










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.17 n.2 Córdoba jul./dic. 2007
TRABAJOS ORIGINALES
Abundancia de Mus musculus en granjas avícolas: efectos locales vs. efectos espaciales
Vanina León 1, *, Juan S Guidobono1 & María Busch1, 2
1. Depto. de Ecología, Genética y Evolución, Fac. de Cs. Exactas y Naturales, Univ. de Buenos Aires, Argentina.
2. Consejo Nacional de Investigaciones Científicas y Tecnológicas.
* Departamento de Ecología, Genética y Evolución, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Cuidad Universitaria, Pabellón II, 4º piso. (C1428EHA) Buenos Aires. Email: vleon@ege.fcen.uba.ar
Recibido: 31 de enero de 2006;
Fin de arbitraje: 11 de julio de 2006;
Revisión recibida: 5 de octubre de 2006;
Aceptado: 25 de abril de 2007
Este trabajo fue arbitrado durante el proceso editorial de Marcelo Cassini.
RESUMEN. El objetivo de este trabajo fue evaluar el efecto de las variables internas y externas sobre la abundancia de Mus musculus en granjas avícolas. Consideramos como variables "internas" a aquellas características asociadas al estado de mantenimiento de la granja y a su infraestructura, y como "externas" o "espaciales" a aquellas que dependieron de la ubicación de la granja respecto a otras granjas y, por lo tanto, relacionadas a la probabilidad de invasión por roedores. El trabajo se desarrolló en diciembre 2004, enero, septiembre, octubre y diciembre de 2005 en granjas avícolas del Partido de Exaltación de la Cruz (Provincia de Buenos Aires, Argentina). El "estado de la granja" mostró una relación negativa con la abundancia de M. musculus, mientras que las otras variables fueron no significativas. No se observó concordancia entre el agrupamiento de granjas teniendo en cuenta distancias geográficas y esfuerzo de captura, ni autocorrelación espacial de la abundancia. La abundancia de M. musculus en las granjas avícolas estudiadas dependería de los efectos internos y no de los externos, por lo que el mantenimiento de las poblaciones dependería de una dinámica "in situ" y no del ingreso de individuos desde sitios vecinos, aunque también podría haber transporte pasivo por el hombre.
Palabras clave: Plaga; Control; Colonización; Movimientos; Autocorrelación espacial.
ABSTRACT. Mus musculus abundance in poultry farms: local vs spatial effects: The aim of this paper was to assess the effects of the internal and spatial (external) variables on the abundance of Mus musculus in poultry farms. The internal variables were the maintenance conditions of the farms and the number of sheds, while the external variables were related to the spatial location of each farm with respect to other farms and thus, to the probability of invasion from surrounding areas. This work was conducted on December 2004, January, September, October and December 2005 at poultry farms located at Exaltación de la Cruz, Buenos Aires Province, Argentina. If the abundance is mainly determined by the re-colonization through the displacement of the individuals, we expected the abundance of M. musculus to show an inverted relationship with the distance between farms; the abundance of M. musculus to show a positive relationship with the number of neighboring farms; as well as a correlation among the differences in abundance and the geographical distances between farms. We also expected the abundance of M. musculus to show a negative relationship with the farm maintenance condition, and a positive relationship with the number of sheds present in the farm. The only variable related to the variation in the abundance of M. musculus among farms was the maintenance condition, suggesting that the dynamics of this species in each farm is independent of the abundance in other farms, and does not depend on movements from their neighborhoods. The persistence of the population could be due to its recovery starting from the individuals who survived to the treatment that is permanently done in the farms, favored perhaps by a dense-dependent response of the reproduction or to the passive re-colonization through men.
Keywords: Pest; Control; Colonization; Movements; Spatial autocorrelation].
INTRODUCCIÓN
En agroecosistemas de la Pampa Central las comunidades de roedores incluyen cinco especies de sigmodontinos: Akodon azarae, Calomys laucha, C. musculinus, Oligoryzomys flavescens y Oxymycterus rufus, un caviomorfo, Cavia aperea, y los murinos Mus musculus y Rattus sp (Mills et al. 1991; Busch & Kravetz 1992; Bilenca et al. 1995). Calomys sp. es numéricamente dominante en las áreas de cultivo, mientras que A. azarae, O. flavescens y C. aperea son más abundantes en las zonas menos perturbadas (Mills et al. 1991; Busch & Kravetz 1992; Bilenca et al. 1995) M. musculus y Rattus sp, están concentrados en el área domiciliar (Kravetz et al 1987; Miño et al. 2001).
Mus musculus es una especie de roedor de origen asiático de distribución cosmopolita y que ocupa una gran variedad de hábitats. Aunque se la considera principalmente comensal, y es en los ambientes urbanos donde alcanza sus máximas densidades, también se la puede encontrar en hábitats silvestres, y en muchos países es una plaga de la agricultura (Ylönen et al. 2002; Jacob et al. 2003). Sin embargo, en ambientes naturales no es un buen competidor frente a otras especies de pequeños roedores, especialmente en condiciones de alta densidad (Crespo 1966; Ryan et al. 1993; Busch et al. 2005).
