










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.17 no.1 Córdoba Jan./June 2007
SECCIÓN ESPECIAL
Fragmentación de hábitat, riqueza de polinizadores, polinización y reproducción de plantas nativas en el Bosque Chaqueño de Córdoba, Argentina
Leonardo Galetto*, Ramiro Aguilar**, Mariana Musicante, Julia Astegiano, Ana Ferreras, Mariana Jausoro, Carolina Torres, Lorena Ashworth & Cecilia Eynard
Instituto Multidisciplinario de Biología Vegetal - CONICET, Universidad Nacional de Córdoba, Córdoba, Argentina.
* Grupo de investigaciones en ecología reproductiva de plantas, Instituto Multidisciplinario de Biología Vegetal - CONICET, Universidad Nacional de Córdoba, CC 495, (5000) Córdoba, Argentina. Tel. y Fax: (54) 0351-4332104. Email: leo@imbiv.unc.edu.ar **raguilar@imbiv.unc.edu.ar
Recibido: 11 de abril de 2007;
Fin de arbitraje: 25 de mayo de 2007;
Revisión recibida: 4 de junio de 2007;
Aceptado: 7 de junio de 2007
RESUMEN. En este trabajo se analiza la riqueza de polinizadores y la polinización y reproducción de plantas en un gradiente de fragmentación en el bosque Chaqueño de Córdoba, Argentina. Se espera encontrar una relación directa entre el área de los fragmentos, la riqueza de polinizadores y estos dos procesos. A partir de los datos obtenidos por varios investigadores, se plantearon los siguientes objetivos: i) evaluar, en la escala de sitio, la relación entre el área de los fragmentos, la riqueza de polinizadores y la polinización (medida como frecuencia de visitas a las flores), ii) analizar, en la escala de un conjunto de especies de plantas, los patrones de respuesta para variables relacionadas con la polinización (riqueza de polinizadores, frecuencia promedio de visitas) y reproducción (producción de frutos) considerando un gradiente de fragmentación, iii) comparar las tendencias obtenidas a partir de estas dos aproximaciones metodológicas y iv) discutir los resultados en el contexto de la conservación de la biodiversidad. No se encontró una menor riqueza o frecuencia de visitas de los polinizadores a medida que disminuye el área de los fragmentos. Los resultados obtenidos a través de meta-análisis muestran que las magnitudes del efecto global fueron significativamente diferentes de cero para la riqueza de polinizadores y la producción de frutos. Por el contrario, no se detectó un efecto global significativo en relación a la frecuencia de visitas de los polinizadores. Las tendencias encontradas considerando las dos aproximaciones no fueron uniformes. Las respuestas de los polinizadores y las plantas resultaron más complejas que lo planteado a priori, ya que si se analiza la heterogeneidad en las tendencias encontradas en el conjunto de especies de plantas estudiado dentro del gradiente de fragmentación se sugiere una susceptibilidad especie-específica. Asimismo, varias de las especies que fueron estudiadas en distintas temporadas mostraron variaciones en las tendencias de las variables consideradas en relación a la disminución del área de los fragmentos. Para políticas de conservación del bosque Chaqueño, sería interesante considerar aquellas especies de plantas y sus polinizadores asociados que estarían siendo afectadas negativamente por la fragmentación de hábitat.
Palabras clave: Comunidades; Fragmentación de hábitat; Meta-análisis; Polinización; Región Chaqueña.
ABSTRACT. Pollinator richness, pollination and plant reproduction are analyzed in a fragmentation gradient of the Chaco forests of Córdoba, Argentina. We expect to find direct relationships between pollinator richness, these two processes and the area of the fragments. Considering data obtained by many researchers for different forest sites, the following objectives were planned: i) to evaluate the relationship between fragment area, pollinator richness and pollination (measure as frequency of visits to the flowers), ii) to analyze at the species level the response patterns of variables related to the pollination (pollinator richness or frequency of visits) and reproduction (i.e., fruit production) processes within a gradient of fragmentation for many plants, iii) to compare trends obtained using different methodologies, and iv) to discuss results in the context of biodiversity conservation. Contrarily to our expectations, direct relationships between fragment area and pollinator richness or frequency of floral visits were not found. On the other hand, results obtained for pollinator richness and fruit set by meta-analyses showed significant overall fragment size effect. The frequency of visits, on the contrary, did not show a significant overall size effect. The trends observed using the two methodologies were not uniform. Pollinator and plant responses to fragmentation seem to be more complex than what was originally predicted. If the heterogeneity of trends found for many species within a fragmentation gradient is analyzed, a species-specific susceptibility is suggested. In addition, many species have showed variability among seasons in their tendencies within the gradient of fragmentation. When discussing conservation politics of the Chaco forests of Córdoba, it would be interesting to analyze those plant species and their pollinators that seem to be affected negatively by reduction of fragment size.
Keywords: Chaco region; Communities; Habitat fragmentation; Meta-analysis; Pollination.
INTRODUCCIÓN
Se considera que nuestro planeta está enfrentando una de las mayores extinciones de especies en los últimos 65 millones de años (e.g., Wilson 1992). La destrucción de hábitats naturales y su fragmentación por las actividades humanas son dos de los principales procesos conductores de esta crisis global de la biodiversidad (e.g., Noss & Cooperrider 1994; Fahrig 2003; Hobbs & Yates 2003; Henle et al. 2004), existiendo un amplio consenso entre los investigadores que los estudios sobre paisajes fragmentados son urgentes y deben ser considerados prioritarios (Saunders et al. 1991). El proceso de fragmentación de hábitat es conceptualmente complejo y puede subdividirse considerando la pérdida de hábitat, entendida como la disminución total del área del hábitat original a partir de una comparación histórica, y la fragmentación per se, referida a la creación de nuevos fragmentos de hábitat cada vez más pequeños y aislados entre sí (Fahrig 2003).
