedad y temperatura sobre los cojines se tomó una similar en los espacios abiertos. Los datos de humedad y temperatura fueron analizados con un análisis de varianza (ANOVA) de una vía y de mediciones repetidas, respectivamente.
Variables fisiológicas y microclimaticas foliares analizadas
Por cada micrositio (entre y fuera del cojín), se seleccionaron tres individuos adultos de T. officinale y de E. collina que presentaran tamaño y morfología foliar similar (Tabla 2). Se midió la tasa de fotosíntesis neta, la conductancia estomática, la temperatura foliar, la tasa de transpiración y la radiación fotosintéticamente activa (PAR) incidente sobre la lámina foliar. Las mediciones fueron realizadas sobre el mismo individuo a las 08:00, 11:00, 14:00 y 17:00 horas mediante un analizador infrarrojo de gases IRGA, Infra Red Gas Analyser, CIRAS- 1, PP-Systems Haverhill. A partir de estos valores se estimó la eficiencia en el uso del agua para la fotosíntesis (Photosyntetic Water Use Efficiency, PWUE) como el cociente entre los valores de la tasa fotosintética y la transpiración (A/E). Según Lambers et. al (1998), este parámetro sería utilizado como indicador del estado de estrés hídrico de una planta en un micrositio determinado.
Los resultados fueron analizados con un análisis de varianza (ANOVA) de mediciones repetidas, donde los factores a considerar fueron la posición dentro-fuera y la hora del día a la cual se tomaron las mediciones. Este análisis permitiría evidenciar si los individuos de una especie presentan un mejor desempeño fisiológico en un determinado micrositio y cómo esta diferencia varía en un ciclo diario.
Este trabajo se llevó a cabo en el mes de abril de 2002, ya que los últimos meses de la temporada de crecimiento (antes de las primeras heladas) se corresponde con el período de máximo estrés hidrico (Körner & Larcher 1988).
RESULTADOS
Análisis de aleatorización
Los resultados del analisis de randomización, indicaron que la frecuencia de encuentro de individuos, tanto de T. officinale como de E. collina, difiere del azar (Tabla 1). T. officinale fue registrada en 41 cojines de L. acaulis y en 22 muestras en los espacios abiertos. E. collina fue registrada 11 veces creciendo entre los cojines, mientras que en los espacios abiertos se registró en 33 muestras (Tabla 1). Las frecuencias de encuentro para ambas especies, fueron determinadas de un total de 50 muestras por cada micrositio.
Tabla 1. Frecuencia de Taraxacum officinale y Euphorbia collina creciendo entre L. acaulis y en espacios abiertos (n = 100)(cincuenta entre cojines de L. acaulis y cincuenta en los espacios abiertos) en la zona de Los Andes a 2800 m.s.n.m. P es el valor de la prueba de aleatorización, se consideró significativo los valores con P < 0.05.
Table 1. Frequency of Taraxacum officinale and Euphorbia collina growing within L. acaulis and in open spaces (n = 100) (fifty whithin cushions plant L. acaulis species and fifty in open spaces) in Los Andes at 2800 m.a.s.l. P is the value of randomization test, considered as significant when P < 0.05.

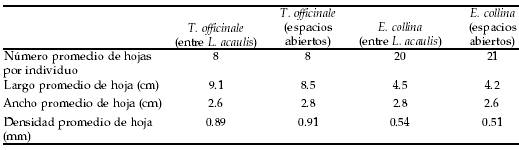
Tabla 2. Características morfológicas de los individuos y de las hojas de Taraxacum officinale y Euphorbia collina utilizadas para las mediciones de la tasa de intercambio gaseoso, a 2800 m.s.n.m. en Los Andes de Chile central (33ºS). (Se muestran los valores de ± 1 DS)
Table 2. Morphologic characteristics of the individuals and leaves of Taraxacum officinale and Euphorbia collina used for the measurements of the gas exchange rate at 2800 m.a.s.l. in Los Andes of central Chile (33ºS). (± 1 SD values are shown).
Caracterización microclimática de los sustratos
El suelo bajo Laretia acaulis presentó potenciales hídricos significativamente menores que en los espacios abiertos (F = 250.00, P < 0.0001), indicando que en el suelo debajo de los cojines hay una mayor cantidad de agua que en el de los espacios abiertos. Esto indicaría que, hacia finales de la temporada de crecimiento, los cojines de L. acaulis serían el micrositio con mayor humedad (Figura 1).
