










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.15 n.2 Córdoba jul./dic. 2005
TRABAJOS ORIGINALES
Culture of Eisenia fetida (Annelida, Lumbricidae) on puffed rice scrap in outdoors and laboratory conditions
Miguel Schuldt 1, 2,*, Alejandra Rumi 1, 2, Diego E Gutiérrez Gregoric 1, 2, Nicolás Caloni 3, Josefina Bodnar 2, Natalia Revora 2, Victoria Tasso 2, Mariano Valenti 2, Julio Varela 2 & Horacio De Belaustegui 3
1. División Zoología Invertebrados, Museo de La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina
2. Fac. de Ciencias Naturales y Museo, Universidad Nacional de La Plata, La Plata, Buenos Aires, Argentina
3. Departamento de Ciencias Básicas, Universidad Nacional de Luján, Luján, Argentina
*División Zoología Invertebrados, Museo de La Plata, Facultad de Ciencias Naturales y Museo, Univ. Nac. de La Plata, Paseo del Bosque s/nº, 1900, La Plata, Buenos Aires, Argentina. CONICET.
Email: miguel_schuldt@hotmail.com
Recibido: 16 de junio de 2004;
Fin de arbitraje: 1 de septiembre de 2004;
Revisión recibida: 8 de marzo de 2005;
Aceptado: 24 de octubre de 2005
ABSTRACT. Industrial puffed rice scrap increases the carbon/nitrogen ratio in carbon-poor biodegradable organic matter, such as broiler chicken beddings (C/N= 14.6) (droppings + sunflower husks), optimizing the composting process. In Eisenia fetida field experiences (bimonthly sampling) done on cultures managed through self-inoculation (addition of 8-16% scrap to the chicken bedding), the substrate was accepted. Population increase and cocoon production, measured 127 days after the start, were not significantly different from control plots (broiler chicken bedding). Population size expanded with a nominal (nd) and effective (ed) increase of 2.69 and 3.51 times respectively compared to the initial numbers. The carrying capacity of the different organic matter mixtures approximates 30000 worms/bed (2m²) (w/B). The reproductive potential ("R") showed fluctuations (between 1000 and 10800 w/B) which agree with the dynamics of a population under this management strategy. Humus production was approximately 35 tons from an original 63.9 tons. In laboratory cultures (weekly weight control) with pure scrap, as worm substratum-feed, the following results, compared to home waste compost and rabbit manure (control), were recorded: more sexually mature animals; higher cocoon-per-animal production; average weight (at 17 weeks) a 47% higher than the weight obtained when using other diets.
Keywords: Vermiculture; Growth rates; Reproductive potential.
RESUMEN. Cultivo de Eisenia fetida (Annelida, Lumbricidae) con scrap de arroz expandido en condiciones de temperie y laboratorio: El arroz expandido se utiliza en la fabricación de galletas para el consumo humano. Esta industria genera una gran cantidad de desperdicios biodegradables, principalmente un scrap con predominio de hidratos de carbono. Este desperdicio eleva la relación carbono/nitrógeno de la materia orgánica biodegradable con déficit de carbono. En el caso de las "camas" de pollos parrilleros (estiércol más cáscara de girasol, relación C/N= 14.6), el empleo de scrap optimiza su compostaje a una relación C/N= 40/1 – 30/1. En experiencias realizadas en el campo, con muestreos bimensuales, y en cultivos de Eisenia fetida con manejo mediante autosiembra (adición a las "camas" de pollo de 8-16% de scrap), resultó una buena aceptación del sustrato. Respecto del testigo ("cama" de pollos parrilleros), el incremento de la población de E. fetida al cabo de 127 días no se diferenció significativamente, al igual que en la producción de cocones. En laboratorio se realizaron cultivos con tres tipos de dieta para E. fetida con un control semanal del peso y con tres réplicas por dieta. La dieta 1 (testigo) se hizo en base a estiércol de conejo, la dieta 2 con compost de residuos domiciliarios y la 3 con scrap puro como medio-alimento de las lombrices. En los cultivos con scrap, con respecto a los otros dos, se observó que en 12 semanas presentaron más animales sexualmente maduros (dieta 1 = 16.7%; dieta 2 = 26.7%; dieta 3 = 91.3%). Independientemente de la dieta, las lombrices adquirieron la madurez con pesos corporales mayores a 0.24 g. Presentaron mayor producción de cocones/ animal (dieta 1 = 0.37; dieta 2 = 0.86; dieta 3 = 1.65) acorde con la elevación del potencial reproductor (dieta 1 = 2330; dieta 2 = 3729; dieta 3 = 12772). El peso medio en 17 semanas superó al de las restantes dietas en un 47% (dieta 1 = 0.178 g, DS = 0.0751, n = 27; Dieta 2 = 0.265 g, DS = 0.0738, n = 30; dieta 3 = 0.567 g, DS = 0.1276, n = 23).