El proceso de fragmentación de hábitat determina la reducción de las poblaciones locales de diferentes organismos y un mayor aislamiento entre ellas. Sin embargo, la respuesta de las plantas y animales frente a la fragmentación de hábitat puede variar, dependiendo de las características relacionadas con su reproducción, dispersión o demografía (e.g., Saunders et al. 1991; Van Dyck & Matthysen 1999; Aguilar et al. 2006). La relación entre el área y la diversidad ha sido ampliamente estudiada (e.g., Jacquemyn et al. 2001 y bibliografía citada allí). En cambio, las consecuencias de las posibles alteraciones en las interacciones planta-animal en relación a la fragmentación de hábitat a escala de la comunidad han sido comparativamente poco estudiadas (Fisher & Matthies 1998; Kearns et al. 1998; Lienert 2004).
Los cambios en la configuración del hábitat disponible pueden modificar las interacciones mutualistas entre plantas y animales; por ejemplo, cuando se comparan sitios continuos con fragmentos pequeños se ha encontrado una disminución tanto en la riqueza de polinizadores, como en la polinización o fructificación de las plantas (e.g., Aizen & Feinsinger 1994; Steffan-Dewenter & Tscharntke 1999; Cane 2001; Aguilar 2005). Varias especies muestran una disminución significativa en la polinización que se explicaría, principalmente, por las modificaciones en la diversidad o abundancia de la fauna polinizadora (Rathcke & Jules 1993; Aizen & Feinsinger 1994). En las plantas, la disminución en la polinización puede repercutir en forma directa sobre la producción de frutos y semillas (e.g., Aguilar & Galetto 2004). Como consecuencia, otras etapas del ciclo de vida de las plantas también podrían verse afectadas negativamente y eventualmente desencadenar el colapso demográfico de las poblaciones de algunas especies (e.g., Jules 1998; Yao et al. 1999; Lennartsson 2002).
El Bosque Chaqueño de Córdoba está actualmente reducido a fragmentos de distinto tamaño, donde la pérdida de hábitat por las actividades humanas durante los últimos 30 años alcanzó tasas sin precedentes (Gavier & Bucher 2004; Zak et al. 2004). Estudios realizados en comunidades del Bosque Chaqueño muestran que muchas de las especies de plantas son polinizadas por unas pocas o bien por numerosas especies de animales (sistemas de polinización especializado o generalizado, respectivamente), y que su reproducción depende fuertemente de los polinizadores (e.g., Torres 2003; Ashworth 2004; Aguilar 2005), tanto de las especies autoincompatibles como de las autocompatibles (e.g., Morales & Galetto 2003). Además, se ha encontrado una relación directa y consistente temporalmente entre el área de los fragmentos, la polinización y el éxito reproductivo para seis de un total de 14 especies de plantas nativas estudiadas (Aguilar 2005).
El objetivo general de este trabajo es analizar si la riqueza de polinizadores, la polinización y la reproducción de las plantas a escala de la comunidad se relaciona con el área de los fragmentos en el bosque Chaqueño de Córdoba, a partir de los datos disponibles desde 1999 para distintos sitios de bosque que conforman un gradiente de fragmentación y que han sido obtenidos por varios integrantes de nuestro grupo de trabajo. El estudio de los posibles cambios en la diversidad de polinizadores, la polinización y el análisis de las consecuencias reproductivas para las plantas serán de gran importancia en el contexto de la conservación a escala regional.
Para evaluar esos patrones generales se dispone de dos grupos de datos recolectados en el mismo gradiente de fragmentación. El primer grupo de datos fue obtenido a escala de sitio, para estimar la riqueza de polinizadores y la polinización en cada uno de los sitios de bosque, mientras que el segundo grupo fue obtenido a escala de numerosas especies de plantas focales que presentan poblaciones en varios de esos sitios, para evaluar los cambios en la riqueza de polinizadores y sobre la polinización y reproducción de las plantas en relación al área de los fragmentos. Esto es, entre los datos disponibles para más de 100 especies de plantas de la región, se seleccionaron los correspondientes a 49 de esas especies en las que han podido medirse distintas variables en cinco o más sitios de bosque que pueden incluirse dentro de un gradiente de fragmentación. La idea subyacente es realizar un análisis comparativo utilizando dos escalas de aproximación tendiente a evidenciar tendencias en relación al área de los fragmentos y riqueza de polinizadores y con variables relacionadas a la polinización y reproducción de las plantas. A partir de estos dos grupos de datos se plantearon los siguientes objetivos particulares: i); evaluar a escala de sitio la relación entre el área de los fragmentos, la riqueza de polinizadores y la polinización (medida como frecuencia de visitas a las flores), ii) analizar a escala de un conjunto de especies de plantas los patrones de respuesta para variables relacionadas a la polinización (riqueza de polinizadores, frecuencia promedio de visitas) y reproducción (producción de frutos) considerando un gradiente de fragmentación, iii) comparar las tendencias obtenidas a partir de estas dos aproximaciones metodológicas y vi) discutir los resultados considerando las posibles consecuencias de la fragmentación de hábitat sobre la polinización y la reproducción de las plantas en el contexto de la conservación de la biodiversidad.
La hipótesis general de trabajo propone que a medida que disminuye el área de los fragmentos de bosque se produce una disminución en la calidad y heterogeneidad del hábitat, lo cual afecta las condiciones de alimentación y reproducción de los polinizadores. Este fenómeno determina que disminuya la diversidad de polinizadores en los fragmentos de bosque y como consecuencia se ve afectada la capacidad reproductiva de la plantas polinizadas por animales. Como consecuencia, se espera encontrar una relación directa entre el área de los fragmentos, la riqueza de polinizadores, la polinización y reproducción de las plantas.
MATERIALES Y MÉTODOS
Área de estudio y selección de los sitios de muestreo
Los sitios de estudio se encuentran en la Provincia de Córdoba, en los Departamentos Santa María, Colón y Punilla, sobre el faldeo oriental de las Sierras Chicas. Se utilizaron un total de 16 sitios, tres áreas consideradas como bosque continuo (i.e., de 1000 ha como superficie mínima) y 13 fragmentos de distintos tamaños, cubriendo un rango de 0.4 a ca. 116 ha. Se seleccionaron fragmentos espacialmente dispersos en el terreno, rodeados de una matriz de cultivos, generalmente de soja o maíz. Todos los lugares se encuentran entre los 500 y 870 m s. n. m. Las geo-referencias y el área de todos los sitios se presentan en la Tabla 1.
