










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.17 no.1 Córdoba Jan./June 2007
SECCIÓN ESPECIAL
Robustness of plant-flower visitor webs to simulated climate change
Mariano Devoto1,*, Martín Zimmermann2 & Diego Medan1
1. Cátedra de Botánica, Facultad de Agronomía de la Universidad de Buenos Aires, Buenos Aires, Argentina.
2. Depto. de Física, Fac. de Ciencias Exactas y Naturales, Univ. de Buenos Aires, Buenos Aires, Argentina.
* Cátedra de Botánica, Facultad de Agronomía de la Universidad de Buenos Aires, Avenida San Martín 4453 (C1417DSE) Buenos Aires, Argentina.
Present address: School of Biological Sciences, University of Bristol, Woodland Road, BS8 1UG, Bristol, UK. Phone: 00441179545960. Email: mdevoto@agro.uba.ar
Recibido: 11 de enero de 2007;
Fin de arbitraje: 20 de febrero de 2007;
Revisión recibida: 27 de marzo de 2007;
Aceptado: 20 de abril de 2007
ABSTRACT. Climate change threatens the integrity of pollination webs by decoupling plants and flower visitors both phenologically and ecologically. We simulated a range shift of flower-visitor species along a steep east-west rainfall gradient to explore resulting patterns of extinction. The systems studied seemed to be rather robust to range shifts. This results from (a) the positive correlation between the geographic range and both the local (i.e. at each site) and the regional (i.e. pooling across sites) number of interaction partners of the plants and flower visitors of these webs, and (b) a nested assembly of interactions at the regional level. In a broader context, this study reveals itself as a useful tool for understanding the links between the interaction pattern of mutualistic webs and their response to a major threat such as climate change.
Keywords: Mutualism disruption; Plant-pollinator interaction; Patagonia; Global change; Extinction; Nestedness.
RESUMEN. Robustez de las redes planta-visitante floral en simulaciones de cambio climático: El cambio climático amenaza la integridad de las redes de polinización al desacoplar la plantas de sus polinizadores tanto fenológica como ecológicamente. En este trabajo simulamos un desplazamiento de los rangos geográficos de las especies de visitantes florales a lo largo de un abrupto gradiente de precipitación este-oeste y exploramos los patrones de extinción resultantes. Los sistemas estudiados resultaron moderadamente robustos ante los desplazamientos de distribución generados. Esto se puede explicar por (a) la correlación positiva entre el número de sitios a lo largo del gradiente en que fue encontrada una especie (de planta o de visitante floral) y su número de especies mutualistas tanto a nivel local (en cada sitio) como regional (agregando entre sitios), (b) el patrón de ensamble anidado que muestran las interacciones a nivel regional. En un contexto más general, este estudio resulta una herramienta útil para comprender la relación entre la estructura de interacciones de una red mutualista y su respuesta ante la amenaza del cambio climático.
Palabras clave: Disrupción de mutualismos; Interacción planta-polinizador; Patagonia; Cambio global; Extinción; Anidamiento.
INTRODUCTION
The ecology of plant-pollinator (p-p) webs has received much attention recently, with the main efforts concentrated in describing general patterns of assembly and interaction between species (Memmott 1999; Vázquez & Aizen 2003; 2004, Jordano et al. 2003; Vázquez 2005; Jordano et al. 2006), the influence of environmental and geographical variables on these patterns (Arroyo et al. 1982; Totland 1993; Elberling & Olesen 1999; Malo & Baonza 2002; Medan et al. 2002; Ollerton & Cranmer 2002) and the response of these systems to human-related perturbations (multiple factors, Bond 1994; alien plants introduction, Memmott & Waser 2002, Morales & Aizen 2002; cattle grazing, Vázquez & Simberloff 2003, 2004; fragmentation, Aizen & Feinsinger 1994ab; Aizen et al. 2002; Ashworth et al. 2003).
One potentially serious threat to p-p mutualisms is climate change, which has been predicted to decouple plants and flower visitors phenologically and ecologically (Kearns et al. 1998; McCarthy 2001; Peñuelas et al. 2004), and to alter patterns of competition for flower visitors among plants (Thórhallsdóttir 1998). There is evidence of altitudinal (upward) and latitudinal (poleward) range shifts and phenological shifts of major groups from arthropods to mammals and from grasses to trees (Hughes 2000; Davis & Shaw 2001; McCarthy 2001; Watt & MacFarlane 2002; Walther et al. 2002; Dixon 2003; Root et al. 2003), and experimental evidence of temporal shifts in plant phenology in response to artificial warming (Price & Waser 1998) and artificial modification of water availability (Peñuelas et al. 2004). However, to our knowledge there is still no evidence of disruption of pollination mutualisms as a consequence of climate change.
In this regard, modeling techniques can help to envision the consequences that range shifts of plants and flower visitors might have on pollination mutualisms. The main obstacle is the lack of a proper data set consisting of several p-p webs in close proximity and along an environmental gradient likely to be affected by directional climate change (e.g. gradual change in temperature and precipitation patterns). Naturally occurring gradients, where variation in a single environmental factor outweighs other less variable factors, are a useful ecological tool in global change research (Vitousek & Matson 1991; Steffen et al. 1999) as they can help predict how environmental factors influence the structure and functioning of terrestrial ecosystems. An initiative of the International Geosphere-Biosphere Program (IGBP) identified on all continents a set of naturally occurring transects of climate and vegetation which represent an array of regional-scale gradients that vary in major environmental variables (Steffen et al. 1999).