Palabras clave: Lombricultura; Tasas de crecimiento; Potencial reproductivo.
INTRODUCTION
Puffed rice is used in the making of rice-cakes for human consumption. This industry generates large amounts of biodegradable wastes, especially carbohydrate-rich scrap. For subsegments such as broiler chicken bedding (C/N=14.6) (droppings + sunflower husks), with less carbon than the optimal ratio for composting (C/N= 40/1 – 26/1), the addition of industrial puffed rice scrap rises the carbon/nitrogen ratio of this biodegradable organic matter (Cormillot 1998).
The goals of this work are an assessment of the aptitude of this scrap as culture media (substratum) for Eisenia fetida (Savigny 1826) to transform into humus (vermicompost), and simultaneously, an analysis of the evolution of the earthworm population structure after colonization of the substratum; an evaluation of its carrying capacity and its incidence on worm growth measured by weight. We also aim to identify the transition of worms to maturity and establish their reproductive potential. To achieve these goals, two vermicultures were established, one in the field (outdoors) and another one under laboratory conditions.
METHODS
Vermicultures: Monitoring
a) Field (outdoors): On 20th September 1999 six 16 x 3 x 0.5 m sectors, dome-shaped and without lateral boundaries, were located in Luján (34º34´S; 59º06´W), Buenos Aires province, Argentina. The total volume for the six sectors (144 m³) was graphically corrected according to the convexity of each pile. It included a total volume estimated of 128 m³ of wastes (beds) from poultry broiler chicken production, comprising sunflower husks and droppings (Fig. 1). By then, the organic matter (OM) had already been stored for 3 - 4 months. On 6th November 1999, 30 and 60 bags (60 dm³ each) of puffed rice wastes were added to sectors 1 and 2 respectively. Drop irrigation in these and the other two neighboring sectors kept moistness at 70 - 90%. Forty-five days after the addition of scrap to the "beds", at temperatures < 40 ºC, the composting sectors were inoculated with E. fetida. The following parameters were considered:
• Nominal density (nd) = nº worms/surface area or volume. For a nominal fecundity = nº worms/cocoon = 1 (conventional figures).
• Effective density (ed) = nº worms/surface area or volume. For an effective fecundity (worms/cocoon) = 2.69 (Schuldt obs. pers.).
• Beds (B) = 2 m².
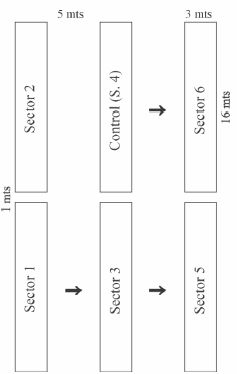
Figure 1. Schematic diagram of the arrangement of field culture sectors. The arrows indicate the escape way.
Figura 1. Diagrama esquemático de los sectores de cultivo en el campo. Las flechas indican la dirección de fuga.
The vermicomposting process started with a "nd" at inoculation = 14615 worms/Bed (w/B). The three inoculated sectors were sampled on a bimonthly basis, as well as the three noninoculated sectors that were afterwards invaded by the worms (self-inoculation, emigration). The substratum samples were obtained using a 7.5 x 7.5 cm cubical extractor (sampling unit (s.u.) = 421.87 cm³ ). Sample size was 2 to 4 s.u. (Schuldt et al. 1998).