Tabla 1. Referencias geográficas, área (ha), riqueza total de polinizadores y riqueza y frecuencia de visitas florales de himenópteros polinizadores para 16 sitios de Bosque Chaqueño en un gradiente de fragmentación en el centro de Argentina. Abreviaturas: sd= sin datos.
Table 1. Geographic references, area (ha), total richness of pollinators, and frequency of floral visits by hymenopterans for 16 sites of Chaco forests within a fragmentation gradient in central Argentina. Abbreviations: sd: no data.

Los datos fueron tomados entre los años 1999 y 2006 por diferentes investigadores (cfr. Apéndice). Cada estudio desarrolló objetivos particulares y, como consecuencia, el número de sitios de estudio, el tiempo total dedicado a las observaciones y la metodología tuvo algunas variantes según el caso. Por ello, las variables fueron estandarizadas para poder ser analizadas.
Los insectos fueron fotografiados o capturados y acondicionados para su transporte a laboratorio, para luego identificarlos. Las plantas donde se recolectaron los polinizadores fueron herborizadas e identificadas y se encuentran depositadas en el Herbario del Museo Botánico de la Universidad Nacional de Córdoba (CORD).
Variables
Riqueza de polinizadores: Esta variable fue obtenida durante la temporada húmeda (octubre-mayo). El criterio utilizado para considerar que se trataba de polinizadores fue observar si los animales hacían contacto con los órganos reproductivos de la flor. Cada sitio fue visitado en distintas oportunidades realizándose observaciones de 10 minutos, distribuidos a lo largo del día. El número total de minutos de observación por sitio fue variable según cada estudio (150 a 1800 minutos). Los muestreos se realizaron sobre transectas en cada sitio. El punto inicial de observación y la dirección de las transectas fueron determinados al azar. Los muestreos dentro de cada transecta estuvieron separados al menos por 5 m. En cada muestreo se registró el total de especies de polinizadores (o de himenópteros; ver más abajo) que interactuaban con todas las especies con flores abiertas en un círculo de 1m de diámetro previamente delimitado, en parcelas de 2 x 2 m y hasta 1.80 m de altura, o bien con algunas especies de plantas focales, dependiendo del estudio. La riqueza de polinizadores de cada sitio, o de cada planta focal en un determinado sitio, corresponde al total de especies de polinizadores (o de himenópteros en algunos casos) registrado durante una determinada temporada. Dado que existieron diferencias en el esfuerzo de muestreo realizado en distintos estudios para estimar la riqueza de polinizadores, los grupos de datos fueron analizados en forma separada (por investigador, por temporada o por especie) ya que lo que se intenta evidenciar son las tendencias en los datos en relación al área de los fragmentos.
Frecuencia de visitas: Existen numerosas variables que pueden relacionarse con el proceso de polinización. Se utilizó esta variable ya que fue considerada en cada uno de los estudios disponibles para la región. A partir de conocer el número de flores abiertas sobre el que se registró la identidad del polinizador y el número de individuos de cada especie de planta en cada uno de los muestreos, se pudo calcular para cada sitio, o para cada especie de planta focal, la frecuencia de visitas promedio por flor y por unidad de tiempo. De esta manera, esta variable quedó estandarizada. Los datos fueron tomados dentro del período comprendido entre las 9 y las 17 h, en general en días soleados y de escaso viento.
Producción de frutos: Para evaluar la reproducción de las plantas se pueden utilizar distintos parámetros relacionados a la función masculina y femenina de la flor. Se utilizó esta variable ya que fue medida en la mayoría de los estudios realizados en este gradiente de fragmentación. Para cada una de las especies focales se seleccionaron varios individuos por sitio. Se marcaron flores abiertas y se las siguió hasta la maduración de los frutos. Para cada especie la producción de frutos por sitio se estimó a partir de la relación frutos maduros obtenidos / flores abiertas marcadas, siendo ésta una medida del éxito reproductivo.
Análisis de datos
Para evaluar estas variables en el gradiente de fragmentación se dispuso de dos grupos de datos, uno para cada uno de los sitios, y otro para varias plantas focales con poblaciones en varios sitios de ese gradiente. Con el primer grupo de datos se realizaron análisis de regresión lineal entre el área de los sitios de bosque y la riqueza de especies de polinizadores y la polinización (medida como frecuencia promedio de visitas por flor). El área de los fragmentos fue transformada a logaritmo en base 10. Se realizaron análisis de regresión con distintos grupos de datos ya que: i) fueron tomados en temporadas diferentes y ii) en un caso corresponde a la riqueza de total de polinizadores y en otros dos a la riqueza de himenópteros polinizadores (más detalles en Fig. 1). Se considera que se pueden comparar las tendencias obtenidas a partir de distintos grupos de datos ya que los himenópteros representan alrededor del 70% de las especies de polinizadores en distintas comunidades, siendo el grupo de insectos más importante relacionado a la polinización (Steffan-Dewnter & Tscharntke, 1999 y trabajos citados allí; Cane 2001).
Figura 1. Análisis de regresión lineal entre el área de los fragmentos de bosque y (a) la riqueza de polinizadores, (b) la polinización, estimada como frecuencia promedio de visitas de polinizadores por flor y por minuto en cada sitio para distintas temporadas y grupos de animales (cuadrados = total de especies de polinizadores para 16 sitios en 2004, círculos = himenópteros para 9 sitios en 2005 y triángulos = himenópteros para 9 sitios en 2006). Resultados obtenidos a través del análisis de regresión para cada grupo de datos: riqueza total de polinizadores R2= 0.32; P= 0.022, riqueza de himenópteros polinizadores en 2005 y 2006 R2= 0.10; P= 0.39 y R2= 0.08; P= 0.47, respectivamente; frecuencia promedio de visitas: R2= 0.27; P= 0.47 y R2= 0.05; P= 0.90 para las temporadas 2005 y 2006, respectivamente.
