










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.17 n.1 Córdoba ene./jun. 2007
TRABAJOS ORIGINALES
Efectos de la frecuencia de corte y la fertilización nitrogenada sobre la estructura de una población implantada de Festuca arundinacea Schreb.
Andrés G Rolhauser1, Sergio Cepeda2, Pablo H Maseda1, José L Rotundo1, Ana M Srur3, Roberto J Fernández1, Claudio M Ghersa1, Rolando JC León1, Susana B Perelman1, William B Batista1 & Martín R Aguiar1,*
1. Instituto de Investigaciones Fisiológicas y Ecológicas Vinculadas a la Agricultura (IFEVA), Facultad de Agronomía de la Universidad de Buenos Aires (FAUBA) - CONICET, Buenos Aires, Argentina.
2. Instituto Nacional de Tecnología Agropecuaria (INTA), Estación Experimental Agropecuaria Pergamino, Pergamino, Argentina.
3. Instituto Argentino de Nivología, Glaciología y Ciencias Ambientales (IANIGLA), Mendoza, Argentina.
* IFEVA, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453 (C1417DSE), Buenos Aires, Argentina. Email: aguiar@ifeva.edu.ar
Recibido: 2 de enero de 2006;
Fin de arbitraje: 16 de mayo de 2006;
Revisión recibida: 11 de diciembre de 2006;
Segunda revisión recibida: 5 de marzo de 2007;
Aceptado: 5 de marzo de 2007.
Este trabajo fue aceptado durante el proceso editorial de Marcelo Cassini.
RESUMEN. La estructura y la dinámica de las poblaciones vegetales de los sistemas pastoriles están principalmente controladas por la competencia intra e interespecífica y por la herbivoría, en el marco impuesto por el ambiente edáfico y el clima. Las defoliaciones producidas por los herbívoros impactan directamente sobre las poblaciones vegetales, pero también indirectamente, mediante el control de la intensidad de la competencia. En este trabajo evaluamos los efectos de la frecuencia de defoliaciones mecánicas y de la fertilización con nitrógeno sobre la estructura poblacional de Festuca arundinacea Schreb. sembrada en una pastura mixta y discutimos los efectos de estas prácticas agronómicas sobre la competencia intraespecífica. Sobre una pastura compuesta por F. arundinacea y Lotus glaber Mill. instalamos un experimento factorial que combina dos niveles de frecuencia de cortes (uno y cuatro cortes anuales) y dos de fertilización nitrogenada (0 y 50 kg N/ha/año) en cuatro bloques. Los cortes frecuentes resultaron en mayor densidad y menor jerarquización de tamaños y no modificaron la cobertura total en comparación con los cortes poco frecuentes. En cambio, la fertilización nitrogenada disminuyó la densidad, generó mayor jerarquización de tamaños y aumentó la cobertura total. El log10 del tamaño promedio estuvo negativamente asociado con el log10 de la densidad; la pendiente estimada de la relación lineal fue más cercana a –3/2 para las parcelas fertilizadas que para las no fertilizadas. El apiñamiento de individuos (medido en una escala de 0.25 m2) disminuyó significativamente con el aumento del tamaño individual. Nuestros resultados sugieren que la dinámica de F. arundinacea estuvo fuertemente controlada por la competencia intraespecífica en condiciones de baja frecuencia de disturbios y alta disponibilidad de nitrógeno. Además, indican que la adición de un recurso posiblemente limitante puede intensificar la competencia intraespecífica en lugar de relajarla.
Palabras clave: Competencia intraespecífica; Disturbio; Dinámica poblacional; Autorraleo; Sistemas pastoriles; Pastura; Festuca alta.
ABSTRACT. Effects of mowing frequency and nitrogen fertilization on the structure of a Festuca arundinacea Schreb. population: Structure and dynamics of plant populations in pastoral systems are mainly controlled by inter and intraspecific competition and herbivory within the constraints imposed by soil characteristics and climate. Defoliations caused by herbivores have a direct impact on plant populations, but also an indirect effect through changes in the intensity of competition. We evaluated the effects of mechanic defoliations and nitrogen fertilization on the structure of a Festuca arundinacea Schreb. population in a mixed pasture, and discussed the effects of these two agronomic tools on the intensity of intraspecific competition. A four-block factorial experiment combining two regimes of mowing frequency (1 and 4 mowings/year) and two levels of nitrogen fertilization (0 and 50 kg N/ha/year), was installed on a mixed pasture composed of F. arundinacea and Lotus glaber Mill. Data was obtained from eight subplots randomly arranged inside each plot. High frequency mowed plots had higher density and lower size asymmetry than the low frequency ones, whereas total cover did not differ between treatments. On the contrary, nitrogen fertilization diminished population density, increased size asymmetry and augmented total cover. Overall, there was a negative association between log10 mean size and log10 density; the estimated slope of the linear relation was closer to –3/2 in fertilized than in nonfertilized plots. Finally, crowding decreased significantly with plant size. These results suggest that the dynamics of F. arundinacea in the experimental plots were strongly controlled by intraspecific competition when nitrogen availability was high and mowing frequency was low. Lack of fertilization and frequent mows would limit individual growth and hence intensity of competition. Furthermore, the intensity of competition seems to have been more closely related to nitrogen availability than to mowing frequency. Additional data showed that the abundance of companion species was precluded in fertilized plots and in low frequency mowed plots, suggesting that F. arundinacea is a highly competitive species, which maximizes vegetative growth in productive, relatively undisturbed conditions. Overall, our results indicate that resource supply could intensify plant competition instead of relax it.