In the Patagonian region of South America, from 40° to 55° S, there is a strong longitudinal rainfall gradient, with a shift in vegetation in less than 150 km from xeric desert shrubland to grass-shrub steppe, leading to a low stature tree cover and finally closed canopy forest (Movia et al. 1982; Austin & Sala 2002). This rainfall gradient, identified as an IGBP transect, provided us with the opportunity to study a wide range of p-p webs occurring in close proximity, encompassing a striking change in mean annual rainfall but a minor 0.5-fold change in altitude (Devoto et al. 2005) and a 0.25-fold change in mean annual temperature (Movia et al. 1982).
Most of the existing climate change models predict an increase in mean annual temperature coupled with a decrease in mean annual precipitation in NW Patagonia for the next century even under optimistic scenarios of atmospheric CO2 increase (see review by Hulme & Sheard 1999). This change in local climate conditions would be evidenced by a westward movement of the isohyets. As mentioned above such shift is likely to affect the distribution of plants and flower visitors within the region and possibly to decouple them spatially. In this paper, we simulate the extinction dynamics of species in plant-flower visitor webs in response to range shifts of flower visitor species and consequent decoupling of pollination mutualisms as expected under climate change. We used data from eight real plant-flower visitor webs distributed along a steep gradient of mean annual rainfall in NW Patagonia, Argentina. Given the strong compositional change in an east-west direction reported between these plant-flower visitor webs (Devoto et al. 2005), we expected that, in general, a range shift of flower visitors would cause major disruptions of pollination interactions and consequently large extinction events both among plants and flower visitors. We also expected that simulating migrations of increasing proportions of insects would result in clearly larger proportions of species becoming extinct. We specifically seek to answer the following questions: (1) How vulnerable are plant-flower visitor webs to a simulated westward shift in the range of the flower visitors? (2) Is there any property of p-p webs associated with robustness to such a shift?
MATERIALS AND METHODS
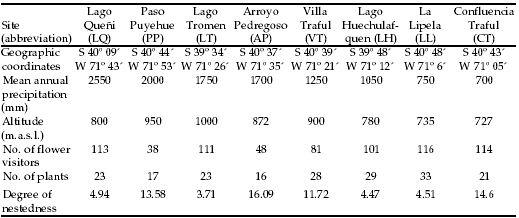
Eight plant-flower visitor webs were constructed with field data gathered from 1997 to 2002 along a steep rainfall gradient on the western part of the IGBP transect mentioned above (Table 1). The study sites comprise a mean annual precipitation from 700 to 2550 mm and range from a xeric grass-shrub steppe to a humid temperate forest (Zegers 1993; Arroyo et al. 1996; Paruelo et al. 1998ab). It should be noted, however, that the rainfall gradient studied here is part of a more extended gradient which extends from ca. 200 mm in central Patagonia to more than 3000 mm on the Chilean slope of the Andes. Extrapolating from temperature charts (Movia et al. 1982), there is a decrease in mean annual temperature (MAT) from CT (ca. 10ºC) to LQ (ca. 8ºC) that is roughly associated to differences in altitude (Table 1). While this small variation may explain some of the observed differences in assemblage composition, the turnover of species composition among sites is mainly due to the much stronger changes in Pre described (Devoto et al. unpubl.). The average difference in assemblage composition between adjacent sites, estimated as Sørensen distances, was 0.71 for flower visitors and 0.75 for plants (Appendix).
Table 1. Characteristics of eight complete plant-pollinator communities located in NW Patagonia, Argentina. For further details see Devoto et al. (2005). Degree of nestedness was measured using Aninhado software (Guimarães and Guimarães 2006).
Tabla 1. Características de ocho redes planta-polinizador completas localizadas en el NO de Patagonia, Argentina. Ver Devoto et al. (2005) para más detalles. El grado de anidamiento fue medido con el programa Aninhado (Guimarães y Guimarães 2006).
For each site, we constructed the plant-flower visitor interaction web as a binary matrix where ones represented observed visitations while zeroes represented absence of interaction (see Olesen & Jordano 2002 for a similar approach). The ecological properties of these pollination webs were described in detail elsewhere (Devoto et al. 2005; Devoto 2006). The original pollination webs as well as a dataset on the distribution of each species along the gradient are available from the first author upon request.
We used an algorithm written in Mathematica software (Wolfram Research, Champaign, Ill.) to model the extinction process of p-p networks. The following assumptions were made: a) An interaction observed among a p-p pair in one site is assumed to be allowed in any of the other sites. Thus, a regional matrix is constructed incorporating all plants, insects and interactions recorded across the eight sites, depicting all the "ecologically possible" interactions in the region. (b) Only insects followed rainfall shifts, as they are the more mobile components of pollination webs (in terms of their ability to alter their geographic range to cope with isohyet displacement). Plants, which may alter their ranges in a much longer time scale, were not simulated to move between sites and their survival was measured after the movement of insects. (c) All flower visitors were considered equally abundant and equally efficient as flower visitors of the plants they visited. (d) Compensatory mechanisms of plants (e.g. automatic self-pollination, vegetative propagation, etc) were not considered. Thus, any plant without flower visitors was considered extinct. e) Shifts in floral specificity of insects (e.g. start to visit a new plant species in the absence of the original partner) were not allowed. Thus, any flower visitor without proper plants to visit i.e. those with which it interacted in the regional matrix) was considered extinct. (f) A parameter p, equal for all 8 sites, is introduced which governs the proportion (between 0 and 1.0, with 0.1 fixed intervals) of flower visitors migrating westward from one site to the next adjacent one.