Figure 1. Regression analyses between the area of the fragments and (a) pollinator richness, (b) pollination, measured as mean frequency of pollinator visits by flower and by minute of each site for different seasons and for different groups of animals (squares = total species of pollinators for 16 sites in 2004, circles = Hymenopterans for 9 sites in 2005, and triangles = Hymenopterans for 9 sites in 2006). Results obtained through regression analyses for each group of data: total richness of pollinators R2= 0.32; P= 0.022, richness of hymenopterans in 2005 and 2006 R2= 0.10; P= 0.39 y R2= 0.08; P= 0.47, respectively; mean frequency of visits: R2= 0,.27; P= 0.47 y R2= 0.05; P= 0.90 for 2005 and 2006 seasons, respectively.
Con el grupo de datos tomados sobre plantas focales utilizamos una aproximación metaanalítica para evaluar tendencias en el proceso de polinización y reproducción en el gradiente de fragmentación analizado. La utilización de esta herramienta estadística permite evaluar las tendencias en la riqueza de polinizadores, polinización y reproducción, encontradas para un gran conjunto de especies de plantas. A partir de los resultados para cada una las variables analizadas, pretendemos hacer una inferencia sobre lo que estaría ocurriendo a escala de la comunidad.
Este tipo de análisis permite integrar y comparar medidas heterogéneas (por ejemplo, en número de muestras, variabilidad, escalas) generadas por distintos investigadores, en diferentes sistemas y temporadas. Para realizar un meta-análisis es necesario estandarizar los datos a través de una ecuación y se calcula lo que se denomina la "magnitud del efecto"(effect size) para cada dato primario. Luego, se realiza una estimación global de la tendencia dentro de un determinado grupo de datos, definida como la "magnitud del efecto global" para cada variable. Además, se determina si esta "magnitud global" es diferente de cero. En nuestro caso calculamos ese "efecto global" para tres variables en relación al gradiente de fragmentación.
En algunos casos, las variables medidas sobre una especie vegetal fueron obtenidas en más de un estudio, aunque sólo se incluyeron estos datos como puntos independientes si fueron tomados por distintos investigadores y en distintas temporadas. Por el contrario, si un mismo investigador repitió la medición de una misma variable en la misma especie vegetal para más de una temporada, siempre se consideró sólo el grupo de datos de la última temporada.
Los datos obtenidos para cada especie de planta fueron tomados en un gradiente de fragmentación, considerando al menos cinco fragmentos de distinto tamaño, y se analizaron con correlaciones. Por esta razón, para calcular la "magnitud del efecto" de cada variable y para cada especie vegetal se utilizó una ecuación que considera el coeficiente de correlación de Pearson (r) normalizado. Esta medida de magnitud del efecto, denominada Z transformada de Fisher, se calcula como Z = 0.5 ln[(1+r)/(1-r)] (Zar 1999), y luego se pondera al multiplicarla por la inversa de la varianza muestral w=1/var(r) de cada grupo de datos (Rosenthal 1991; Zar 1999). La media ponderada de Z (estimador de la magnitud del efecto) es definida como Z w = ∑ wizi / ∑ wi, de manera que a los valores de Z con varianzas pequeñas se les otorga mayor peso que a los valores Z con varianzas grandes. Valores positivos o negativos de Z implican relaciones directas o inversas respectivamente entre el gradiente de fragmentación y la variable evaluada. Es decir, si Z tiene valor positivo indica que a medida que disminuye el área de los fragmentos de bosque, el valor medio de la variable de respuesta disminuye (i.e., relación directa). En los casos que Z presenta valores negativos indica que a medida que disminuye el área de los fragmentos el valor medio de la variable de respuesta aumenta (i.e. relación inversa).
Para el cálculo de las magnitudes del efecto y los meta-análisis se utilizó el programa estadístico MetaWin 2.0 (Rosenberg et al. 2000). Los intervalos de confianza (IC) de las magnitudes del efecto se calcularon con procedimientos de re-muestreo como se describe en Adams y colaboradores (1997). Una determinada magnitud del efecto (Z) se considera significativamente distinta de cero si el intervalo de confianza del 95% (corregido de sesgos por el re-muestreo) no se superpone con el valor cero (Rosenberg et al. 2000). Para analizar los datos se utilizó un modelo meta-analítico de efecto al azar (Raudenbush 1994), el cual asume que los estudios incluidos en el meta-análisis difieren no sólo por error de muestreo sino también por un componente al azar en las magnitudes del efecto entre los estudios (Rosenberg et al. 2000). Si bien la aproximación convencional es utilizar modelos de efecto fijo, en la síntesis de datos ecológicos se aconseja particularmente utilizar el modelo de efecto al azar por sobre el modelo de efecto fijo, debido a que los supuestos de los modelos de efecto al azar son más probables de ser cumplidos (Gurevitch & Hedges 1999). La heterogeneidad de las magnitudes del efecto se examinó con estadísticos Q (Hedges & Olkin 1985), los cuales representan sumas de cuadrado ponderadas con una distribución χ2 y n-1 grados de libertad. La heterogeneidad total para cada variable se evaluó con Qtotal, y el valor de P obtenido para cada variable indica si la varianza entre las magnitudes de efecto de los datos (i.e., especies en nuestro caso) es mayor o no que lo esperado por azar.
RESULTADOS
No se encontró una menor riqueza y/o frecuencia de visitas promedio de los polinizadores a las flores a medida que disminuye el área de los fragmentos (Figura 1). Asimismo, tampoco se encontraron tendencias claras entre estas variables y el gradiente de fragmentación cuando se analizaron los datos disponibles para distintas temporadas. Por un lado, se encontró una relación significativa e inversa entre la riqueza de polinizadores por sitio y el área de los fragmentos y una tendencia comparable entre temporadas en el caso de los himenópteros, aunque no significativa (Figura 1 A). Por otra parte, la relación entre el área de los fragmentos y la frecuencia de visitas promedio de los himenópteros a las flores mostró diferencias en la tendencia entre las dos temporadas analizadas (Figura 1 B), aunque en ningún caso fueron significativas.