Keywords: Intraspecific competition; Disturbance; Population dynamics; Self-thinning; Pastoral systems; Pasture; Tall fescue.
INTRODUCCIÓN
Los sistemas pastoriles, naturales, semi-naturales o implantados, están conformados por un conjunto de especies predominantemente herbáceas que interactúan entre sí y con los herbívoros, y presentan diferentes tipos de respuesta en tiempos ecológico y evolutivo (Aarssen 1983; Coughenour 1985; Mehrhoff & Turkington 1990). Las principales interacciones que modulan la estructura y la dinámica de las poblaciones vegetales de los sistemas pastoriles son la herbivoría junto con la competencia intraespecífica y la interespecífica (McNaughton 1979; Tilman 1982). En general, es común que el estudio de estos sistemas se realice en la escala de comunidad (e.g. análisis de la composición florística) o desde una perspectiva ecosistémica (e.g. análisis de la producción de biomasa). Estudios detallados de las respuestas de especies individuales (e.g. de las respuestas poblacionales a la herbivoría o a la competencia) son menos frecuentes, si bien las respuestas poblacionales explican las respuestas a nivel de comunidad y ecosistema. En aquellas oportunidades en que se estudia una especie, generalmente se documenta su producción de biomasa mientras que la caracterización de la estructura poblacional (la densidad, la estructura de tamaños, la organización espacial) es dejada de lado. En este trabajo evaluamos la estructura poblacional de Festuca arundinacea Schreb. como respuesta a la defoliación y a la fertilización nitrogenada, dos prácticas agronómicas comunes en sistemas pastoriles.
Festuca arundinacea es una gramínea C3 utilizada como forrajera invernal en establecimientos ganaderos de todo el mundo (Gibson & Newman 2001). En ausencia de ganado, Festuca domina las pasturas y también invade y domina pastizales en los que no ha sido sembrada Clay & Holah 1999; Gibson & Newman 2001). En pastizales de la Depresión del Salado (Buenos Aires, Argentina) clausurados al pastoreo también ha demostrado un comportamiento invasor y dominante (E. Chaneton, IFEVA, com. pers.). Esta dominancia es un indicador de que, en ausencia de disturbios, su dinámica poblacional debería estar controlada por la competencia intraespecífica más que por la interespecífica (Tilman 1982; Aguiar et al. 2001).
En las poblaciones vegetales compuestas por individuos coetáneos (i.e. emergidos al mismo tiempo), la competencia intraespecífica tiene dos efectos encadenados (Silvertown & Lovett-Doust 1993). En una primera etapa, promueve la formación de jerarquías de tamaños, es decir, una estructura de tamaños heterogénea, con muchos individuos pequeños y pocos grandes. Esta jerarquía puede originarse por pequeñas diferencias de crecimiento inicial (debidas a diferencias individuales o del microambiente) que luego se retroalimentan positivamente y aumentan progresivamente las diferencias de tamaños (Harper 1977). En una segunda etapa, si la competencia persiste, comienza la muerte de los individuos de menor tamaño, proceso conocido como autorraleo. Como consecuencia, para un rango relativamente amplio de densidades, la asociación entre el tamaño individual y la densidad es frecuentemente negativa, y en forma repetida se ha medido que la relación lineal entre ambas tiene un valor aproximado a –3/2 cuando son expresadas en escala logarítmica con base 10 (Yoda et al. 1963; Harper 1977; White 1985). Este patrón se conoce con el nombre de "regla de la pendiente –3/2" (Yoda et al. 1963; Harper 1977). Como resultado del autorraleo, los individuos sobrevivientes se ubican en un arreglo espacial regular, el cual minimizaría la competencia entre individuos (Pielou 1969; Kenkel 1988). En el caso de poblaciones con jerarquías de edades o de tamaños (e.g. luego del autorraleo), la competencia intraespecífica también puede controlar los procesos de establecimiento y crecimiento de nuevos individuos. En estos casos, los individuos de mayor tamaño ocupan el espacio en forma amplia y monopolizan el uso de recursos del suelo y luz, lo cual puede restringir la disponibilidad de sitios favorables para la regeneración ("sitios seguros" sensu Harper 1977). En estas situaciones, disturbios tales como la herbivoría suelen promover el establecimiento de nuevos individuos porque eliminan o reducen la biomasa de los individuos dominantes (Harper 1977; Tilman 1982; Silvertown & Lovett-Doust 1993).