Estudios realizados por Valenzuela (2001) mostraron que M. musculus muestra un rango muy estrecho de movimientos y su área de acción varía entre 10 y 100 m2 (Chambers et al. 2000). La mayoría de sus poblaciones esta dividida en pequeños demes (Anderson 1970; Berry 1986), por lo que en áreas rurales existiría como una serie de poblaciones relativamente aisladas (Ryan et al. 1993) con poco intercambio de individuos incluso entre poblaciones situadas en distintos galpones de una misma granja (Selander 1970). Sin embargo, en condiciones de alta densidad los individuos subordinados se verían forzados a emigrar (Petras 1967; Anderson 1970; Macintosh 1981) y por lo tanto contribuirían a conectar las distintas poblaciones.
En los agroecosistemas de la pampa central, a diferencia de los agroecosistemas australianos donde alcanza niveles de plaga, la presencia de M. musculus es ocasional en los campos de cultivo y sus bordes (Busch et al. 2005). Esta diferencia se ha atribuido a la presencia en la pampa de varias especies de roedores nativos, especialmente de Akodon azarae, la especie dominante en los límites de las granjas (Miño 2003) y en los bordes de los campos de cultivo y que sería competitivamente superior a M. musculus (Busch et al. 2005). En establecimientos de cría de aves se observa una distribución diferencial de los roedores nativos y comensales: M. musculus se encuentra casi exclusivamente en los galpones de cría, mientras que A. azarae es la especie dominante en los perímetros de la granja, y Calomys sp se encuentra principalmente en los galpones aunque es menos abundante que M. musculus (Miño et al. 2001). Esta última especie puede alcanzar altas abundancias en las granjas avícolas probablemente debido a que la alta disponibilidad de alimento y las condiciones de temperatura y humedad benignas le permiten reproducirse durante todo el año, y debido a la menor abundancia de especies competidoras, ya que A. azarae no se encuentra en los galpones de aves (Miño 2003). Sin embargo, hay una fuerte mortalidad debido a la aplicación de medidas de control, especialmente rodenticidas anticoagulantes (Gómez Villafañe et al 2001; Gómez Villafañe 2003).
En la mayor parte de las granjas, el control más efectivo se realiza en las casas de manera que M. musculus se encuentra principalmente en los galpones donde hay alimento permanente y poco movimiento humano. El mantenimiento de la granja y los galpones, así como el grado de aplicación de medidas de control, son muy variables entre granjas. Mientras que en algunas se mantiene permanentemente la cobertura vegetal baja, tanto entre galpones como en los perímetros, no se acumula materiales como basura, leña, o maquinaria en desuso en el predio, y se mantienen los galpones en buenas condiciones (en cuanto al estado del cielorraso, entramado de alambre, paredes y aberturas), en otras hay un gran desarrollo de malezas y acumulación de basuras (Gómez Villafañe et al. 2001; Gómez Villafañe 2003). Gómez Villafañe et al. (2001) observaron una asociación positiva entre el grado de enmalezamiento de los perímetros de las granjas y el grado de infestación por roedores (sin discriminar especies), sugiriendo que la probabilidad de recolonización favorecía la infestación. También describieron una asociación negativa entre infestación y grado de mantenimiento de los galpones (galpones en peor estado tenían más roedores). Sin embargo, trabajos posteriores mostraron que, a diferencia de las especies silvestres, M. musculus no mostraba variaciones de abundancia según el grado de enmalezamiento del perímetro (Miñarro 2002), ni por la presencia de galpones cercanos a la periferia (Miño 2003).
Relevamientos efectuados en granjas avícolas del Partido de Exaltación de la Cruz (Provincia de Buenos Aires, Argentina) mostraron que, pese a la aplicación de medidas de control, M. musculus está presente prácticamente en el 100% de las granjas. La persistencia de sus poblaciones puede deberse a su recuperación a partir de los individuos que sobreviven al tratamiento (respuesta local), o al hecho de que los controles son asincrónicos entre granjas lo que podría generar una dinámica de metapoblaciones en la que la permanencia de las poblaciones locales depende de la recolonización desde áreas vecinas. Tampoco se descarta el movimiento pasivo de individuos transportados por el hombre con el alimento o los pollos, como proponen Ryan et al. (1993) para M. musculus en granjas de Irlanda.