Four basic population segments were considered (Schuldt et al. 1998): 1) fertile (filled) cocoons (C1); 2) juvenile individuals, translucent,< 1.5 cm long (C2); 3) subadult individuals, pigmented, without visible clitellum, frequently < 3 cm long (C3) (all pigmented individuals lacking a clitellum, even those that were > 3 cm long were included in C3); and 4) adult worms (with visible clitellum) (C4) (Schuldt et al. 1999). In addition, the presence of empty cocoons (Co) was considered and their numbers counted, as it gives information about the magnitude of the reproductive event. The differences between samples from each sector/date and between different population segments, as well as the presence of empty cocoons, were contrasted using two-factor ANOVA. A "t" test for differences between means was used to identify the sectors with the highest differences between population segments.
b) Laboratory
The culture was started on 12th August 2000 in plastic Petri dishes (9 cm diameter). Six weeks later, the worms were moved into larger plastic dishes (13 x 9 x 4 cm) where they were kept until 19th December 2000. Three diets were used: 1) fresh rabbit manure (control); 2) home wastes compost from Villa Elisa, comprising mainly vegetables and fruits; 3) pure whole grain rice scrap. For each treatment, three replicates were established using 10 worms/replicate. Live weight was measured weekly using digital scales (0.0001 gr precision). The feed was added once a week, assuming that consumption was equal to the weight of the animals. Temperatures fluctuated between 10ºC and 25ºC. Moistness was kept at 70 - 90%. The worms were handled using hand-made steelwire loops.
Prior to the application of the growth model by weight, the differences between replicates for each treatment were assessed using ANOVA (P = 0.05%) (one-factor analysis). The individuals were randomly sampled (weight range for newly-hatched worms = 0.007 - 0.03 gr). For the purpose of assessing the growth (measured as weight increase) of the animals for each diet, the observed mean weights by date were adjusted to a growth curve derived from the logistic model (Ricklefs 1967):
W = Wmax / 1+be-kt
where W: weight expressed as decimal fraction of its asyntotic weight; b = 1 (b: constant, the inflection point occurs at t = 0 when b = 1); k: a constant proportional to the growth rate; Wmax: maximum weight, estimated from the mean weight values, and t: time expressed in weeks. The adjustment of the model was evaluated using the "![]() " test for the comparison of observed and estimated weight values.
" test for the comparison of observed and estimated weight values.
c) Fecundity. Reproductive potential.
The calculation of the effective fecundity of E. fetida (2.69 embryos/cocoon) made it possible to distinguish between nominal (nd) and effective size (ed) in the evolution of the global vermiculture population and its sectors. The "ed" of the samples tends to be a better approximation of the estimated actual number of animals in the population (Figs. 1, 2, 3). The reproductive potential for the circumstances (field conditions) was calculated as: "R" = x (bnp) (x: worms of the population in a given situation, in this case x = 100; b: mean number of embryos-worms/cocoon; n: mean number of clutches/year; p: the ratio of adults and subadults) (Wildish 1979, 1980; Schuldt obs. pers.). In the calculation of "R" for laboratory conditions, the term "p" is calculated from the consideration of the individuals with incipient clitellum and conspicuous clitellum only, distinguishing these from the subadults (immature animals).
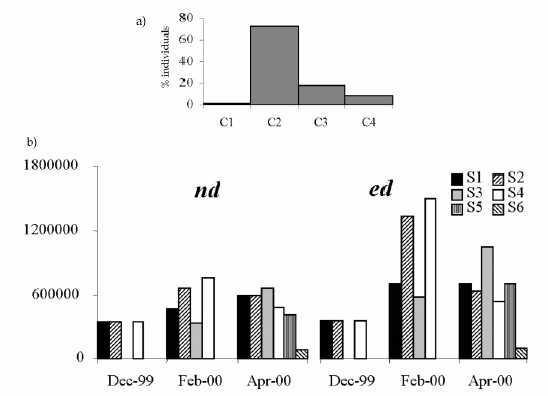
Figure 2. a: Population structure of Eisenia fetida before the inoculation (overcrowding). Abscises: population segments. b: Population evolution (nd and ed) in the different culture sectors (S1 to S6). Abscises: sample date; Ordinates: nº of worms/sector.
Figura 2. a: Estructura poblacional de Eisenia fetida antes de la inoculación (apiÒamiento). Abscisas: estamentos poblacionales. b: Evolución poblacional (nd y ed) en los diferentes sectores de cultivo (S1 a S6). Abscisas: fecha de muestreo; Ordenadas: nº de lombrices/sector.