Nuestra hipótesis general propone que la competencia intraespecífica es un control importante de la dinámica de F. arundinacea y por lo tanto afecta la estructura y organización de la población. Entonces, poblaciones sometidas a defoliaciones frecuentes deberían presentar menor intensidad de competencia intraespecífica, pues las plantas tienen bajas tasas de acumulación de biomasa, mientras que poblaciones con defoliaciones poco frecuentes, pero fertilizadas, deberían presentar mayores tasas de acumulación de biomasa y, por lo tanto, una mayor importancia de la competencia intraespecífica. Para examinar nuestra hipótesis describimos y evaluamos la estructura poblacional de F. arundinacea en parcelas que han sido sometidas a regímenes de defoliaciones y niveles de fertilización nitrogenada contrastantes.
MATERIALES Y MÉTODOS
Diseño del experimento
Este estudio se realizó utilizando un experimento de larga duración instalado por la Cátedra de Ecología de la FAUBA en el campo experimental de esta Facultad (Ciudad de Buenos Aires; 34º 35´ Sur, 58º 22´ Oeste). El clima en la región es templado-cálido, con precipitación media anual de 1215 mm y temperatura media anual de 17.7 ºC (Servicio Meteorológico Nacional Argentino, com. pública). El suelo del área experimental es un Argiudol con características vérticas (Mirta González, Cátedra de Edafología FAUBA, com. pers.). En abril de 2000 se sembró una superficie de 0.2 ha con una mezcla de F. arundinacea cultivar El Palenque (en adelante Festuca) y Lotus glaber Mill. cultivar Toba (en adelante Lotus). La siembra se realizó al voleo con densidades de 7 kg/ha en el caso de Festuca y de 4 kg/ha en el de Lotus. En octubre de 2001, se consideró terminada la etapa de establecimiento de la pastura y se realizó la delimitación de cuatro bloques de aproximadamente 250 m² cada uno. Los bloques controlan dos fuentes de variación ambiental: la topografía y la presencia de árboles en uno de los lados del área. El diseño en bloques permitió alcanzar una razonable homogeneidad florística y topográfica dentro de cada bloque. En cada bloque se delimitaron cuatro parcelas de 49 m² (7 m x 7 m) a las que se asignaron al azar cuatro tratamientos (16 parcelas en total, cuatro bloques x cuatro tratamientos), los cuales comenzaron a aplicarse regularmente desde ese momento. Tales tratamientos surgen de la combinación de dos factores con dos niveles cada uno: frecuencia de corte (uno y cuatro cortes anuales) y fertilización nitrogenada (0 y 50 kg de N/ha/año [N- y N+, respectivamente]). Los cortes de la vegetación se realizaron mecánicamente a una altura de 5 cm. En las parcelas asignadas al tratamiento con fertilización se aplicó nitrato de amonio en una dosis equivalente a 50 kg de N/ha/año. Para obtener más información acerca de este experimento de larga duración puede verse: http://www.agro.uba.ar/carreras/agronomia/materias/ecologia/practico/index.htm.
Al momento de realizar las mediciones (septiembre de 2003), las parcelas estaban claramente dominadas por Festuca. Mediciones realizadas en mayo de 2003 indicaban que la frecuencia de Festuca promedio de los tratamientos se encontraba entre 90 y 94%. En cambio, la frecuencia de Lotus (la otra especie sembrada) era en promedio ~5%, excepto para el tratamiento con cuatro cortes anuales y sin fertilización, donde trepaba a 40% (M. R. Aguiar, datos no publ.). Además, la cobertura aérea de especies espontáneas e invasoras era muy baja (A. G. Rolhauser, obs. pers.), lo cual, sumado a la relativa escasez de Lotus, le daba a la pastura el aspecto de un monocultivo de Festuca.