For each parameter p the following steps were iterated: (1) A random set of flower visitor species for a fixed value of p was selected from each site and moved to the contiguous site to the west. In this way, each site gained new flower visitors "immigrating" from the adjacent site to the east and lost flower visitors "emigrating" to the adjacent site to the west. This step thus created a new assemblage of flower visitors at each site. As the easternmost matrix, CT, only lost species, it was not included in calculations. (2) All the ecologically possible interactions between the already existing plants and the new assemblage of flower visitors were determined for each site. To this end, the records of the regional matrix were used as a reference. For example, in any given new matrix, if the insect "A" and the plant "B" had been recorded interacting in any of the eight original matrices, then an interaction between both species was established. (3) The proportion of plant and flower-visitor species at each site that were left without interactions (i.e. had gone "extinct") was calculated. In the case of plants, this proportion was measured with respect to all the plants present, whereas in the case of flower visitors it was referred only to the migrating animals. These two measures are thus not comparable. In order to obtain statistically valid results, 100 iterations of the above steps were taken for each parameter p.
Additionally, as a first step to relate the structural features of the pollination webs to their robustness, we measured the degree of nestedness (Bascompte et al. 2003) of the eight local and the regional webs with Aninhado software (Guimarães & Guimarães 2006). In addition, as a way to calculate the probability that a matrix with a given degree of nestedness might be randomly produced, for each real network we generated 1000 random networks using the CE null model in Aninhado. This model in equivalent to Model #2 of Bascompte et al. (2003) and allows separating two confounding interaction patterns that can cause nestedness: heterogeneity in number of interactions per species and true asymmetric ecological specialization (see also Model #2 of Vázquez & Aizen 2003 for a somewhat similar approach).
Finally, we evaluated to what extent assuming that when a link between a plant species and a flower visitor species is observed at one site, it will also occur at another site, provided both the plant and the flower visitor species are present (assumption a). For each plant-flower visitor interaction of the regional matrix we calculated the number of sites in which the pair was observed interacting, and the total number of sites where the pair existed (interacting or not).
RESULTS
The average proportion of extinct species across sites and migration proportions was 0.366 (range: 0.363-0.370) for flower visitors, and 0.076 (0.024-0.102) for plants (n=100 randomizations per site for both guilds). However, there were notable differences in the proportion of extinct species between sites, both among flower visitors and plants. These between-site differences remained roughly constant under different percentages of migration (Figure 1). Across sites, the proportion of extinction was moderate among flower visitors and remained roughly constant under increasing migration percentages. This pattern is the result of the way calculations were made, and should be interpreted cautiously (see Discussion). For plants, however, there was an evident increase in the proportion of extinctions with increasing migration of flower visitors (Figure 2), although the maximum values achieved were quite low (roughly 0.1; Figure 2). Overall, the plant-flower visitor systems of Andean Patagonia seemed to be quite resilient to a westward shift in the range of flower visitors.


Figure 1. Proportion (mean ± SE) of flower visitors (left) and plants (right) "extinct" (i.e. left without mutualists) as a function of the migration of different proportions (0.2-0.8, indicated above each graph) of flower visitors (n = 100 randomizations).
Figura 1. Proporción (media ± ES) de visitantes florales (izq.) y plantas (der.) "extinguidos" (i.e. que quedaron sin mutualistas) en función de la migración de diferentes proporciones (0.2-0.8, indicado sobre cada gráfico) de visitantes florales (n = 100 repeticiones aleatorias).

Figure 2. Proportion of extinct plant (![]() ) and flower visitor (
) and flower visitor (![]() ) species at each percentage of flower visitor migration (mean ± SE). Average across the seven sites. Both curves were included in the same graphs for simplicity, but they are not directly comparable as they were calculated differently (see Discussion).
) species at each percentage of flower visitor migration (mean ± SE). Average across the seven sites. Both curves were included in the same graphs for simplicity, but they are not directly comparable as they were calculated differently (see Discussion).
Figura 2. Proporción de especies de plantas (![]() ) y visitantes florales (
) y visitantes florales ( ![]() ) extinguidos según distintos porcentajes de migración de visitantes florales (media ± ES). Promedio de los siete sitios. Por simplicidad, ambas curvas fueron incluidas en el mismo gráfico, pero no son comparables entre sí debido a que fueron calculadas de diferente manera (ver Discusión).
) extinguidos según distintos porcentajes de migración de visitantes florales (media ± ES). Promedio de los siete sitios. Por simplicidad, ambas curvas fueron incluidas en el mismo gráfico, pero no son comparables entre sí debido a que fueron calculadas de diferente manera (ver Discusión).
Concerning the degree of nestedness, all webs were significantly nested. For all webs, including the regional web (Figure 3), the probability that the matrix was randomly generated was P<0.05 (Monte Carlo test; n=1000 runs; Table 1).

Figure 3. Regional matrix of interactions between 113 plant (columns) and 418 flower visitor (rows) species constructed from eight pollination webs from NW Patagonia, Argentina (for further details on original webs see Devoto et al., 2005). Four types of interactions are labelled, A: generalist plants interacting with generalist flower visitors; B: specialist plants interacting with generalist flower visitors; C: specialist flower visitors interacting with generalist plants; D: specialist plants interacting with specialist flower visitors.
Figura 3. Matriz regional de interacciones entre 113 especies de plantas (columnas) y 418 especies de visitantes florales (filas) construida a partir de ocho redes de polinización del NO de Patagonia, Argentina (ver Devoto et al., 2005 para más detalles sobre las redes originales). Se indican cuatro tipos de interacciones, A: plantas generalistas que interactúan con visitantes florales generalistas; B: plantas especialistas que interactúan con visitantes florales generalistas; C: visitantes florales especialistas que interactúan con plantas generalistas; D: plantas especialistas que interactúan con visitantes florales especialistas.
Finally, the evaluation of the validity of assumption a showed that out of 1056 interactions in the regional matrix, 369 (34%) could have occurred in more than one site. Out of these possible interactions, only 122 (33%) were actually measured in the field, and on average occurred in 55% of the possible sites (Figure 4).