Los resultados obtenidos a través de los meta-análisis para el grupo de especies que se encontraban presentes en cinco o más sitios del gradiente de fragmentación muestran que las magnitudes de efecto global para la riqueza de polinizadores (Z= 0,12; intervalo de confianza 0.02 - 0.22) y para la producción de frutos (Z= 0.26; intervalo de confianza 0.08-0.44) fueron significativamente diferentes de cero, aunque estos valores fueron bajos (Figura 2 A, C). Por el contrario, la frecuencia de visitas de los polinizadores no mostró una magnitud del efecto global significativa (Z= 0.09; intervalo de confianza -0.08-0.26; Fig. 2 B). Los estadísticos Q no mostraron valores significativos para los meta-análisis realizados (Q total riqueza = 60.8; g.l.=58; p=0.37; Q total frecuencia de visitas = 33.7; g.l.= 33; p=0.43; Q total fructificación= 34.1; g.l.= 44; p=0.86). Es decir, la heterogeneidad de las magnitudes del efecto entre las especies consideradas en cada meta-análisis no es mayor que lo esperado por azar y por lo tanto cada grupo de especies considerado al analizar cada variable es estadísticamente homogéneo. En la figura 2 se puede observar, además, los valores correspondientes a la magnitud del efecto encontrada para cada una de esas especies con cada variable. Asimismo, es interesante señalar que varias de las especies que fueron estudiadas independientemente en distintas temporadas mostraron diferentes respuestas (i.e., algunos años la magnitud del efecto resultó positiva y en otros negativa) en relación a la disminución del área (Apéndice).


Figura 2. Magnitudes del efecto (Zr) e intervalos de confianza del 95% (IC) para especies de plantas a partir de las correlaciones entre el área de los fragmentos de bosque y (a) la riqueza de polinizadores, (b) polinización (medida como frecuencia de visita de polinizadores) y (c) reproducción (medida como producción de frutos). Cada punto representa la magnitud del efecto para una especie vegetal del Chaco sobre la que se realizaron las observaciones. El punto lleno de mayor tamaño representa el valor medio ponderado de la magnitud del efecto global de cada variable y su IC para la muestra de especies considerada. La línea de puntos denota el valor cero de la magnitud del efecto. En aquellas variables (o especies) donde se obtuvieron valores de Z (o su IC) superpuestos con el "0", la magnitud del efecto es considerada no significativo; en cambio, si los valores de Z son negativos o positivos y el intervalo de confianza no se superpone con "0", se considera que la magnitud del efecto es significativa, aunque en forma directa o inversa respectivamente.
Figure 2. Effects size (Zr) and 95% confidence intervals (IC) for plant species using correlation data between fragment areas and (a) pollinator richness, (b) pollination (measured as frequency of pollinator visists), and (c) reproduction (measured as fruit production). Each point represents the effect size for the data obtained for a particular plant species of the Chaco. The bigger filled point represents the mean value for the cumulative effect size of each variable and the IC for the considered sample of species. Dotted line indicates the zero value for the magnitude of the effect size. In those variables (or species) where Z values (or the IC) bracket the "0", the effect size is considered not significant. On the contrary, if Z values are negative or positive and if the IC does not bracket zero, then the cumulative effect size is significant in a direct or inverse form respectively.
DISCUSIÓN
Las dos aproximaciones utilizadas para evaluar tendencias en las variables relacionadas al mutualismo planta-polinizador dentro del gradiente de fragmentación considerado son interesantes de analizar, ya que muestran que los resultados obtenidos no fueron uniformes.
Por un lado, el análisis de regresión a escala de sitio mostró que no se evidencia una disminución en la riqueza de polinizadores (o de himenópteros) o en la frecuencia de visitas promedio a medida que disminuye el área de bosque como se hipotetizó a priori. Resultados publicados en estudios previos permiten especular sobre algunas razones que explicarían estos resultados que no se ajustan a la predicción planteada. Muchas especies de polinizadores tendrían la capacidad de vuelo suficiente (Rathcke & Jules 1993; Steffan-Dewenter et al. 2002 y trabajos citados allí; Westphal & Steffan-Dewenter 2006) para poder desplazarse entre sitios atravesando la matriz que rodea a los fragmentos (e.g., Jules & Shahani 2003) y de esa manera explotar los recursos florales disponibles en los distintos sitios de bosque. Se ha visto que las poblaciones locales de polinizadores tendrían la capacidad para sobrevivir y mantenerse en cada sitio, independientemente de su capacidad de vuelo, y por lo tanto no serían mayormente afectados por la disminución del área. Distintos datos de la literatura pueden considerarse en este sentido: i) se ha observado que muchos polinizadores serían muy "fieles" a los sitios en donde nidifican, (Osborne & Williams 2001 y trabajos citados allí, Goverde et al. 2002) por lo que no se desplazarían de un sitio a otro aunque tengan la capacidad de vuelo para hacerlo; por ejemplo, los abejorros del género Bombus (Hymenoptera: Apidae), que presentan una capacidad de vuelo de hasta 20 Km, permanecen en el sitio en el que nidifican incluso cuando se han eliminado casi por completo los recursos alimenticios del fragmento que habitan (Bhattacharya et al. 2003); ii) los polinizadores serían renuentes a atravesar barreras artificiales (por ejemplo, pequeños cultivos, pastizales, carreteras o ambientes muy modificados; sin embargo ver Jules & Shahani 2003), como lo observado para especies de Euglocinae (Powell & Powell 1987), Bombus (Bhattacharya et al. 2003) o lepidópteros (Lovejoy et al. 1986).
Por otro lado, cuando se analizan los resultados obtenidos a partir de datos recolectados para un conjunto de especies focales de plantas que presentan poblaciones en varios de los fragmentos estudiados, las tendencias observadas son distintas, lo cual evidencia que las respuestas de los polinizadores y las plantas frente al proceso de fragmentación son mucho más complejas que lo planteado a priori. A la vez, con esta segunda aproximación es posible desarrollar una discusión más detallada sobre algunos factores relacionados al proceso de polinización. Tenemos que la magnitud del efecto global mostró una relación directa y significativa entre el área de los fragmentos de bosque y la riqueza de polinizadores y la producción de frutos (i.e., se puede inferir una tendencia general que indica que a medida que disminuye el área de los fragmentos también disminuye la riqueza de polinizadores y la producción de frutos), aunque la relación entre el área de los fragmentos de bosque y la polinización (medida como frecuencia de visitas a las flores) no fue significativa.¿Cómo podríamos explicar estos resultados considerando además las diferencias encontradas entre las dos aproximaciones consideradas?