Colección de datos
En septiembre de 2003 dispusimos al azar en cada una de las 16 parcelas ocho subparcelas de 50 x 50 cm y dentro de ellas contamos los individuos de Festuca. El tamaño de las subparcelas fue suficiente para incluir a los individuos más grandes de Festuca, que rara vez excedían los 50 cm de diámetro basal (A. G. Rolhauser, obs. pers.). Basándonos en observaciones de campo de la morfología de las plantas (especialmente del largo de rizomas), decidimos contar como un individuo a todo aquel macollo o grupo de macollos separado a más de 5 cm de su vecino más cercano. A cada individuo encontrado en las subparcelas le medimos, a nivel de la superficie del suelo, los diámetros basales mayor y menor. Con los diámetros basales calculamos el tamaño individual (i.e. la cobertura basal individual) como π [(ØM + Øm)/4]2, donde ØM y Øm son respectivamente los diámetros mayor y menor.
Análisis de datos
Densidad, cobertura basal y tamaño promedio. Analizamos los datos de densidad, de cobertura basal total y de tamaño individual promedio (i.e. la cobertura individual promedio) promedio) mediante análisis de la varianza (ANVA) bifactorial con bloques utilizando a las parcelas como unidades experimentales. Pusimos a prueba los supuestos de normalidad y homocedasticidad de los errores y, cuando fue necesario para su cumplimiento, transformamos los datos (Devore 2003). Comparamos a posteriori los promedios de los tratamientos mediante el método de Tukey (Devore 2003). Examinamos la relación entre el tamaño individual y la densidad mediante modelos de regresión lineal simple. Para este análisis consideramos al tamaño individual como variable respuesta y a la densidad como variable predictora, ambas transformadas por log10. Esta forma de análisis nos permitió comparar nuestros resultados con la literatura. Construimos un modelo de regresión para cada tratamiento y utilizamos como unidad experimental a cada subparcela, i.e. consideramos una escala de percepción de 50 x 50 cm. La ausencia de efecto significativo de los bloques sobre el tamaño individual (ordenadas) y la densidad (abscisas) (i.e. los valores de los distintos bloques comparten rangos similares en ambos ejes) nos permitió agrupar en un solo conjunto las 32 observaciones correspondientes a cada tratamiento. Los cuatro conjuntos de datos cumplieron con los supuestos de normalidad y homocedasticidad.
Estructura de tamaños. Comparamos las distribuciones de frecuencias acumuladas de tamaños de los distintos tratamientos mediante la prueba de Kolmogorov-Smirnov (Devore 2003). Debido a que este análisis involucra comparaciones múltiples entre tratamientos, corregimos la tasa de error de tipo I mediante el método de Bonferroni (αindividual = αglobal / k; donde k es el número de comparaciones; Devore 2003). En nuestro caso, el número de comparaciones k fue 6.
Patrón espacial. Delimitamos cinco clases de tamaños (cm²): 0 a 2, >2 a 8, >8 a 32, >32 a 128 y >128. Calculamos el cociente entre la varianza y la media de la densidad de individuos en cada una de las cinco clases de tamaño para cada parcela (V/m), para lo cual utilizamos las subparcelas como unidades experimentales. El cociente V/m es un estimador del patrón espacial de los individuos: los valores superiores a la unidad indican apiñamiento y los menores una distribución regular (Pielou 1969). Evaluamos los efectos de los tratamientos sobre el cociente V/m por medio de un ANVA con bloques y tres factores (fertilización, frecuencia de corte y clase de tamaño), considerando a las parcelas como unidades experimentales. Realizamos pruebas t de Student unilaterales (Devore 2003) para comparar los cocientes V/m promedio de cada tratamiento con la unidad. Utilizamos pruebas t unilaterales izquierdas para los cocientes V/m promedio mayores a la unidad y derechas para los menores a la unidad. Todos los análisis fueron realizados mediante InfoStat (InfoStat 2002).
RESULTADOS
Densidad, cobertura basal y tamaño individual promedio
La densidad de Festuca fue significativamente mayor (aproximadamente el doble) en las parcelas con cuatro cortes anuales que en aquellas con un único corte anual (Figura 1a). Además, fue significativamente menor en las parcelas fertilizadas que en las no fertilizadas cuando se realizaron cuatro cortes pero no cuando se realizó uno solo (Figura 1a). La cobertura basal aumentó con la fertilización nitrogenada pero no difirió significativamente entre tratamientos de corte (Figura 1b). El tamaño promedio de los individuos fue menor en las parcelas con cuatro cortes y mayor en las fertilizadas; ambos efectos fueron aditivos, i.e. no hubo interacción entre factores (Figura 1c). El log10 del tamaño promedio estuvo negativamente asociado con el log10 de la densidad en los cuatro tratamientos (Figura 2). Esta relación fue más ajustada (de acuerdo al valor de r2) y cercana al valor de –3/2 (i.e. -1.5) en las parcelas fertilizadas que en las no fertilizadas (Figura 2).