Figure 4. Distribution of number of sites where flower visitor interactions occurred. From top to bottom, each graph corresponds to a subset of interactions which could have occurred in a maximum potential number of sites. For this, a site was considered to be able to accommodate a given interaction only if the plant and the flower-visitor species involved had been recorded interacting at least once in any one site. Additionally, the total number of interactions in each subset is provided. For example, out of 222 interactions that could have occurred in a maximum of two sites (first graph), only 49 were recorded on two sites, while 173 were only recorded in one site.
Figura 4. Distribución del número de sitios en que ocurrió una interacción entre una planta y un visitante floral. De arriba hacia abajo, cada gráfico corresponde a un subconjunto de interacciones que podría haber ocurrido en un número máximo de sitios potenciales. Para esto, un sitio fue considerado capaz de registrar una determinada interacción sólo si la interacción entre las especies de planta y de visitante floral había sido registrada al menos una vez en cualquiera de los sitios. Además, se provee el número total de interacciones en cada subconjunto. Por ejemplo, de las 222 interacciones que podrían haber ocurrido en un máximo de dos sitios (primer gráfico), sólo 49 fueron efectivamente registradas en dos sitios, mientras que las restantes 173 fueron registradas en sólo un sitio.
DISCUSSION
Model assumptions
Given the unavoidable constraints of a modeling approach such as ours it should be considered in what ways the simplifying assumptions made could have influenced the results. Whereas some of the assumptions are likely to overestimate species´ extinction probabilities, others may cause the opposite effect. First, in light of the evidence that plant-flower visitor interaction networks are highly stochastic both spatially and temporally (Herrera 1988, 2005, Gómez & Zamora 1999; Minckley et al. 1999 p. 138 and references therein, Thompson 2001; Williams et al. 2001), assuming that when a link between a plant species and a flower visitor species is observed at one site, it will also occur at another site, provided both the plant and the flower visitor species are present (assumption a), is arguable. Indeed our results show that pairs of potential mutualists interact only in a fraction of the sites in which both are present.
Second, we only allowed flower visitors to migrate, whereas plants were "left to their fate" because we assume that flower visitors have higher dispersal ability than plants (assumption b). To our knowledge, there is no published evidence to validate this assumption, but it seems reasonably sound in light of the evident higher mobility of insects as compared to plants. Had we shifted plants at the same rate than flower visitors the extinct proportion of species of both guilds would have probably been much lower; in addition, if both guilds had been allowed to migrate but in an uncoupled way (that is, at different rates) it is safe to assume that the extinction figures would have been intermediate.
Third, we considered that all flower visitors of a plant species are equally abundant and efficient as pollinators (assumption c). Typically, in plant-flower visitor webs a rather small subset of the visitor species is conducting the majority of pollination of a plant species (Johnson and Steiner 2000; Fenster et al. 2004; Vázquez et al. 2005; Sahli and Conner 2006). However, it is close to impossible to actually measure the pollination contribution of each visitor to each plant species when dealing with more than 100 and 400 plant and flower visitor species, respectively, as it is our case. Introducing in our model variation, for instance, among visitors in their contribution as pollinators or among plants in abundance is an interesting theoretical refinement that would, nevertheless, complicate significantly our model beyond the objectives of this work. Another shortcoming somewhat related to the one just mentioned, and that is not restricted to this particular study, is the lack of consideration of interaction strength in plant-flower visitor interaction networks (Memmott 1999). For instance, a plant-flower visitor interaction observed only once in the field has, in our approach, an equal value as an interaction observed a hundred times. However, the additional amount of sampling effort needed to characterize interaction strengths among all pairs of interacting species determines that this shortcoming is unsolvable for our data set. Nevertheless, it remains a matter of speculation whether it would alter our results significantly if considered.
Fourth, we considered that any plant or flower visitor left with no interactions would become extinct. However, plants could exhibit compensatory mechanisms of reproduction, which are alternative to animal pollination, such as vegetative reproduction, pollinator-independent (i.e. automatic) self-pollination, etc. (Bond 1994). To grossly assess the extent to which this assumption biased our results, we compiled information on pollinator dependence of the local flora from Arroyo & Squeo (1990), Aizen & Basilio (1995), Riveros et al. (1996), Aizen & Ezcurra (1998), Devoto et al. (2006), and our own unpublished results. Given that these sources are rather disparate with respect to their methods each plant species was assigned to one of three groups of pollinator dependency if it matched at least one of a set of criteria (below), as follows: high dependency (entomophilous species, dioecious, highly selfincompatible, pollen-ovule ratio [P:O; Cruden 1977] indicating obligate xenogamy), moderate dependency (ambophilous species, gynomonoecious or andromonoecious flowers, partially self-compatible; P:O indicating facultative xenogamy or facultative autogamy) and low dependency (agamospermous species, highly self-compatible, high automatic fruit formation, P:O indicating obligate autogamy). Out of 113 plant species recorded in the field we found information on 60: 30 (50%) showed high dependence, 22 (36.7%) showed moderate dependence, and only 8 (13.3%) species showed low dependence (details available on request). In this context, our model would be overestimating extinction rates only moderately given the high dependency on pollinators evidenced in the subsample of plants we could characterize.
Fifth, we assumed a fixed floral specificity of insects (e.g. we did not allow an insect to start to visit a new plant species in the absence of the original partner): after migrations our model only established interactions previously extant in the regional matrix. However, both plants and flower visitors may be capable of finding alternative pollination partners when deprived from their original mutualists. For example, insects migrating to a new community could be capable of visiting a plant species which was not present in their original community but that has cues which allow the insects to identify it as a useful resource. This assumption may have inflated the extinction rates reported. In this context, it is useful to think about the ecology of biological invasions: very few introduced plants are known to have failed to establish in a new environment because of lack of pollinators, or other mutualistic partners and also there is evidence of mutualist shifts in plants and pollinators after invasion to a new environment (see recent reviews by Richardson et al. 2000 and Vázquez 2006).