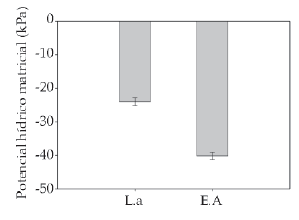
Figura 1. Promedios del potencial hídrico matricial (± 1 DS) en el sustrato debajo de Laretia acaulis (L.a) y los espacios abiertos (E.A) a los 2800 m.s.n.m. en Los Andes de Chile central (33ºS).
Figure 1. Matrix water potential averages (± 1 SD) in the ground below Laretia acaulis (L.a) and the open spaces (E.A) at 2800 m.a.s.l. in Los Andes of central Chile (33ºS).
El test de ANOVA de mediciones repetidas mostró diferencias significativas (F = 192.71, P < 0.001) en las temperaturas del suelo (entre y fuera de los cojines); evidenció además diferencias significativas en el tiempo (F = 741.33, P < 0.001), manteniendo los cojines una temperatura menos oscilante durante el ciclo diario (Figura 2).
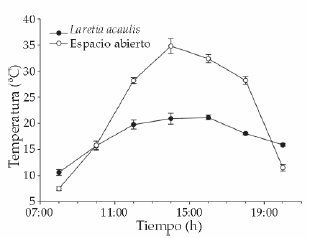
Figura 2. Perfil diario de temperatura entre cojines de Laretia acaulis (círculos negros) y los espacios abiertos (círculos blancos) (± 1 DS) medidos a 1 cm de profundidad a los 2800 m.s.n.m., en Los Andes de Chile central (33ºS).
Figure 2. Daily profile of temperatures within cushions of Laretia acaulis (closed circles) and open spaces (open circles) (± 1 SD) measured at 1 cm underground, at 2800 m.a.s.l. in Los Andes of central Chile (33ºS).
Variables fisiológicas y microclimaticas foliares analizadas
Los individuos de T. officinale creciendo entre los cojines de L. acaulis presentaron tasas de fotosíntesis neta mayores (F = 29.57, P < 0.01) que sus congéneres en los espacios abiertos (Figura 3), y esa diferencia se mantuvo a lo largo del día. En correspondencia con ello, la conductancia estomática para el caso de T. officinale fue significativamente mayor (F = 29.00, P < 0.01) en los individuos presentes dentro de los cojines (Figura 3). Para el caso de E. collina, no se registraron diferencias en la tasa de fotosíntesis neta entre los individuos presentes en los dos micrositios (F = 0.014, P = 0.91) (Figura 4). Tampoco se registraron diferencias significativas (F = 0.003, P = 0.96) en los niveles de conductancia estomática con respecto a la posición (Figura 4). Tanto para T. officinale como para E. collina, la temperatura foliar y la radiacion (PAR) fue similar considerando los individuos presentes en los dos micrositios (dentro y fuera del cojín) (Figuras 3 y 4).
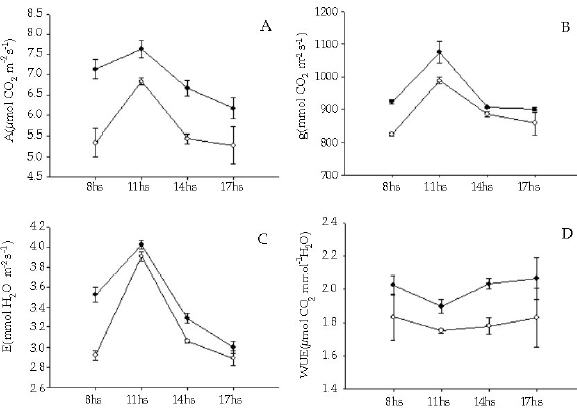
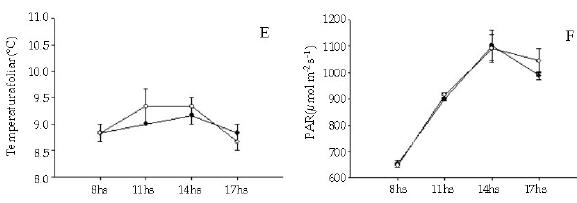
Figura 3. Ciclo diario del intercambio gaseoso (tasa de fotosintesis neta (a), conductancia estomática (b), transpiración (c) eficiencia en el uso del agua (d), temperatura foliar (e) y radiación fotosinteticamente activa (PAR)) en individuos de la especie Taraxacum officinale que crecen entre Laretia acaulis (círculos negros) y en los espacios abiertos (círculos blancos), a 2800 m.s.n.m. en Los Andes de Chile central (33ºS). Se muestran los valores de ± 1 DS.