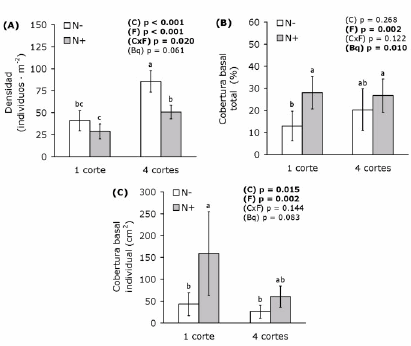
Figura 1. Densidad poblacional (A), cobertura basal total (B) y tamaño individual (C) promedios de F. arundinacea para las combinaciones de dos frecuencias de corte y dos niveles de fertilización nitrogenada (n=4 para cada tratamiento). Se presentan los valores p de cada factor según un ANVA bifactorial con bloques (C: frecuencia de cortes; F: fertilización nitrogenada; C x F: interacción). En negritas se resaltan los valores significativos al 5 %. Letras distintas indican diferencias significativas según Tukey (p ≤ 0.05) y las barras verticales indican el intervalo de confianza (95%). N-: sin aplicación de nitrógeno; N+: con aplicación de nitrógeno; Bq: bloque.
Figure 1. Mean population density (A), mean total basal cover (B) and mean plant size (C) of F. arundinacea for the combinations of two mowing frequencies and two levels of nitrogen fertilization (n=4 for each treatment). The p values of each factor according to bifactorial ANVA with blocks are presented (C: mowing frequency; F: nitrogen fertilization; C x F: interaction). Significant values at 5% are shown in bold. Different letters indicate significant differences according to Tukey (p ≤ 0.05) and vertical bars are confidence intervals (95%). N-: without nitrogen fertilization; N+: with nitrogen fertilization; Bq: block.
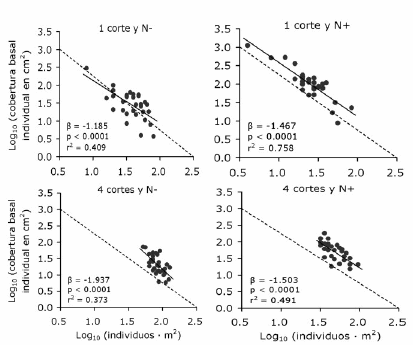
Figura 2. Relación entre los log10 del tamaño (cobertura basal) individual y de la densidad de F. arundinacea para las combinaciones de dos frecuencias de corte y dos niveles de fertilización nitrogenada. Cada punto corresponde a una subparcela (n=32 para cada tratamiento). Se presentan la pendiente estimada (β) y los valores p y r2 correspondientes al análisis de regresión lineal. Se colocó en cada gráfico una recta discontinua con pendiente –3/2 (i.e. -1.5). Las rectas de regresión fueron ligeramente extrapoladas para facilitar la comparación con las rectas discontinuas. N-: sin aplicación de nitrógeno; N+: con aplicación de nitrógeno.
Figure 2. Relationship between mean plant size (basal cover) and population density of F. arundinacea for the combinations of two mowing frequencies and two levels of nitrogen fertilization, both variables log10 transformed. Each point corresponds to a subplot (n=32 for each treatment). The estimated slope (β) and the corresponding p and r2 values according to a linear regression analysis are presented. A dotted line showing -3/2 (i.e. -1.5) slope was placed in each graphic. Regression lines were slightly extrapolated to facilitate the comparison with dotted lines. N-: without nitrogen fertilization; N+ : with nitrogen fertilization.
Estructura de tamaños
El tamaño de los individuos de Festuca (medido en términos de cobertura basal) varió desde 0.2 cm² (0.5 cm de diámetro promedio, correspondiente a un individuo de un solo tallo) hasta 1385 cm² (42 cm de diámetro promedio). En general, las poblaciones de Festuca presentaron alta proporción de individuos pequeños y muy pocos individuos grandes (Figura 3). Sin embargo, todas las comparaciones entre distribuciones acumuladas del tamaño individual mostraron diferencias significativas. Las poblaciones de Festuca presentaron la mayor jerarquización (i.e. mayores proporciones de individuos grandes y de individuos pequeños) cuando se fertilizó y la frecuencia de cortes fue baja ("1 corte y N+") (Figura 3). La distribución acumulada del tratamiento "1 corte y N-" se apartó en mayor medida de la del tratamiento "1 corte y N+" que la observada en el tratamiento "4 cortes y N+" (Figura 3). Esto indica que, con relación al tratamiento "1 corte y N+", el grado de jerarquización decreció en mayor medida cuando no se fertilizó ("1 corte y N-") que cuando la frecuencia de cortes fue alta ("4 cortes y N+"). Las parcelas con "4 cortes y N-" presentaron una estructura poco jerarquizada, con individuos muy pequeños en todo el gradiente de tamaños (Figura 3).