Finally, we moved flower visitors by only one step to the west. This decision was based on the worst possible climate change scenario for the next century for Argentina (Hulme & Sheard 1999), in which an 18% decrease in mean annual precipitation is predicted for the region of our study. On average, this change would imply that the annual rainfall currently reported for each of our study sites (Table 1) would shift to the next site to the west. Although the effect of a movement of flower visitors of two sites to the west would have caused a larger disruption of pollination interactions than the one-site move we tested, it is not unreasonable to assume that the presence of regional generalists still would have significantly buffered extinction rates. One final concern related to this point is the lack of an appropriate null model against which to compare our extinction rates. Calculating extinction rates under an eastward range shift scenario or even under a random shift of species between any pair of sites would not render any useful measure to compare our results with.
Robustness of pollination webs and its causes
The significant turnover of species between the eight plant-flower visitor communities studied (Devoto et al. 2005) suggested that the simulated displacement of flower visitor ranges due to climate change would show catastrophic extinctions in pollination webs. However, our simulations in general only had a low (in the case of plants) to moderate (in the case of flower visitors) impact on the structure of the webs studied. But, direct comparisons between the responses of plants and flower visitors should be avoided as they were measured in different ways. In the case of plants, the interpretation of extinction rates is straightforward, as they were measured in relation to all plant species originally present in the community. For flower visitors, however, this proportion was measured with respect to the migrated species. In addition, the fairly stable extinction rate of pollinators throughout all the scenarios is owed to the fact that no matter how many species are selected randomly, each of them has a specific probability to survive the migration which is just given by the mismatch of potential plant species in the new habitat. Therefore, the constant extinction rates in each site irrespective of the migrating proportion (Figure 1) and the average proportion across all sites will be just the same (and only vary due to different random samples taken). Mean values from 100 randomizations from, say, 10% selected pollinators will be the same as from 100 randomizations from 50% pollinators drawn from the same population.
Assessing the effect of potential sampling biases on our results (Olesen & Jordano 2002) is difficult. If our dataset suffered from sampling-related flaws an additional sampling effort would have increased the number of species as well as of interactions detected. New species detected would probably be among the less abundant and with rather few interactions (Stang et al. 2006), which are also the more vulnerable components of ecological webs. In addition, an increased additional sampling would have detected new interactions which would have increased connectance (the proportion of potential interactions actually realized) of each community, a feature which is positively correlated to robustness (Dunne et al. 2002; Estrada 2007).
Three concomitant properties of these pollination webs seem to have contributed to their resilience to even the most extreme climate change scenarios simulated (50% migration and above). The three properties can only be appreciated by adopting a regional perspective on the pollination webs studied. The first two properties have been described in detail elsewhere (Devoto et al. submitted), although not in the context of robustness in the face of resilience to climate change. First, generalist species tend to be more geographically widespread than specialist species. In other words, there is a positive correlation both among plant and flower visitor species between regional degree of generalization (i.e. the overall number of pollination partners that one species has across the gradient) and geographic range (i.e. the number of sites where the species was found across the gradient) (Devoto et al. submitted). Second, regional and local estimates of degree of generalization are highly correlated for the generality of species. This results in a positive correlation between local degree of generalization (i.e. the number of pollination partners one species has at a particular site) and geographic range (i.e. the number of sites where the species was found across the gradient) both among plant and flower visitor species (Devoto et al. submitted). These two properties alone would suffice to explain a large proportion of the resilience to simulated climate change exhibited by the webs studied. As the more generalist species are also the more widespread they would not be seriously put at risk when shifted from one place to another (in the case of flower visitors) or when receiving new partners from a neighboring community (in the case of plants) as, given their generalist character, they would probably find a suitable partner in the new community where they are placed. However, a third property, regional-level nestedness, which is here first reported for any pollination system, adds an additional resilience cue to these systems.
The nested nature of assembly patterns has been reported recently for individual (i.e. based on a single community) plant-flower visitor webs (Petanidou & Ellis 1996; Bascompte et al. 2003). In a nested web "the more specialist species interact only with proper subsets of those species interacting with the more generalists" (Bascompte et al. 2003). In other words," the set of interactions recorded for any species is nested within any other more generalist species" (Jordano et al. 2006). This assembly pattern organizes the community around a central core of plant and flower visitor species that strongly interact among themselves. Nestedness has been claimed to have strong implications for community persistence as it can provide alternative routes for system responses to perturbations such as the elimination of a species or an interaction: a species is more unlikely to become "extinct" after the elimination of other species when embedded on such a highly cohesive network (Memmott et al. 2004; Jordano et al. 2006).
Nestedness can be appreciated in the regional matrix (constructed at the start of our modeling procedure) by sorting first rows (plants) and then columns (flower visitors) from the most generalist to the most specialist species (Figure 3). The salient feature that immediately arises is that both generalist and specialist species of one guild tend to interact mainly with generalists of the opposite guild. Thus, there is a nucleus of highly interacting species of both guilds (i.e. generalists interacting with generalists; region A in Figure 3) with two "tails" of specialists interacting with generalists, one of specialist plants interacting with generalist flower visitors (region B in Figure 3) and the other of specialist flower visitor interacting with generalist plants (region C in Figure 3). The regional nested pattern provides additional resilience to climate change because it implies that, regionally, the specialist species are linked to generalist (and thus more persistent) species. Just as it happens with generalists, this increases the chances that a specialist species will find a suitable pollination partner under any range-shift scenario.