El análisis de la heterogeneidad en las tendencias encontradas, tanto al comparar los resultados de las dos aproximaciones utilizadas como las tendencias entre distintas especies de plantas dentro del gradiente de fragmentación, sugiere una susceptibilidad especie-específica frente a la fragmentación de hábitat. Es decir, algunas especies de plantas, y quizás de polinizadores, serían mucho más susceptibles que otras a la disminución del área de los fragmentos. Este grupo de especies de plantas sería prioritario de ser considerado en políticas de conservación del bosque Chaqueño de los alrededores de Córdoba, ya que su reproducción se vería afectadas por la constante disminución del tamaño de los sitios de bosque nativo. Algunas de las especies de plantas más afectadas serían Mandevilla pentlandiana (Apocynaceae), Acacia caven (Fabaceae), Porlieria microphylla (Zygophyllaceae), Dicliptera tweediana (Acanthaceae) y Heimia salicifolia (Lythraceae). Es interesante remarcar que no existiría un sesgo filogenético o de forma de vida en este grupo de plantas. En general son relativamente frecuentes a escala regional, por lo cual podrían utilizarse como especies de referencia para evaluar los efectos negativos del proceso de fragmentación. Por otro lado, también queda evidenciado que en otras especies de plantas el proceso de polinización o la reproducción no se ve mayormente afectado (i.e., relaciones neutras), o bien la relación de las variables con la disminución del área de los fragmentos resultaron ser inversas y significativas. Asimismo, la fructificación de algunas plantas podría estar siendo afectada por los cambios en la composición cualitativa y/o cuantitativa de las especies de polinizadores entre sitios.
A partir de los resultados encontrados queda de manifiesto que sería interesante profundizar los estudios sobre las características particulares de los polinizadores que interactúan con las especies de plantas cuya producción de frutos estaría siendo afectada negativamente al disminuir el área de los fragmentos. Es decir, empezar por dar algunas respuestas desde el punto de vista de los animales como por ejemplo ¿se trata de polinizadores que se alimentan de unas pocas especies de plantas o son polinizadores generalistas que cubren sus necesidades alimenticias a partir de un amplia variedad de plantas?, ¿es alterada la utilización estacional de los recursos florales por parte de los polinizadores ante los cambios en la diversidad de plantas relacionados con la disminución del área de los fragmentos?, ¿cuáles son los requerimientos de hábitat que presentan distintos polinizadores a la hora de reproducirse o nidificar?
Muy pocos son los aportes que se han hecho en este sentido a escala de comunidad para otros lugares del mundo, con respuestas dispares según los distintos grupos de polinizadores considerados. Se ha observado que polinizadores altamente especializados en las flores que visitan para su alimentación sufren más las consecuencias de la fragmentación que aquéllos generalistas (Steffan-Dewenter & Tscharntke 2000, 2002; Goverde et al. 2002; Steffan-Dewenter et al. 2006). En cuanto a los requerimientos de nidificación/reproducción de las especies y su relación con la fragmentación de hábitat, Steffan-Dewenter et al. (2006) compararon el grupo de abejas que nidifican en el suelo con aquéllas que lo hacen en orificios preexistentes, sin encontrar diferencias significativas entre ellos. Sin embargo, cuando se utilizaron nidos trampa se encontró que la riqueza total de especies de abejas y avispas que utilizan estos nidos disminuye con el área del fragmento, siendo el grupo más afectado el de las abejas solitarias (Morato & Campos 2000; Steffan-Dewenter 2003; Morato 2003). En resumen, dada la diversidad de respuestas encontradas hasta el momento en ambientes fragmentados para plantas y polinizadores, y también para los procesos asociados a esta interacción mutualista, es necesario realizar estudios a largo plazo que contemplen este doble punto de vista de las interacciones de polinización para poder evaluar posibles medidas efectivas de conservación frente a la creciente pérdida de ambientes naturales.
CONCLUSIONES Y PERSPECTIVAS
Los resultados obtenidos indican que la riqueza total de polinizadores no muestra una tendencia clara en relación al área de los fragmentos y que las respuestas de los animales y las plantas frente al proceso de fragmentación son mucho más complejas que lo pensado. La posibilidad de haber estudiado un gran número de especies con distintas escalas de análisis evidenció que es necesario conocer mucho más sobre otras características biológicas y ecológicas de los polinizadores y de las plantas con las que interaccionan, antes de poder generalizar las consecuencias de la fragmentación del hábitat sobre distintos procesos ecológicos. Es decir, al analizar un gran conjunto de especies de plantas se evidenciaron diferencias en las tendencias encontradas considerando la relación entre el área de los fragmentos, la riqueza de polinizadores y las variables relacionadas a la polinización y/o su reproducción.
Es necesario incrementar el número de trabajos que evalúen las interacciones planta-polinizador a escala de comunidad y que consideren distintas historias de uso de la tierra. Por ejemplo, es posible que la altísima tasa de deforestación que experimentó la región durante las últimas décadas (Gavier & Bucher 2004, Zak et al. 2004) impida evidenciar las tendencias esperadas para el actual paisaje altamente fragmentado, debido a que quizás muchas especies de polinizadores nativos ya hayan desaparecido de esta región del Chaco. Es decir, quizás muchos de los polinizadores más vulnerables a la fragmentación de hábitat (por ejemplo, los especialistas) ya no se encuentren presentes en la región, y quizás sean las especies que pudieron sobrevivir (por ejemplo, especies generalistas) las que se encuentran actualmente en la mayoría de los sitios interactuando con una gran cantidad de plantas.
Asimismo, hacen falta trabajos que consideren la trama completa de interacciones en cada uno de estos fragmentos y evalúen además algunas características ecológicas de los organismos, ya que las tendencias encontradas aquí muestran que las respuestas de los animales y las plantas frente al proceso de fragmentación son difíciles de generalizar. Una perspectiva interesante sería estudiar cómo varía la topología de las redes de interacción plantapolinizador en un gradiente de fragmentación y, en particular, si dentro de un mismo sistema es posible evidenciar las asimetrías en las interacciones especializadas y generalizadas postuladas recientemente (Ashworth et al. 2004).