Figure 3. Population structure at each sector on each sample date. a: February; b: April. Abscises: population segments; Ordinates: nº of worms.
Figura 3. Estructura poblacional en cada sector en cada fecha de muestreo. a: Febrero; b: Abril. Abscisas: estamentos poblacionales; Ordenadas: nº de lombrices.
RESULTS
Vermicultures: Monitoring.
a) Field (outdoors): The initial population structure at insemination (Fig. 2a) had a predominance of juvenile (C2) individuals, as is commonly observed in expanding populations that are not yet balanced with respect to the carrying capacity of the substratum. This reflects the management based on trimestral subdivisions of the population and periodical (weekly) feeding. Figure 3 shows the evolution of age structure in the different sectors. Population size increased by a nominal (nd) and effective (ed) growth of 2.69 and 3.51 times the initial value respectively. The carrying capacity for the different organic matter mixtures approximates 30000 w/B. The reproductive potential ("R") showed fluctuations (between 1000 and 10800) according to the dynamics of a population managed by self-inoculation.
The ulterior development of the culture showed the typical events of self-propagation of the earthworms once the substratum/feed is spent. After reaching densities of about 30000 worms/bed, and before 60 days had elapsed since the start of the culture, emigration of the worms and colonization of sector 3 (Figs. 1, 3) was observed. The bimonthly rhythm for the colonization of new sectors (S5 and S6) was maintained afterwards.
After 127 days, sectors 1, 2 and 4, which had been inoculated on December 2000, still held significant residual populations, with nominal densities of between 20000 and 25000 worms/bed. Humus production was approximately 35 tons, from the processing of 128 m³ of egests and scrap. Earthworms to be used in other projects were also obtained.
ANOVA analysis:
a-1) Worm densities/sector/date were contrasted, and the differences between simples/sector were not significant. In contrast, there were differences between the population segments of the samples from different sectors (Table 1).
Table 1. Results of ANOVA on worm densities by sector and by date for outdoors/field culture (P < 0.05). *: differs significant; **: differs highly significant.
Tabla 1. Resultado del test de ANOVA sobre densidades de lombrices por sector y por fecha para el cultivo en temperie
(P < 0.05). *: diferencia significativa; **: diferencia altamente significativa.

a-2) On the basis of the (a-1) results, an analysis was performed to identify the segments with statistically significant differences (Table 2).
Table 2. ANOVA for the segments (P < 0.05). *: differs significant; **: differs highly significant.
Tabla 2. ANOVA sobre los estamentos (P < 0.05). *: diferencia significativa; **: diferencia altamente significativa.

a-3) The mean individuals/stratum - Co/sector were also contrasted by dates. "Co" was the only parameter to show differences for the April sample (Table 3). On the basis of this result, the greatest variation was identified between sector 2 and the control plot (sector 4) (t = 6.39; P < 0.05; DF = 4).
Table 3. ANOVA for the mean nº of individuals per stratum and Co / sector / date (P < 0.05). *: differs significant.
Tabla 3. ANOVA sobre el promedio de individuos por estamentos por sector por fechas (P < 0.05). *: diferencia significativa.

b) Laboratory:
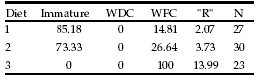
Evolution of maturity: On 02 November (week 10), the first individuals with indications of clitellum, which corresponded to diet 1 (3 out of 30) (average: 0.352 g; SE: 0.0188) and diet 3 (6 out of 30) (average: 0.4249 g; SE: 0.0807), were observed. None of the animals kept on diet 2 showed signs of maturity. In the case of the latter diet, only on 15 November (week 12) the first individuals with developing clitellum (3 out of 30) (average: 0.3161 g; SE: 0.0274) and with conspicuous clitellum (5 out of 30) appeared (Table 4). At 12 weeks, the cultures with added scrap had more sexually mature animals (diet 1 = 16.7%; diet 2 = 26.7%; diet 3 = 91.3%).
Table 4. Prematurity-maturity transition: clitellum development and reproductive potential ("R") (Diets 1-3 at week 12) (Relative values – percentages). WDC: With developing clitellum; WFC: With full clitellum.