Overall, we acknowledge that our assumptions represent a gross simplification of a rather complex phenomenon as is the extinction of species triggered by a regional range shift of species in response to climate change. Nevertheless, it seems that relaxing our assumptions would make the networks even more resilient than we report above, thus strengthening the conclusion that pollination networks are rather robust to climate change. However, it should also be highlighted that this study only focuses on the mutualistic interactions themselves, and does not regard other potential impacts on plants or flower visitors, some of which may be severe (e.g. drought, modification of larval habitats, phenological mismatch).
CONCLUSIONS
Our results support the idea that pollination networks are, at least in simulations, rather robust to the consequences of climate change. Memmott et al. (2004) explored probable patterns of extinction in two large networks of plants and flower visitors by simulating the removal of flower visitors and consequent loss of the plants they visited. Following systematic removal of flower visitors from the most generalized to the most specialized, they expected (such as we did with highest migration rates) catastrophic extinctions among plants. However, although the decline in species number was high (as compared to other sequential extinction criteria they also tested) declines were no worse than linear. They related this resilience to extinction to system symmetry (which, as mentioned above, causes a redundancy in flower visitors per plant) and to the nested pattern of interactions in their networks (Memmott et al. 2004).
The present comparative study of real geographically-related pollination webs reveals itself as a useful tool for understanding the possible consequences of predicted climate change on the structure and function of pollination systems. However, considering that our simulations are exclusively based on visitation webs there is a clear need to distinguish between mere visitors and those species which are efficient pollinators. These species are ultimately responsible for the resilience of pollination systems to human-induced perturbations which will surely increase in the near future. Simulations like the one we have showed, but applied to quantitative rather than qualitative webs, are likely to provide a useful insight into this issue.
All in all, our simulation can be viewed as a theoretical prediction against which real extinction rates in Patagonian plant-flower visitor webs could be compared in the near future, when predicted climate changes take place. The differences between our randomly-generated extinction rates and those that will actually occur may prove a useful benchmark to explore more in depth the extinction dynamics that will likely take place in the temperate forests of Patagonia.
APPENDIX
Sorensen distances between sites in (a) flower-visitor and (b) plant assemblage composition. Diagonal values in bold correspond to distances between sites that are adjacent in terms of mean annual rainfall (see Table 1 for details).

ACKNOWLEDGEMENTS
This work was funded with a grant from the Agencia Nacional de Promoción Científica y Tecnológica of Argentina (PICT 25450). An earlier version of this paper benefited from comments by N. Blüthgen, P. R. Guimarães, J. Memmott, D. P. Vázquez and N. M. Waser. D.M. is associated to CONICET, Argentina.
REFERENCES
AIZEN, MA; L ASHWORTH & L GALETTO. 2002. Reproductive success in fragmented habitats: do compatibility systems and pollination specialization matter? J. Veg. Sci. 13: 885-892. [ Links ]
AIZEN, MA & AM BASILIO. 1995. Within and among flower sex-phase distribution in Alstroemeria aurea (Alstroemeriaceae). Can. J. Bot. 73: 1986-1994. [ Links ]
AIZEN, MA & C EZCURRA. 1998. High incidence of plant-animal mutualisms in the woody flora of the temperate forest of southern South America: biogeographical origin and present ecological significance. Ecol. Aust. 8: 217-236. [ Links ]
AIZEN, MA & P FEINSINGER. 1994 a. Habitat fragmentation, native insect pollinators, and feral honey bees in Argentine "Chaco Serrano". Ecol. App. 4: 378-392. [ Links ]
AIZEN, MA & P FEINSINGER. 1994 b. Forest fragmentation, pollination, and plant reproduction in a Chaco Dry Forest, Argentina. Ecology 75: 330-351. [ Links ]
ARROYO, MK & F SQUEO. 1990. Relationship between plant breeding systems and pollination. Pp. 205-227 in: S Kawano (ed.). Biological approaches and evolutionary trends in plants. Academic Press, London. [ Links ]
ARROYO, MTK; L CAVIARES; A PEÑALOZA; M RIVEROS & A FAGGI. 1996. Phytogeographic relationships and species richness patterns in the temperate rain forest flora of South America. Pp. 71-100 in: J Armesto, C Villagrán & MTK Arroyo (eds.). Ecología de los bosques nativos de Chile. Editorial Universitaria, Santiago, Chile. [ Links ]
ARROYO, MTK; R PRIMACK & J ARMESTO. 1982. Community studies in pollination ecology in the high temperate Andes of Central Chile. I. Pollination mechanisms and altitudinal variation. Amer. J. Bot. 69: 82-97. [ Links ]