Figure 3. Daily cycle of the gas exchange (net photosynthesis rates (a), stomatal conductance (b), transpiration, (c) water-use efficiency (d), leaf temperature (e) and photosynthetically active radiation (PhAR)) in individuals of Taraxacum officinale species that grow within Laretia acaulis (solid circles) and in open spaces (open circles), at 2800 m.a.s.l. in Los Andes of central Chile (33ºS). ± 1 SD values are shown.
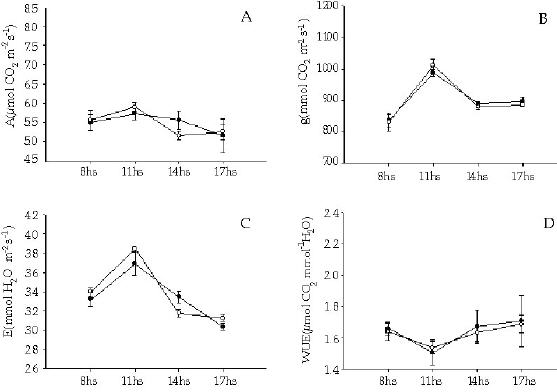
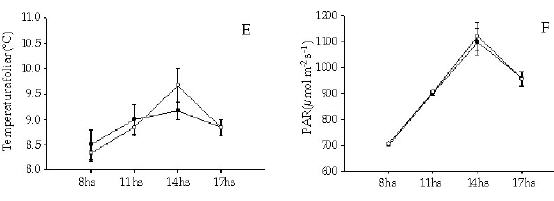
Figura 4. Ciclo diario del intercambio gaseoso (tasa de fotosintesis neta (a), conductancia estomática (b), transpiración (c), eficiencia en el uso del agua (d), temperatura foliar (e) y radiación fotosinteticamente activa (PAR)) en individuos de la especie Euphorbia collina que crecen entre Laretia acaulis (círculos negros) y en los espacios abiertos (círculos blancos), a los 2800 m.s.n.m. en Los Andes de Chile central (33ºS). Se muestran los valores de ± 1 DS.
Figure 4. Gas exchange daily cycle (net photosynthesis rates (a), stomatal conductance (b), transpiration (c), water-use efficiency (d), leaf temperature (e) and photosynthetically active radiation (PhAR)) in individuals of Euphorbia collina species growing within Laretia acaulis (closed circles) and in open spaces (open circles), at 2800 m.a.s.l. in Los Andes of central Chile (33ºS). ± 1 SD values are shown.
Al considerar la tasa de transpiración, se evidenció que solamente los individuos de T. officinale presentaron diferencias significativas entre los dos micrositios (F = 20.67, P < 0.01), siendo mayor la tasa de los individuos presentes entre los cojines (Figura 3).
Con respecto a la eficiencia en el uso del agua, no se encontraron diferencias significativas para ninguna de las dos especies (F = 5.22, P = 0.094) y (F = 0.18, P = 0.693) T. officinale y E. collina respectivamente. La alta varianza de los datos, debido posiblemente al bajo número de réplicas, no habría permitido evidenciar diferencias entre los individuos de ambas especies ubicadas en los distintos micrositios (Figuras 3 y 4).
DISCUSIÓN
Según Chambers et al. (1990), en los ambientes de alta montaña con buena disponibilidad de agua y con condiciones de temperatura más favorables que su entorno, varios procesos en las plantas serían facilitados.
Conjuntamente, otros autores sugieren que la disponibilidad de agua y la amortiguación de temperaturas extremas mejorarían procesos, principalmente del tipo fisiológico como la fotosíntesis (Kikvidze 1996).