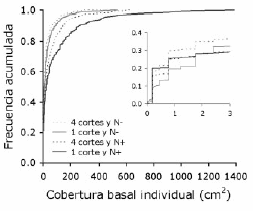
Figura 3. Distribuciones de frecuencias acumuladas del tamaño (cobertura basal) individual de F. arundinacea para las combinaciones de dos frecuencias de corte y dos niveles de fertilización nitrogenada. Todas las distribuciones difirieron significativamente de las restantes con αglobal = 0.05 según pruebas de Kolmogorov-Smirnov. La figura interior muestra las distribuciones de frecuencias para el rango de tamaños 0-3 cm2. N-: sin aplicación de nitrógeno; N+: con aplicación de nitrógeno.
Figure 3. Cumulative frequency distributions of individual plant size (basal cover) of F. arundinacea for the combinations of two mowing frequencies and two levels of nitrogen fertilization. All distributions differed significantly from the rest with αoverall = 0.05 according to Kolmogorov-Smirnov tests. Frequency distributions for the 0-3 cm2 size range are shown in the inside figure. N-: without nitrogen fertilization; N+: with nitrogen fertilization.
Patrón espacial
Ni la fertilización ni el corte modificaron significativamente el cociente entre la varianza y la media de la densidad (V/m), en cambio encontramos un efecto significativo del tamaño, el cual no interactuó con ninguno de los tratamiento (Figura 4). El cociente V/m para plantas de mayor tamaño fue significativamente menor a la unidad en los tratamientos con un corte anual (indicador de patrón regular) (Figura 4). En cambio, para las plantas más pequeñas el cociente fue significativamente mayor a 1 (indicador de patrón agrupado) (Figura 4).
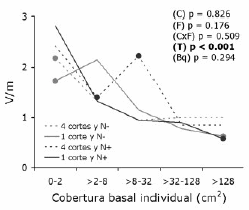
Figura 4. Cociente entre la varianza y la media de la densidad poblacional (V/m) de F. arundinacea en función del tamaño (cobertura basal) individual para las combinaciones de dos frecuencias de corte y dos niveles de fertilización nitrogenada (n=4 para cada tratamiento). El cociente V/m puede utilizarse como indicador de patrón espacial: los valores superiores a la unidad indican apiñamiento y los inferiores repulsión. Se presentan los valores p de cada factor según un ANVA con tres factores y bloques (C: frecuencia de cortes; F: fertilización nitrogenada; C x F: interacción; T: cobertura basal individual) y en negritas se resaltan los valores significativos al 5 %. Ninguna de las interacciones del tamaño individual con los tratamientos fue significativa (p<0.5). Los símbolos llenos sobre las líneas indican los valores significativamente distintos de la unidad con p ≤ 0.05 según pruebas t de Student. N-: sin aplicación de nitrógeno; N+: con aplicación de nitrógeno; Bq: bloque.
Figure 4. Variance/mean ratio of population density (V/m) of F. arundinacea as a function of individual plant size (basal cover) for the combinations of two mowing frequencies and two levels of nitrogen fertilization (n=4 for each treatment). The V/m ratio can be used as indicator of spatial pattern: values higher than one indicate crowding and lower values indicate repulsion. The p values of each factor according to ANVA with tree factors and blocks are presented (C: mowing frequency; F: nitrogen fertilization; C x F: interaction; T: individual basal cover) and significant values at 5% are shown in bold. No plant sizetreatment interaction was significant (p<0.5). Filled symbols on the lines show significant differences from one with p ≤ 0.05 according to Student´s t tests. N-: without nitrogen fertilization; N+: with nitrogen fertilization; Bq: block.
DISCUSIÓN
Nuestros resultados muestran que el tamaño promedio fue máximo y la densidad de individuos mínima con baja frecuencia de cortes y fertilización nitrogenada. Contrariamente, el tamaño promedio fue mínimo y la densidad de individuos máxima con alta frecuencia de cortes sin fertilización. Interpretamos que en nuestro experimento existen dos situaciones ambientales extremas que determinan dos niveles extremos de competencia intraespecífica en la población de Festuca. Por un lado, la baja frecuencia de cortes (disturbios) y la mayor oferta de nitrógeno (disponibilidad de recursos del suelo) habrían determinado una alta intensidad de competencia y, en consecuencia, la supervivencia de solo unos pocos individuos de gran porte. En el otro extremo, la alta frecuencia de corte y la no fertilización habrían limitado el crecimiento de los individuos y su capacidad de absorber recursos del suelo y luz, lo cual a su vez habría limitado el autorraleo y favorecido la supervivencia de una mayor cantidad de individuos. Nuestros resultados son entonces consistentes con la hipótesis de que los disturbios relajan la competencia entre individuos (Harper 1977; Tilman 1982; Silvertown y Lovett-Doust 1993). Además, ponen de manifiesto que la adición de un recurso posiblemente limitante (que para gramíneas es usualmente el nitrógeno) puede intensificar la competencia en lugar de relajarla.