ASHWORTH L; R AGUILAR; L GALETTO & MA AIZEN. 2004. Why do pollination generalist and specialist plant species show similar reproductive susceptibility to habitat fragmentation? J. Ecol. 92: 717-719. [ Links ]
AUSTIN, AT & OE SALA. 2002. Carbon and nitrogen dynamics across a natural gradient of precipitation in Patagonia, Argentina. J. Veg. Sci. 13: 351-360. [ Links ]
BASCOMPTE, J; P JORDANO; CJ MELIÁN & JM OLESEN. 2003. The nested assembly of plant-animal mutualistic networks. PNAS (USA) 100: 9383-9387. [ Links ]
BOND, WJ. 1994. Do mutualisms matter? Assessing the impact of pollinator and disperser disruption on plant extinction. Phil. Trans. R. Soc. B 344: 83-90. [ Links ]
CRUDEN, RW. 1977. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants. Evolution 31: 32-46. [ Links ]
DAVIS, MB & RG SHAW. 2001. Range shifts and adaptive responses to Quaternary climate change. Science 292: 673-679. [ Links ]
DEVOTO, M; D MEDAN & NH MONTALDO. 2005. Patterns of interaction between plants and pollinators along an environmental gradient. Oikos 109: 461-472. [ Links ]
DEVOTO, M. 2006. Interacciones planta-polinizador a lo largo de un gradiente ambiental: una aproximación en escala de comunidad. Escuela para Graduados "Ing. Agr. Alberto Soriano", Facultad de Agronomía, Universidad de Buenos Aires. Tesis de Maestría. 130 pp. (http://agro.uba.ar/~mdevoto/Tesis-MSc.pdf). [ Links ]
DEVOTO, M; NH MONTALDO & D MEDAN. 2006. Mixed hummingbird—long-proboscid-fly pollination in "ornithophilous" Embothrium coccineum (Proteaceae) along a rainfall gradient in Patagonia, Argentina. Aust. Ecol. 31: 512-519. [ Links ]
DIXON, AFG. 2003. Climate change and phenological asynchrony. Ecol. Entom. 28: 380-381. [ Links ]
DUNNE, JA; RJ WILLIAMS & ND MARTINEZ. 2002. Network robustness and biodiversity loss in food webs: robustness increases with connectance. Ecol. Lett. 5: 558-567. [ Links ]
ELBERLING, H & JM OLESEN. 1999. The structure of a high latitude plant-flower visitor system: the dominance of flies. Ecography 22: 314-323. [ Links ]
ESTRADA, E. 2007. Food webs robustness to biodiversity loss: The roles of connectance, expansibility and degree distribution. J. Theor. Biol. 244: 296-307. [ Links ]
FENSTER, CB; WS ARMBRUSTER; P WILSON; MR DUDASH & JD THOMSON. 2004. Pollination syndromes and floral specialization. Annu. Rev. Ecol. Evol. Syst. 35: 375-403. [ Links ]
GÓMEZ, JM & R ZAMORA. 1999. Generalization vs. specialization in the pollination system of Hormathophylla spinosa (Cruciferae). Ecology 80: 796-805. [ Links ]
HERRERA, CM. 1988. Variation in mutualisms: the spatio-temporal mosaic of a pollinator assemblage. Biol. J. Linn. Soc. 35: 95-125. [ Links ]
HERRERA, CM. 2005. Plant generalization on pollinators: species property or local phenomenon? Amer. J. Bot. 92: 13-20. [ Links ]
HUGHES, L. 2000. Biological consequences of global warming: is the signal already apparent? Trends Ecol. Evol. 15: 56-61. [ Links ]
HULME, M & N SHEARD. 1999. Climate Change Scenarios for Argentina. Climatic Research Unit, Norwich, UK. [ Links ]
JOHNSON, SD & KE STEINER. 2000. Generalization versus specialization in plant pollination systems. Trends in Ecology and Evolution 15: 140-143. [ Links ]
JORDANO, P; J BASCOMPTE & JM OLESEN. 2003. Invariant properties in coevolutionary networks of plantanimal interactions. Ecol. Lett. 6: 69-81. [ Links ]
JORDANO, P; J BASCOMPTE & JM OLESEN. 2006. The ecological consequences of complex topology and nested structure in pollination webs. Pp. 173-199 in: NM Waser & J Ollerton (eds.). Plant-Pollinator Interactions. From specialization to generalization. University of Chicago Press, Chicago. [ Links ]
KEARNS, CA; DW INOUYE & NM WASER. 1998. Endangered mutualisms: the conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 28: 83-112. [ Links ]
MALO, JE & J BAONZA. 2002. Are there predictable clines in plant-pollinator interactions along altitudinal gradients? The example of Cytisus scoparius (L.) Link in the Sierra de Guadarrana (Central Spain). Div. Distr. 8: 365-371. [ Links ]
MCCARTHY, JJ; OF CANZIANI; NA LEARY; DJ DOKKEN & KS WHITE. 2001. Climate Change 2001: Impacts, Adaptations, and Vulnerability. Cambridge University Press, New York. [ Links ]
MEDAN, D; NH MONTALDO; M DEVOTO; A MANTESE; V VASELLATI; GG ROITMAN & NH BARTOLONI. 2002. Plant-pollinator relationships at two altitudes in the Andes of Mendoza, Argentina. Arc. Antarc. Alp. Res. 34: 233-241. [ Links ]
MEMMOTT, J. 1999. The structure of a plant-pollinator food web. Ecol. Lett. 2: 276-280. [ Links ]
MEMMOTT, J; NM WASER & M PRICE. 2004. Tolerance of pollination networks to species extinctions. Proc. R. Soc. B 271: 2605-2611. [ Links ]
MEMMOTT, J & NM WASER. 2002. Integration of alien plants into a native flower-pollinator visitation web. Proc R. Soc. B 269: 2395-2399. [ Links ]
MINCKLEY, LR; JH CANE; L KERVIN & TH ROULSTON. 1999. Spatial predictability and resource specialization of bees (Hymenoptera: Apoidea) at a superabundant, widespread resource. Biol. J. Linn. Soc. 67: 119-147. [ Links ]