Tabla 4. Transición premadurez-madurez: desarrollo clitelar y potencial reproductor (ìRî) (Dietas 1-3 a la semana 12) (valores relativos - porcentajes). WDC: Con clitelo en vías de desarrollo; WFC: Con clitelo formado.

Table 4 shows the global culture conditions with respect to sexual maturity for the three diets; an increase in reproductive potential from diet 1 to diet 3 is observed. The trends observed here persisted to the end of the experiment (week 17), as seen in Table 5. In some individuals the development of clitellum was observed from 0.248 g, independently of the supplied diet. The greatest production of cocoons/ animal was recorded in scrap cultures (diet 1 = 0.37; diet 2 = 0.86; diet 3 = 1.65), which is in accordance with the rise in reproductive potential (diet 1 = 2330; diet 2 = 3729; diet 3 = 12772). The average weight at 17 weeks on pure scrap was higher than in the other diets by a 47% (diet 1 = 0.178g, SE = 0.0751, n = 27; diet 2 = 0.265 g, SE = 0.0738, n = 30; diet 3 = 0.567g, SE = 0.1276, n = 23).
Tabla 5. Prematurity ñ maturity transition: clitellum development and reproductive potential ("R") (Diets 1-3 at week 17) (Relative values – percentages). WDC: With developing clitellum; WFC: With full clitellum.
Tabla 5. Transición premadurez-madurez: desarrollo clitelar y potencial reproductor ("R") (Dietas 1-3 a la semana 17) (valores relativos – porcentajes). WDC: Con clitelo en vías de desarrollo; WFC: Con clitelo formado.
Growth rates: Figure 4 illustrates the Fisher values obtained from the ANOVA for each treatment and date, which indicate an initial dispersion in the observed individual weights. These differences tend to disappear in the course of the experiment according to the diet. The application of logistic growth curves granted an adequate description of the development of the animalsí weight for each diet, since ![]() showed good fit. The curves obtained are shown in figure 5 and the values for each of them are given in table 6. From the observation of the results of an ANOVA between diets (DF: 2 -48; Fisher: 6.17; P < 0.05) and figures 5 and 6, significant differences between growth rates are seen; the growth observed in specimens kept on rice scrap is higher following a trend that becomes evident from the fifth week.
showed good fit. The curves obtained are shown in figure 5 and the values for each of them are given in table 6. From the observation of the results of an ANOVA between diets (DF: 2 -48; Fisher: 6.17; P < 0.05) and figures 5 and 6, significant differences between growth rates are seen; the growth observed in specimens kept on rice scrap is higher following a trend that becomes evident from the fifth week.

Figure 4. ANOVA results for each diet and date. Ordinates: Fisher values. The straight line indicates the reference level at á = 0.05; n = 30; F = 3.543. Empty circles indicates n < 30.
Figura 4. Resultados del ANOVA para cada dieta y fecha. Ordenadas: valores Fisher. Línea recta indica el nivel de la referencia a á = 0.05; n = 30; F = 3.543. Los círculos vacíos indican n < 30.

Figure 5. Growth curves for the three diets. Dots/ Points: Observed weight; -: Standard Deviation; Line: Theoretical growth curve. Abscises: Weeks; Ordinates: weight in grams.
Figura 5. Curvas de crecimiento para las tres dietas. Puntos: peso observado; -: desviación estándar;
Línea,: Curva de crecimiento teórica. Abscisas: Semanas; Ordenadas: peso en gramos.
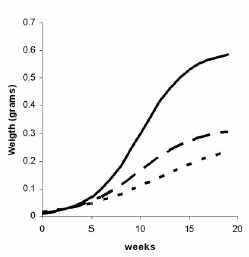
Figure 6. Growth curves expressed in weight for the three diets. Continuous line: Diet 3; Interrupted line: Diet 2; Dotted line:
Diet 1.
Figura 6. Curva de crecimiento en peso para las tres dietas. Línea continua: Dieta 3; Línea entrecortada: Dieta 2; Línea punteada: Dieta 1.
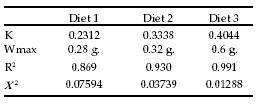
Table 6. Parameters and constants for the growth model calculated for the three types of diets.
Tabla 6. Parámetros y constantes del modelo de crecimiento halladas en las tres tipos de dietas.