De acuerdo a lo dicho anteriormente, y considerando que las plantas en cojín mantienen mayor humedad y temperaturas menos estresantes que su entorno, las mismas permitirían que las plantas que crecen asociadas a ellas mantengan una mayor tasa de fotosíntesis, incluso en periodos de sequía extrema (Kikvidze 1996; Körner 1999; Nardini et al. 1999). De esta manera, las especies menos tolerantes a las condiciones estresantes de alta montaña que se asocian a nodrizas podrían aumentar su rango de distribución altitudinal, permitiendo, en este caso, que las comunidades de alta montaña presenten una mayor riqueza que en ausencia de ellas (Cavieres et al. 1998; Núñez et al. 1999; Molina-Montenegro et al. 2000; Badano et al. 2002).
Las modificaciones microclimáticas producidas por especies en cojín han sido descritas con anterioridad por diversos autores para comunidades de alta montaña, destacándose la protección contra la desecación, las temperaturas extremas y el estrés hídrico (Hager & Faggi 1990; Cavieres et al. 1998; Núñez et al. 1999). La mayor humedad en el suelo bajo los cojines, en oposición al suelo desnudo, podría proporcionar una extensión del período de crecimiento durante los meses de mayor sequía (marzo-abril), permitiendo que las plantas que crecen entre los cojines mantengan su biomasa fotosintética por más tiempo. Además, las plantas que generan microhábitats de mayor humedad, permiten a las especies facilitadas mantener sus estomas abiertos durante más horas por día, mejorando sus balances de carbono (Holmgren et al. 1997). Este ambiente de mayor humedad que ofrecen los cojines, podría otorgar a las especies perennes que crecen entre ellos (e.g. Cerastium arvense, Hordeum comosum, Taraxacum officinale) más tiempo para la producción de compuestos de reserva, lo que podría mejorar su desempeño en el siguiente pulso reproductivo.
La mayor temperatura sobre plantas en cojín con respecto a los espacios abiertos ha sido descrita con anterioridad (Körner 1999). Sin embargo, nuestros resultados demuestran que el efecto nodriza por parte de las plantas en cojín puede darse a través de una disminución de las temperaturas máximas. Esto sugiere un efecto ëbufferí por parte de los cojines, ya que no sólo mantienen mayor temperatura que los espacios abiertos sino que permanecen mas fríos a las horas de máxima radiación (Figura 1). Este fenómeno ha sido principalmente atribuido a la arquitectura compacta y achaparrada que poseen estas formas de vida (Alliende & Hoffmann 1985). Por lo tanto, los cojines al poseer mayor temperatura a las horas más frías y menor temperatura a las horas de máxima radiación, provocarían una disminución del estrés en las plantas que crecen sobre ellos.
La tasa de fotosíntesis es un reflejo del estado fisiológico de una planta (Lambers et al. 1998), pudiendo verse reducida por las condiciones estresantes del ambiente. La mayor tasa de fotosíntesis de T. officinale entre los cojines, sugiere que en este micrositio disminuyen las condiciones estresantes del ambiente. Esto podría determinar una mayor captación de carbono, lo que estaría en concordancia con la mayor apertura estomática por parte de los individuos de T. officinale presentes en los cojines. En cambio, la tasa fotosintética y la conductancia estomática en E. collina fueron similares en los dos micrositios. Esto podría deberse a que, si bien los cojines presentan temperaturas y contenido hídrico menos estresantes, el efecto de crecer entre ellos disminuiría la captacion de recursos de esta especie (e.g. carbono) como respuesta a la competencia entre la especie facilitada y la nodriza (Cavieres et al. 1998).
En correspondencia con las mayores tasas fotosintéticas y de conductancia estomática y el marginal aumento del PWUE de T. officinale creciendo sobre L. acaulis, esta especie tiende a distribuirse preferentemente sobre estos cojines donde la disponibilidad de agua es mayor y la temperatura durante las horas de máxima radiación es menor que en los espacios abiertos. Para el caso de E. collina, no hubo una correspondencia entre la tasa fotosintética, la conductancia estomática, la PWUE y su distribución espacial. Esta especie se distribuye preferentemente fuera de los cojines, lo cual indicaría que la competencia generada con la especie nodriza por otros recursos diferentes que el agua, podría limitar su crecimiento entre los cojines.
Recientes experimentos de trampas de semillas sobre cojines y espacios aledaños, evidenciaron que el número de semillas de T. officinale y de E. collina no fue diferente entre los micrositios (Molina-Montenegro, datos no publ.), sugiriendo que la distribución espacial de estas especies no estaría determinada en la etapa de dispersión. Sin embargo, sería el reclutamiento de individuos la etapa que podría determinar la distribución no equitativa entre los micrositios, donde asociarse a una especie que reduzca las condiciones limitantes podría aumentar la supervivencia para una especie determinada o extender el tiempo de permanencia del tejido aéreo (Holmgren et al. 1997).