MORALES, CL & MA ALZEN. 2002. Does invasion of exotic plants promote invasion of exotic flower visitors? A case study from the temperate forests of the southern Andes. Biol. Inv. 4: 87-100. [ Links ]
MOVIA, CP; GH OWER & CE PÉREZ. 1982. Estudio de la vegetación natural de la Provincia del Neuquén. Ministerio de Economía y Hacienda, Subsecretaría de Estado de Recursos Naturales, Argentina. [ Links ]
OLESEN, J & P JORDANO. 2002. Geographic patterns in plant-pollinator mutualistic networks. Ecology 83: 2416-2424. [ Links ]
OLLERTON, J & L CRANMER. 2002. Latitudinal trends in plant-pollinator interactions: are tropical plants more specialised? Oikos 98: 340-350. [ Links ]
PARUELO, JM; AM BELTRÁN; E JOBBÁGY; OE SALA & RA GOLLUSCIO. 1998a. The climate of Patagonia: general patterns and controls on biotic processes. Ecol. Aust. 8: 85-101. [ Links ]
PARUELO, JM; EG JOBBAGY & OE SALA. 1998b. Biozones of Patagonia (Argentina). Ecol. Aust. 8: 145-153. [ Links ]
PEÑUELAS, JI; I FILELLA; SW ZHANG; L LLORENS; R OGAYA; F LLORET ET AL. 2004. Complex spatiotemporal phenological shifts as a response to rainfall changes. New Phytol. 161: 837-846. [ Links ]
PETANIDOU, T & WN ELLIS. 1996. Interdependence of native bee faunas and floras in changing Mediterranean communities. Pp. 201-226 in: A Matheson; SL Buchmann; C O´Toole; P Westrich& IH Williams (eds.). The conservation of bees. Academic Press, London. [ Links ]
PRICE, MV & NM WASER. 1998. Effects of experimental warming on plant reproductive phenology in a subalpine meadow. Ecology 79: 1261-1271. [ Links ]
RICHARDSON, DM; N ALLSOPP; CM D´ANTONIO; SJ MILTON & M REJMÁNEK. 2000. Plant invasions-the role of mutualisms. Biol. Rev. 75: 65-93. [ Links ]
RIVEROS, GM; PAM HUMAÑA & MK ARROYO. 1996. Sistemas de reproducción en especies del bosque valdiviano (40º Latitud Sur). Phyton 58: 167-176. [ Links ]
ROOT, TL; JT PRICE; KR HALL; SH SCHNEIDER; C ROSENZWEIG & JA POUNDS. 2003. Fingerprints of global warming on wild animals and plants. Nature 421: 57-60. [ Links ]
SAHLI, HF & JK CONNER. 2006. Characterizing ecological generalization in plant-pollination systems. Oecologia 148: 365-372. [ Links ]
STEFFEN, WL; RJ SCHOLES; C VALENTIN; X ZHANG & JC MENAUT. 1999. The IGBP terrestrial transects. Pp. 66-87 in: BH Walker; WL Steffen; J Canadell & JSI Ingram (eds.). The terrestrial biosphere and global change. Implications for natural and managed ecosystems. Cambridge University Press, New York. [ Links ]
THOMPSON, JD. 2001. How do visitation patterns vary among pollinators in relation to floral display and floral design in a generalist pollination system? Oecologia 126: 386-394. [ Links ]
THÓRHALLSDÓTTIR, TE. 1998. Flowering phenology in the central highland of Iceland and implications for climatic warming in the Arctic. Oecologia 114: 43-49. [ Links ]
TOTLAND, O. 1993. Pollination in alpine Norway: flowering phenology, insect visitors, and visitation rates in two plant communities. Can. J. Bot. 71: 1072-1079. [ Links ]
VÁZQUEZ, DP. 2005. Degree distribution in plant-animal mutualistic networks: forbidden links or random interactions? Oikos 108: 421-426. [ Links ]
VÁZQUEZ, DP. 2006. Exploring the relationship between niche breadth and invasion success. Pp. 307-322 in: MW Cadotte; SM McMahon & T Fukami (eds.). Conceptual ecology and invasions biology: reciprocal approaches to nature. Springer, New York. [ Links ]
VÁZQUEZ, DP & MA AIZEN. 2003. Null model analyses of specialization in plant-pollinator interactions. Ecology 84: 2493-2501. [ Links ]
VÁZQUEZ, DP & MA AIZEN. 2004. Asymmetric specialization: a pervasive feature of plant-pollinator interactions. Ecology 85: 1251-1257. [ Links ]
VÁZQUEZ, DP & D SIMBERLOFF. 2003. Changes in interaction biodiversity induced by an introduced ungulate. Ecol. Lett. 6: 1077-1083. [ Links ]
VÁZQUEZ, DP & D SIMBERLOFF. 2004. Indirect effects of introduced ungulates on pollination and plant reproduction. Ecol. Monog. 74: 281-308. [ Links ]
VÁZQUEZ DP, WF MORRIS & P JORDANO. 2005. Interaction frequency as a surrogate for the total effect of animal mutualists on plants. Ecol. Lett. 9: 1088-1094. [ Links ]
VITOUSEK, PM & PA MATSON. 1991. Gradient analysis of ecosystems. Pp 287-298 in: J Cole; G Lovett & S Findlay (eds.). Comparative analyses of ecosystems: patterns, mechanisms and theories. Springer, New York. [ Links ]
WALTHER, G-R; E POST; P CONVEY; A MENZEL; C PARMESAN; TJC BEEBEE ET AL. 2002. Ecological responses to recent climate change. Nature 416: 389-395. [ Links ]
WATT, AD & AM MCFARLANE. 2002. Will climate change have a different impact on different trophic levels? Phenological development of winter moth Operophtera brumata and its host plants. Ecol. Entom. 27: 254-256. [ Links ]
WILLIAMS, NM; RL MINCKLEY & FA SILVERA. 2001. Variation in native bee faunas and its implications for detecting community change. Cons. Ecol. 5: 57-89. [ Links ]
ZEGERS, CD 1993. Bosques templados de Chile y Argentina. Variación, Estructura y Dinámica. Editorial Universitaria, Santiago, Chile. [ Links ]