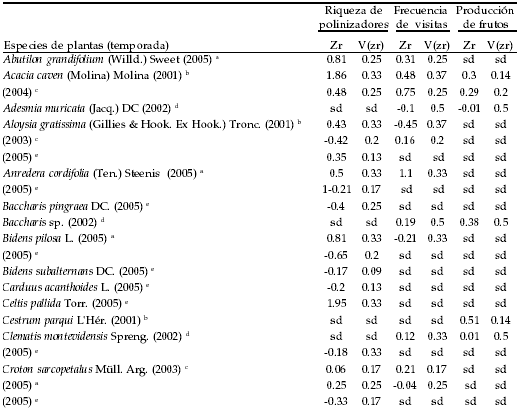
APÉNDICE
Especies vegetales estudiadas en el Chaco de Córdoba en un gradiente de fragmentación de hábitat (i.e., sitios de bosque de diferente tamaño) en diferentes temporadas. Los valores de Zr y V (zr) de cada variable corresponden a de la magnitud de efecto (Z-transformada de Fischer) y su varianza respectivamente, fueron calculadas a partir de los valores de correlación (ver detalles en M y M). Los supraíndices indican las personas que tomaron los datos .Abreviaturas: sd = sin datos.
Plant species studied in the Chaco of Córdoba within a habitat-fragmentation gradient (i.e., forest sites of a different size) during different seasons. The values of the effect size (Fisher's z-transform) and its variance (Vzr) that were calculated from the correlation data are given (for more details see M&M). Supra indexes indicate researchers who collected the data. Abbreviations: sd = no data.



AGRADECIMIENTOS
A C. Morales y N. Chacoff por la invitación a participar en un Simposio durante las XXII Reunión Argentina de Ecología y también por su estímulo para concretar la presente contribución. A C. Morales, L. Cagnolo y un revisor anónimo que con sus comentarios y sugerencias mejoraron versiones anteriores de este trabajo, a B. Caruso por su colaboración en la toma de algunos datos de campo, a M. Cabido y M. Zak por su asesoramiento en la identificación espacial de los fragmentos a través de imágenes satelitales. A los dueños y/o responsables de la administración de las tierras, especialmente los de la Estancia Santo Domingo y Agencia Córdoba Ambiente, por permitirnos el acceso a varios de los sitios para realizar los estudios, a FONCyT, CONICET y SECyT (UNC) por el apoyo financiero otorgado durante todos estos años. CT, LA, RA y LG son miembros de la Carrera del Investigador de CONICET y JA, AEF, MM y MJ son becarios de CONICET.
BIBLIOGRAFÍA
ADAMS, DC; J GUREVITCH & MS ROSENBERG. 1997. Using randomization techniques to analyze behavioural data. An. Behav. 51:733-738. [ Links ]
AGUILAR, R. 2005. Efectos de la fragmentación del hábitat sobre el éxito reproductivo de especies nativas del Bosque Chaqueño Serrano de Córdoba. Tesis Doctoral. Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba. [ Links ]
AGUILAR, R & L GALETTO. 2004. Effects of forest fragmentation on male and female reproductive success in Cestrum parqui (Solanaceae). Oecologia 138:513-520. [ Links ]
AGUILAR, R; L ASHWORTH; L GALETTO & MA AIZEN. 2006. Plant reproductive susceptibility to habitat fragmentation: review and synthesis through a meta-analysis. Ecol. Lett. 9:968-980. [ Links ]
AIZEN, MA & P FEINSINGER. 1994. Forest fragmentation, pollination, and plant reproduction in a Chaco dry forest, Argentina. Ecology 75:330-351. [ Links ]
ASHWORTH, L. 2004. Variabilidad espacio-temporal en el éxito reproductivo de plantas leñosas del Bosque Chaqueño Serrano. Tesis Doctoral, Facultad de Ciencias Exáctas, Físicas y Naturales, Universidad Nacional de Córdoba. [ Links ]
ASHWORTH, L; R AGUILAR; L GALETTO & M AIZEN. 2004. Why do pollination generalist and specialist plant species show similar reproductive susceptibility to habitat fragmentation? J. Ecol. 92:717-719. [ Links ]
BHATTACHARY, AM; RB PRIMACK & J GERWEIN. 2003. Are roads railroads barriers to bumblebee movement in a temperate suburban conservation area? Biol. Conserv. 109:37-45. [ Links ]
CANE, JH. 2001. Habitat fragmentation and native bees: a premature veredict? Conserv. Ecol. 5:3. [ Links ]
FAHRIG, L. 2003. Effects of habitat fragmentation on biodiversity. Ann. Rev. Ecol. Syst. 34:487-515. [ Links ]
FISHER, M & D MATTHIES. 1998. Effects of population size on performance in the rare plant Gentianella germanica. J. Ecol. 86:195-204. [ Links ]
GAVIER, GI & EH BUCHER. 2004. Deforestación de las Sierras Chicas de Córdoba (Argentina) en el período 1970 - 1997. Miscelánea Nº 101. Academia Nacional de Ciencias (Córdoba). [ Links ]
GOVERDE, M; K SCHWEIZER; B BAUR & A ERHARDT. 2002. Small-scale habitat fragmentation effects on pollinator behaviour: experimental evidence from the bumblebee Bombus veteranus on calcareous grasslands. Biol. Conserv. 104:293-299. [ Links ]
GUREVITCH, J & LV HEDGES. 1999. Statistical issues in ecological meta-analyses. Ecology 80:1142-1149. [ Links ]
HEDGES, LV & I OLKIN. 1985. Statistical methods for Meta-analysis. Academic Press, New York. [ Links ]
HENLE, K; DB LINDENMAYER; CR MARGULES; DA SAUNDERS & C WISSEL. 2004. Species survival in fragmented landscapes: where are we now? Biodiv. Conserv. 13:1-8. [ Links ]
HOBBS, RJ & CJ YATES. 2003. Impacts of ecosystem fragmentation on plant populations: generalizing the idiosyncratic. Austr. J. Bot. 51:471-488. [ Links ]
JACQUEMYN, H; J BUTAYE & M HERMY. 2001. Forest plant species richness in small, fragmented mixed deciduous forest patches: the role of area, time and dispersal limitation. J. Biogeogr. 28:801-812. [ Links ]