Para explorar el grado en que las poblaciones de Festuca se estructuraban y autorraleaban debido a la competencia intraespecífica luego de la implantación, estudiamos las estructuras de tamaños y las relaciones entre el tamaño individual y la densidad de individuos (Harper 1977; Silvertown y Lovett-Doust 1993). De acuerdo con nuestro análisis de la estructura de tamaños, la combinación de fertilización con nitrógeno y baja frecuencia de cortes habría estructurado fuertemente a la población de Festuca, promoviendo la formación de matas de gran tamaño (las más grandes en nuestro experimento) y relativamente muchas de tamaño muy pequeño. Con relación a esta situación ambiental (fertilización con nitrógeno y baja frecuencia de cortes), la estructura de tamaños tendió en mayor medida hacia una estructura más equilibrada (o con menos jerarquías) cuando no se fertilizó que cuando se cortó el tapiz vegetal con mayor frecuencia. Esto sugiere que una mayor frecuencia de cortes fue menos efectiva para relajar la competencia intraespecífica que la no fertilización. Por otro lado, de acuerdo con la llamada regla de la pendiente –3/2, el tamaño promedio de las plantas que crecen en un stand esta negativamente relacionado con la densidad cuando la cobertura del tapiz vegetal es cercana al 100% (Yoda et al. 1963, Harper 1977). En nuestro estudio, las parcelas fertilizadas, independientemente de la frecuencia de corte, presentaron mayor cobertura basal (potencialmente proporcional a la cobertura del tapiz vegetal) y reflejaron un mayor ajuste a la pendiente de –3/2. Según este patrón y el observado en las estructuras de tamaños, el autorraleo habría sido un proceso especialmente importante en las parcelas fertilizadas, al menos en los primeros dos años de aplicación de los tratamientos de fertilización y corte. El hecho de que la frecuencia de cortes no afectó la cobertura basal ni el ajuste a la pendiente de –3/2 (y fue menos importante en la estructuración de las poblaciones) sugiere que aún la frecuencia de cuatro cortes no fue suficiente para suprimir o atemperar el efecto promotor de la fertilización sobre la competencia intraespecífica.
La organización espacial de la población el resultado, pero también es el determinante, de los procesos de una población (Aguiar al. 2005). Nuestros resultados indican que, en una escala de percepción de 0.25 m2 (nuestras subparcelas), ni el corte ni la fertilización modificaron la organización espacial. En cambio, encontramos que los individuos más chicos (<5 cm²) tienen un patrón de distribución diferente al de los individuos más grandes (320 cm²): mientras los primeros tienden crecer agrupados, los segundos lo hacen de forma regular. El crecimiento apiñado puede ser un indicador de la concentración de individuos en sitios favorables para la regeneración (Harper 1977). Al mismo tiempo podría suponerse que la presencia de agrupamientos puede ser un indicador de que los procesos de autorraleo aún no se han intensificado. En cambio, un patrón regular indica que los procesos de autorraleo han actuado y han quedado individuos grandes equi-espaciados (e.g. Pielou 1969; Kenkel 1988). Nosotros interpretamos que los tratamientos de fertilización y corte probablemente no modificaron la distribución de micrositios de regeneración y, si bien afectaron la densidad y la estructura de tamaños, no modificaron sensiblemente la organización de los individuos de mayor tamaño.
Impacto en la comunidad vegetal e implicancias para el manejo de pastizales
Nuestros resultados sugieren que la competencia intraespecífica es un control importante de la dinámica poblacional de Festuca, lo cual es esperable para una población dominante (Tilman 1992; Aguiar et al. 2001). Esto es consistente con resultados obtenidos en otros ecosistemas, donde Festuca tiene un comportamiento dominante en la comunidades en que se establece (Clay & Holah 1999; Gibson & Newman 2001). Según los patrones estructurales que hemos mostrado, la intensidad de la competencia intraespecífica fue mínima con alta frecuencia de cortes y no fertilización. Precisamente en estas condiciones se observaron las mayores abundancias de L. glaber Mill. y otras especies espontáneas e invasoras (Cátedra de Ecología, FAUBA, datos no publ.). Esto indica que la abundancia de otras especies vegetales en nuestro sistema de estudio dependería de la capacidad competitiva de los individuos de Festuca, la cual a su vez podría ser controlada manipulando los niveles de nitrógeno en el suelo y la frecuencia de disturbios. En este sentido, el pastoreo (la principal herramienta de manejo en los establecimientos ganaderos argentinos) y la fertilización nitrogenada podrían utilizarse en forma diferencial dependiendo de los objetivos de manejo del pastizal. Si el objetivo es mantener la alta productividad de Festuca, una intensidad de pastoreo relativamente baja evitaría el "envejecimiento" de la pastura, i.e. la invasión de especies no deseadas. En cambio, si el objetivo fuese restaurar pastizales dominados por Festuca, una intensidad alta crearía oportunidades para establecimiento y recolonización de especies nativas. A su vez, la fertilización nitrogenada se presenta como una herramienta para el manejo de Festuca antagónica al pastoreo. Un alto nivel de fertilización tendría efectos análogos a una baja intensidad de pastoreo, de manera que se evitaría el envejecimiento de la pastura. En cambio, el cese de la fertilización podría favorecer, tal como una alta intensidad de pastoreo, la recuperación del pastizal natural.