DISCUSSION
In field conditions, no significant differences in density were observed between the different sectors and segments, despite the varied conditions of the culture medium for each sector. The sampling strategy did not allow us to track the destination of the escapes from the monitored sectors, which somewhat obscured the origin of the populations found in the noninoculated sectors (sectors 3, 5 and 6). However, in the case of sector 3, the animals were presumed to have originated in sector 1. This was inferred from the presence of subadults and adults in sector 3, which, when considered together with the ones in sector 1, added up to an amount similar to the adults in sectors 2 and 4 (control) (Fig. 3). The presence of juveniles in sector 3 indicates that the escapes from sector 1 had started approximately 20 days earlier. Sector 4, which held 31800 w/B, was considered unaffected by escapes; this assumption is backed up by the absence of individuals in sector 6. Sectors 5 and 6 had animals during the April 2000 sampling, presumably derived from sectors 3 and 4 respectively.
The population structure and the "R" parameter (Table 7), show that colonization of sector 6 was the most recent event. The notoriously lower "R" for the animals in sector 5 indicates an early colonization, which corresponds to the evolution of sector 3 (the first one to be colonized, February/2000). In the case of E. fetida, invasive populations have high "R", often above 5000 w/B. The residual populations show an initial decrease in "R" values due to the reduction of adult and subadult specimens, and afterwards they tend towards an increase of "R". This is related to the drastic reduction of copulation in impoverished subsegments (Schuldt 2001). The residual populations in sectors 1, 2 and 4 were significant in April 2000, with nominal densities between 20000 and 25000 w/B. This amounts to more than a 65% (up to 83%) of the carrying capacity of the substratum before the start of the vermicomposting procedure.
Table 7. Reproductive potential ("R") for the worms in each culture sector (Sn) on each sampling date. * Colonized by escapes.
Tabla 7. Potencial Reproductor ("R") de las lombrices en cada sector de cultivo (Sn) en cada fecha muestreada. * Colonizado por fugas.

Managing experiences of self-inoculation cultures using cow manure and approximately 30% fiber have shown that the densities of residual populations at 60 days post-escape are usually below half of the carrying capacity of the substratum. The higher relative carrying capacity of broiler chicken droppings might be due to the presence of sunflower husk fibers which have lower destabilization rates and offer support to the microflora and microfauna that in turn make up the substratum/food for the earthworms. This tends to compensate for the lower carrying capacity of bird droppings with respect to cow manure. The amount of food provided for the worms using broiler chicken beds was estimated as adequate for 90 days on the basis of previous experiences using cow egests, but it actually only sufficed for 60 days.
In general, the 55 days culture grew to 3.19 (de) times the initial population (from 1069680 earthworms to 4164244 earthworms). Afterwards it became stabilized, primarily due to the emigration of adult and subadult animals, coinciding with a carrying capacity of the substratum of approx. 30000 w/B. At the end of 127 days (since the start of the culture), the population increase with respect to the control ("broiler chicken bedding"). Cocoon production was not significantly different.
The higher growth and early attainment of sexual maturity observed in scrap-fed animals (diet 1) under lab conditions, contrast markedly with the lack of effects observed in field-culture populations, where neither the population size nor the cocoon (C1) segment were affected by the diet. As empty cocoons (Co) persist for more than six months in the substratum, they are good estimators of total reproductive performance. The differences between sectors with respect to this parameter appeared not to be directly related to scrap concentration. Pure scrap was used in lab cultures, whereas in field sectors the scrap proportion was below 16%, so that this lack of measurable effects could be due to insufficient concentrations.
The choice of pure rabbit manure as control diet is based on the existence of reliable information on the growth of E. fetida using this food (Herrera & de Mischis 1994). Besides, it was one of the diets tested by Di Masso et al. (1997), who compared the growth of earthworms fed separately on "coypo" ("rodent otter") (Myocastor coypus (Molina, 1782)), rabbit, cow and horse manure. He observed, after 20 weeks, the highest growths in the worms fed with "coipo" egests, followed by rabbit egests, whereas the growth on other media was slower. In this respect, the fact that both rice scrap and composted home wastes allow greater weight increase of the animals, as well as earlier attainment of sexual maturity, is an interesting corroboration.