Sin embargo, una explicación alternativa por el cual existe un mayor número de individuos fuera de los cojines estaría dado por el ´óptimo´ de las condiciones ambientales. Para E. collina, no serían limitantes las temperaturas y las concentraciones de agua en los espacios abiertos y por lo tanto, establecerse fuera evitaría el efecto de competir con una especie sucesionalmente más temprana como lo es L. acaulis. La distribución no equitativa del número de individuos de cada especie entre y fuera de los cojines, y la variación en sus tasas de fotosíntesis, podrían sugerir que los individuos entre los cojines presentan un balance de carbono más positivo (Holmgren et al. 1997). Esto, podría traducirse en que entre los cojines se registre un mayor número de individuos creciendo y por un periodo de tiempo mayor. Sin embargo, dependiendo de si el efecto de asociarse es más o menos beneficioso que el crecer solo, cada especie podría aumentar la frecuencia de establecimiento en uno de los dos micrositios o evidenciar un mejor desempeño fisiológico.
AGRADECIMIENTOS
Los autores agradecen a E. Gianoli, C. Torres y B. Morales por sus valiosos comentarios y sugerencias sobre el manuscrito. Tambien agradecemos los valiosos comentarios de los evaluadores durante la etapa de revision. A C. Lusk por el software Resampling Stats. M.A.MM. agradece a la beca doctoral CONICYT. E.I. Badano agradece a MECESUP UCO 9906. Este trabajo fue financiado por el proyecto Fondecyt 1030821. Este articulo forma parte de las actividades de investigación del Centro Milenio para Estudios Avanzados en Ecología y Biodiversidad No. P02-051-F ICM.
REFERENCIAS
Alliende, MC & AJ Hoffmann. 1985. Plants intruding Laretia acaulis (Umbelliferae), a high Andean cushions plant. Vegetatio 60:151-156. [ Links ]
Badano, EI; MA Molina-Montenegro; C Quiroz & LA Cavieres. 2002. Efectos de Oreopolus glacialis (Rubiáceae) sobre la riqueza y diversidad de especies de plantas en una comunidad altoandina. Rev. Chil. Hist. Nat. 75:757-765. [ Links ]
Bertness, M & S Yeh. 1994. Cooperative and competitive interactions in the recruiment of marsh elders. Ecology 75:2416-2429. [ Links ]
Billings, WD. 1972. Arctic and alpine vegetation: plants adaptations to cold summer climates. Pp. 403-443 en: JD Ives & RG Barry (eds). Arctic and Alpine Environments. Methuen. London. [ Links ]
Callaghan, TV. 1987. Plant population processes in arctic and boreal regions. Ecol. Bull. 38:58-68. [ Links ]
Callaway, RM & S Pennings. 2000. Facilitation may buffer competitive effects: indirect and diffuse interactions among salt marsh plant. Am. Nat. 156:416-424. [ Links ]
Callaway, RM & FI Pugnaire. 1999. Facilitation in plant communities. Pp. 623-648 en: FI Pugnaire & F Valladares (eds). Handbook of Functional Plant Ecology. Marcel Dekker Press. Nueva York. [ Links ]
Cavieres, LA & MTK Arroyo. 1999. Tasa de enfriamiento adiabático del aire en el valle del río Molina, provincia de Santiago, Chile central (33ºS). Rev. Geográf. Chil. Terra Australis 44:79-86. [ Links ]
Cavieres, LA & MTK Arroyo. 2000. Altitudinal vegetation belts in the high-Andes of central Chile (33ºS). Rev. Chil. Hist. Nat. 73:331-334. [ Links ]
Cavieres, LA; MTK Arroyo; MA Molina-Montenegro; C Torres & A Peñaloza. 2002. Nurse effect of Bolax gummigera (Apiaceae) cushion plant in the alpine vegetation of the Chilean Patagonian Andes. J. Veg. Sci. 13:547-554. [ Links ]
Cavieres, LA; A Peñaloza; C Papic & M Tambutti. 1998. Efecto nodriza del cojín Laretia acaulis (Umbelliferae) en la zona alto-andina de Chile central. Rev. Chil. Hist. Nat. 71:337-347. [ Links ]