JULES, ES. 1998. Habitat fragmentation and demographic change for a common plant: Trillium in old-growth forest. Ecology 79:1645-1656. [ Links ]
JULES, ES & P SHAHANI. 2003. A broader ecological context to habitat fragmentation: Why matrix habitat is more important than we thought. J. Veg. Sci. 14:459-464. [ Links ]
KEARNS, CA; DW INOUYE & NM WASER. 1998. Endangered mutualisms: The conservation of plant-pollinator interactions. Ann. Rev. Ecol. Syst. 29:83-112. [ Links ]
LENNARTSSON, T. 2002. Extinction thresholds and disrupted plant-pollinator interactions in fragmented plant populations. Ecology 83:3060-3072. [ Links ]
LIENERT, J. 2004. Habitat fragmentation effects on fitness of plant populations - a review. J. Nature Conserv. 12:53-72. [ Links ]
LOVEJOY, TE; RO BIERREGAARD; JR RYLANDS; JR MALCOM; et al. 1986. Edge and other effects of isolation on Amazon forest fragments. Pp. 257-285 in: ME Soule (ed), Conservation Biology. Sinauer Associates, Sunderland. [ Links ]
MORALES, LC & L GALETTO. 2003. Influence of compatibility system and life form on plant reproductive success. Plant Biol. 5:1-7. [ Links ]
MORATO, EF. 2003. Biologia de Megachile (Austromegachile) orbiculata Mitchell (Hymenoptera, Megachilidae) em matas contínuas e fragmentos na Amazonia Central. En: G. A. R. Melo & A. Dos Santos (eds). Apoidea Neotropica: Homenagem aos 90 anos de Jesus Santiago Mour. Editora UNESC, Criciúma. [ Links ]
MORATO, EF & LA CAMPOS. 2000. Efeitos da fragmentacao florestal sobre vespas e abelhas solitárias em uma área da Amazonia Central. Rev. Zool. 17: 429-444. [ Links ]
NOSS, RF & AY COOPERRIDER. 1994. Saving nature´s legacy: protecting and restoring biodiversity. Island Press, Washington. [ Links ]
OSBORNE, JL & IH WILLIAMS. 2001. Site constancy of bumble bees in an experimentally patchy habitat. Agriculture, Ecosystems and Environment 83:129-141. [ Links ]
POWELL, AH & GVN POWELL. 1987. Population dynamics of male euglossine bees in Amazonian forest fragments. Biotropica 19:176-179. [ Links ]
RATHCKE, BJ & ES JULES. 1993. Habitat fragmentation and plant-pollinator interactions. Curr. Sci. 65:273-277. [ Links ]
RAUDENBUSH, SW. 1994. Random effects models. Pp. 301-321, in Cooper H & LV Hedges (eds) The handbook of research systhesis. Russell Sage foundation, New York. [ Links ]
ROSENBERG, MS; DC ADAMS & J GUREVITCH. 2000. MetaWin: Statistical Sofware for Meta-analysis. Version 2.0 Sinauer Associates, Sunderland, Massachusetts. [ Links ]
ROSENTHAL, R. 1991. Meta-analytic procedures for social research. Sage, Newbury Park, CA. [ Links ]
SAUNDERS, DA; RJ HOBBS & CR MARGULES. 1991. Biological consequences of ecosystem fragmentation: a review. Conserv. Biol.5:18-32. [ Links ]
STEFFAN-DEWENTER, I. 2003. Importance of habitat area and landscape context for species richness of bees and wasps in fragmented orchard meadows. Conserv. Biol. 17:1036-1044. [ Links ]
STEFFAN-DEWENTER, I & T TSCHARNTKE. 1999. Effects of habitat isolation on pollinator communities and seed set. Oecologia 121:432-440. [ Links ]
STEFFAN-DEWENTER, I & T TSCHARNTKE. 2000. Butterfly community structure in fragmented habitats. Ecol. Lett. 3:449-456. [ Links ]
STEFFAN-DEWENTER, I & T TSCHARNTKE. 2002. Insect communities and biotic interactions on fragmented calcareous grasslands-a mini review. Biol. Conserv. 104:275-284. [ Links ]
STEFFAN-DEWENTER, I; U MÜNZENBERG; C BÜRGER; C THIES & T TSCHARNTKE. 2002. Scale-dependent effects of landscape context on three pollinator guilds. Ecology 83:1421-1432. [ Links ]
STEFFAN-DEWENTER, I; A KLEIN; V GAEBELE; T ALFERT & T TSCHARNTKE. 2006. Bee diversity and plant-pollinator interactions in fragmented landscapes. Pp.387-405 in: Waser NM & J Ollerton (eds). Plant-Pollinator Interactions from specialization to generalization. Oxford University Press, Oxford. [ Links ]
TORRES, C. 2003. Ecología reproductiva de Asteraceae nativas de Argentina central. Tesis Doctoral. Facultad de Ciencias Exáctas, Físicas y Naturales, Universidad Nacional de Córdoba. [ Links ]
VAN DYCK, H & E MATTHYSEN. 1999. Habitat fragmentation and insect flight: a changing "design" in a changing landscape? Trends Ecol. Evol. 14:172-174. [ Links ]
WESTPHAL, C & I STEFFAN-DEWENTER. 2006. Bumblebees experience landscapes at different spatial scales: possible implications for coexistence. Oecologia 149:289-300. [ Links ]
WILSON, EO. 1992. The diversity of life. Harvard University Press, Cambridge, Massachusetts. [ Links ]
YAO, J; RD HOLT; PM RICH & WS MARSHALL. 1999. Woody plant colonization in an experimentally fragmented landscape. Ecography 22:715-728. [ Links ]
ZAK, M; MR CABIDO & J HODGSON. 2004. Do subtropical seasonal forests in the Gran Chaco, Argentina, have a future? Biol. Conserv. 120:589-598. [ Links ]
ZAR, JH. 1999. Biostatistical analysis. Prentice Hall, New Jersey. [ Links ]