AGRADECIMIENTOS
Este trabajo formó parte del curso "Ecología de poblaciones vegetales" dictado por MRA y WBB en la Escuela Para Graduados "Alberto Soriano" de la FAUBA (EPG). Agradecemos a la EPG y al personal de la Cátedra de Ecología de la FAUBA por su ayuda en diferentes etapas del trabajo. Los comentarios de dos revisores anónimos aumentaron sensiblemente la calidad del manuscrito. La pastura estudiada fue implantada gracias a la desinteresada ayuda de H. Aquino, quien aportó equipos, materiales y tiempo.
BIBLIOGRAFÍA
AARSSEN, LW. 1983. Ecological combining ability and competitive combining ability in plants: toward a general evolutionary theory of coexistence in systems of competition. Am. Nat. 122: 707-731. [ Links ]
AGUIAR, MR; WK LAUENROTH & DP PETERS. 2001. Intensity of intra- and interspecific competition in coexisting shortgrass species. J. Ecol. 89:40-47. [ Links ]
AGUIAR, MR; PA CIPRIOTTI & T WIEGAND. 2005. Procesos poblacionales denso-dependientes y la generación de patrones de heterogeneidad dentro de la comunidad. Pp. 59-80 en: M Oesterheld, MR Aguiar, CM Ghersa & JM Paruelo (ed.). La heterogeneidad de la vegetación de los agroecosistemas. Editorial Facultad de Agronomía, Buenos Aires. [ Links ]
CLAY, K & J HOLAH. 1999. Fungal endophyte symbiosis and plant diversity in successional fields. Science 285:1742-1744. [ Links ]
COUGHENOUR, MB. 1985. Graminoid responses to grazing by large herbivores: adaptations, exaptations, and interacting processes. Annals of the Missouri Botanical Garden 72:852-863. [ Links ]
DEVORE, JL. 2003. Probabilidad y estadística para ingeniería y ciencias. 5ta. edición. International Thomson Editores. [ Links ]
GIBSON, DJ & JA NEWMAN. 2001. Biological Flora of the British Isles: F. arundinacea Schreber (F. elatior subsp. arundinacea (Schreber) Hackel). J. Ecol., 89:304-324. [ Links ]
KENKEL, NC. 1988. Pattern of self-thinning in jack pine: testing the random mortality hypothesis. Ecology, 69:1017-1024. [ Links ]
HARPER, JL. 1977. Population biology of plants. Academic Press, London. [ Links ]
INFOSTAT 2002. Infostat version 1.1. Grupo InfoStat, SCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
MCNAUGHTON, SJ. 1979. Grassland-herbivore dynamics. Pp. 46-81 in: ARE Sinclair & M Norton-Griffiths (eds.). Serengeti: Dynamics of an Ecosystem. University of Chicago Press. [ Links ]
MEHRHOFF, LA & R TURKINGTON. 1990. Microevolution and site-specific outcomes of competition among pasture plants. J. Ecol., 78: 745-756. [ Links ]
PIELOU, EC. 1969. The measurement of aggregation. Pp. 90-98 in: EC Pielou (ed.). An Introduction to Mathematical Ecology. Wiley Intersciences, New York. [ Links ]
SILVERTOWN, JW & J LOVETT-DOUST. 1993. Introduction to plant population biology. Blackwell Scientific Publications, Oxford. [ Links ]
TILMAN, D. 1982. Resource competition and community structure. Princeton University Press, Princeton. [ Links ]
WHITE, J. 1985. The thinning rule and its application to mixtures of plant populations. Pp 291-309 in: J White (ed.). Studies on Plant Population Demography. Acdemic Press, Londres. [ Links ]
YODA, K; T KIRA; H OGAWA & K HOZUMI. 1963. Intraspecific competition among higher plants. XI. Self-thinning in overcrowded pure stands under cultivated and natural conditions. Journal of Osaka City University Institute of Plitechnics 14:107-129. [ Links ]