Chambers, JC; JA Macmahon & RW Brown. 1990. Alpine seedling establishment: the influence of disturbance type. Ecology 71:1323-1341. [ Links ]
di Castri, F & E Hajek. 1976. Bioclimatología de Chile. Ediciones de la Pontificia Universidad Católica de Chile. Santiago de Chile, Chile. 129 pp. [ Links ]
Hager, J & AM Faggi. 1990. Observaciones sobre distribución y microclima de cojines enanos de la isla de Creta y del noroeste de la Patagonia. Parodiana 6:109-127. [ Links ]
Hoffman, A; MTK Kalin-Arroyo; F Liberona; M Muñoz & J Watson. 1998. Plantas altoandinas. Imprenta Salesianos. Santiago de Chile, Chile. 280 pp. [ Links ]
Holmgren, M; M Scheffer & M Huston. 1997. The interplay of facilitation and competition in plant communities. Ecology 78:1966-1975. [ Links ]
Hunter, A & L Aarssen. 1988. Plants helping plants. BioScience 38:34-40. [ Links ]
Kikvidze, Z. 1996. Neighbour interaction and stability in subalpine meadow communities. J. Veg. Sci. 7:41-44. [ Links ]
Kikvidze, Z; L Khetsuriani; D Kikodze & RM Callaway. 2001. Facilitation and interference in subalpine meadows of the central Caucasus. J. Veg. Sci. 12:833-838. [ Links ]
Körner, CH. 1999. Alpine Plant Life. Springer-Verlag. Berlin. 338 pp. [ Links ]
Körner, CH & P Cochrane. 1983. Influence of plant physiognomy on leaf temperature on clear midsummer days in the Snowy Mountains, south-eastern Australia. Acta Oecologia, Oecol. Pl. 4:117-124. [ Links ]
Körner, CH & JAP De Moraes. 1979. Water potential and diffusion resistance in alpine cushion plants on clear summer days. Oecol. Pl. 14:109-120. [ Links ]
Lambers, H; F Chapin III & T Pons. 1998. Plant physiological ecology. Springer-Verlag. New York. 540 pp. [ Links ]
Loik, ME & KD Holl. 2001. Photosynthetic responses of tree seedlings in grass and under shrubs in early-successional tropical old fields, Costa Rica. Oecologia 127:40-50. [ Links ]
Molina-Montenegro, MA; C Torres; MJ Parra & LA Cavieres. 2000. Asociación de especies al cojín Azorella trifurcata (Apiaceae) en la zona andina de Chile central (37ºS). Gayana Bot. 57:161-168. [ Links ]
Nardini, A; MA Lo Gullo & S Salleo. 1999. Competitive strategies for water availability in two Mediterranean Quercus species. Plant Cell Envir. 22:109-116. [ Links ]
Niinements, U; W Bilger; O Kull & JD Tenhunen. 1999. Responses of foliar photosynthetic electron transport, pigment stoichiometry, and stomatal conductance to interacting environmental factors in a mixed species forest canopy. Tree Physiol. 19:839-852. [ Links ]
Núñez, C; M Aizen & C Ezcurra. 1999. Species associations and nurse plant effect in patches of high-Andean vegetation. J. Veg. Sci. 10:357-364. [ Links ]
Pugnaire, FI; P Haase & J Puigdefabregas. 1996. Facilitation between higher plant species in a semiarid environment. Ecology 77:1420-1426. [ Links ]
Pysek, P & J Lyska. 1991. Colonization of Sibbaldia tetrandra cushions on alpine scree in the PalmiroAlai mountains, Central Asia. Arct. Alp. Res. 23:263-272. [ Links ]
Resampling Stat. 1990-9995. Resampling Stat user´s guide. Resampling Stat, Inc., Arlington, VA, EE.UU. [ Links ]
Sakai, A & W Larcher. 1987. Frost survival of plants: responses and adaptation to freezing stress. Springer-Verlag. Berlin. 323 pp. [ Links ]
Slade, N & D Hall. 1999. Randomization test using standard statistical software. Bull. Ecol. Soc. Am. 80:179-180. [ Links ]
Smith, AP & TP Young. 1987. Tropical alpine plant ecology. Annu. Rev. Ecol. Syst. 22:177-191. [ Links ]