The comparison of field and lab results shows that, even though the rice scrap concentrations used in field cultures elevated the carbon-nitrogen ratio with respect to the control, they were not sufficiently high (8 and 16%) to achieve an increase as significant as the result obtained in the lab using a pure scrap diet as culture medium.
CONCLUSIONS
On the basis of growth rates and cocoon production, laboratory experiments have shown that pure rice scrap is an excellent diet. Nevertheless, its effects become diluted in the case of the mixtures used for field tests. In the case of cultures managed by self-inoculation the conclusions are:
a) On the addition of 8-16% scrap to chicken "beds", the substratum was accepted.
b) The population increase at 127 days was not significantly different in any of the sectors; neither was the cocoon production. Population size increased with a nominal (nd) and effective (ed) growth of 2.69 and 3.51 times the initial size respectively.
c) The carrying capacity for the diverse organic matter mixtures approximates 30000 w/B.
d) The reproductive potential ("R") oscillated between 1000 and 10800 w/B.
e) Total humus production was approximately 35 tons, from an original 63,9 tons of organic matter (128 m³), after a 127-day processing period.
f) By the end of the experiment, sectors 1, 2 and 4 still held important residual populations, with nominal densities of 20000 to 25000 w/B, in contrast with other diets (cow manure).
In laboratory cultures using pure scrap as medium-feed for the worms (diet 3), the comparison with home wastes compost (diet 2) and rabbit manure (diet 1 -check/control) showed the following:
a) More sexually mature animals at 12 weeks (diet 1 = 16.7%; diet 2 = 26.7%; diet 3 = 91.3%).
Regardless of the diet used, the earthworms reached sexual maturity at body weight > 0.24 g.
b) Higher cocoon per worm production (diet 1 = 0.37; diet 2 = 0.86; diet 3 = 1.65), parallel with the increase in reproductive potential (diet 1 = 2330; diet 2 = 3729; diet 3 = 12772).
c) Average weight at 17 weeks was higher than weight for other diets by a 47% (diet 1 = 0.178 g, SE = 0.0751, n = 27; diet 2 = 0.265 g, SE = 0.0738, n = 30; diet 3 = 0.567 g, SE = 0.1276, n = 23).
ACKNOWLEDGEMENTS
Field activities were performed as part of an Agreement for Technological Transference between the Enterprise Hatchery of Universidad Nacional de Luján (UNLu) and CEREALKO S. A., which provided partial funding for these activities. We also thank the Technology Department at UNLu for permission to use the experimental field and its equipment.
REFERENCES
Cormillot, A. 1998. Qué comes cuando comes. Tablas de calorías y nutrientes de todos los alimentos para armar una dieta sana. Editorial Perfil. Buenos Aires. [ Links ]
Di Masso, RJ; LB Marc & NR Biasatti. 1997. Earthworm Eisenia foetida (Savigny) growth in Coypu and other animal faeces as nutritional subsegments. Megadrilogica 6(12):105-112. [ Links ]
Herrera, JAD & C DE Mischis. 1994. Influence of feeding in the biological cycle of Eisenia foetida (Savigny) (Annelida, Oligochaeta, Lumbricidae). Part I. Megadrilogica 5(11):117-124. [ Links ]
Schuldt, M. 2001. Lombricultura. Su teoría y práctica en el ámbito agropecuario, industrial y doméstico. Ed. IMPRELyF. La Plata. [ Links ]
Schuldt, M; A Rumi; L Guarrera & PH De Belaustegui. 1998. Programación de muestreos de Eisenia foetida (Annelida, Lumbricidae). Adecuación a diferentes alternativas de manejo. Rev. Arg. Prod. Animal 18(1):53-66. [ Links ]
Schuldt, M; A Rumi; L Guarrera; HP De Belaustegui & JP Mauro. 1999. Producción de lombricompuesto a partir de egestas de feedlot: Población de lombrices y gestión de residuos. Rev. Arg. Prod. Animal 19(2):331-346. [ Links ]
Wildish, DJ. 1979. Reproductive consequences of the terrestrial habit in Orchestia (Crustacea: Amphipoda). Internat. J. Invertebr. Reprod. 1:9- 20. [ Links ]
Wildish, DJ. 1980. Reproductive bionomics of two sublittoral amphipods in a Bay of Fundy estuary. Internat. J. Invertebr. Reprod. 2:311-320. [ Links ]