El rango de movimientos de M. musculus es limitado y poco abundante en los alrededores de las granjas, por lo que los individuos que colonicen deberían atravesar las distancias que separan las granjas que se encuentran ubicadas relativamente aisladas unas de otras dentro de una matriz de campos de cultivo. Por otro lado, la presencia de especies de roedores silvestres en los alrededores también dificultaría su traslado de una granja a otra. En este sentido, este trabajo pretende comprender los mecanismos que hacen que las poblaciones de M. musculus persistan en estos ambientes con el fin de contribuir al diseño de medidas de control más eficientes, invirtiendo más esfuerzo en aquéllas que resulten en un mayor impacto sobre la abundancia a corto y largo plazo. Específicamente nos proponemos establecer la relación entre las características propias de las granjas (variables internas – efectos locales) y las características de las granjas en relación a su ubicación respecto a otras granjas (variables externas – efectos espaciales) y la abundancia de M. musculus. Debido a la gran mortalidad causada por las medidas de control implementadas, la abundancia de roedores en las granjas dependería de la recolonización, por lo tanto esperamos que: (1) la abundancia de M. musculus muestre una relación inversa con la distancia entre granjas; (2) la abundancia de M. musculus muestre una relación positiva con el número de granjas vecinas; (3) haya una correlación entre las diferencias en abundancia y las distancias geográficas entre granjas. Por otro lado, esperamos también que la abundancia de M. musculus muestre: (4) una relación negativa con el grado de mantenimiento de la granja; y (5) una relación positiva con el número de galpones presentes en la granja.
MÉTODOS
El presente estudio se llevó a cabo en diciembre de 2004, enero, septiembre, octubre y diciembre de 2005 en granjas avícolas del Partido de Exaltación de la Cruz (34ºS; 59ºE), Provincia de Buenos Aires, Argentina (Fig. 1a) distribuidas en un área de 190 km2. Como se observa en la figura 1b, las granjas no están distribuidas uniformemente en el espacio sino que se encuentran formando grupos. Las granjas González, Grevé, Niglia, Peter, Balduzzi y Noriega se encuentran dentro de una zona relativamente urbanizada correspondiente a Diego Gaynor (Fig. 2b). Las granjas Stanicia, Abuelo y Dogo son las más cercanas al casco urbano de Capilla del Señor (Fig. 2b), la cual es una zona mucho más grande y urbanizada (5725 habitantes) que la mencionada anteriormente.


Figura 1. a) Localización del Partido de Exaltación de la Cruz, Provincia de Buenos Aires, Argentina; b) Localización geográfica de las granjas muestreadas (subrayadas) y no muestreadas en el área de estudio.
Figure 1. a) Location of Exaltación de la Cruz, Buenos Aires Province, Argentina; b) Location of the farms sampled (underlined) and not sampled in the study area.
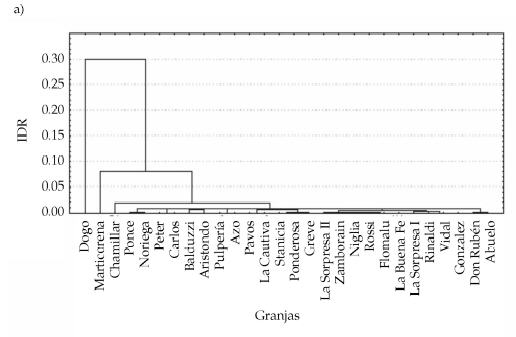

Figura 2. a) Agrupamientos obtenidos del análisis de cluster para las diferencias de IDR entre granjas. IDR = número de individuos capturados/número trampas x noche; b) Agrupamientos obtenidos del análisis de cluster para las distancias geográficas entre granjas muestreadas.
Figure 2. a) Groups obtained from the cluster analysis for the differences in IDR among farms. IDR = number of captured individuals /number of tramps per night; b) Groups obtained from the cluster analysis for the geographic distances between the sampled farms.
Área de estudio
El área está situada en la subregión Pampa Ondulada caracterizada por el clima templado, con temperatura y precipitación media anual de 16ºC y 1000 mm respectivamente. Los pastizales originales se vieron modificados por la introducción de árboles y el desempeño de actividades agrícola-ganaderas, quedando relictos de la flora original restringidos a terraplenes y bordes de caminos y cultivos (Mills et al. 1991; Busch & Kravetz 1992).
Actualmente se cultiva soja, maíz, trigo y girasol. Se cría ganado vacuno, equino, caprino, porcino y aves en forma intensiva. Esta última actividad comenzó aproximadamente en 1980 y mostró un rápido incremento, alcanzando una cantidad mayor a 130 granjas avícolas en la zona de estudio y sus alrededores (Miño 2003).
Las granjas estudiadas se dedican a la cría de pollos que son provistos por empresas que se encargan del comercio mayorista de este producto. Los pollos de aproximadamente 5 días de edad son ingresados a las granjas y ubicados en galpones. Los galpones son rectangulares, de perímetro y superficie variables. El piso es de tierra y está cubierto por una cáscara de girasol o arroz ("cama") que aísla a los pollos de la humedad y del producto de sus excretas. Las paredes de ladrillos o chapa son de baja altura (50 cm aproximadamente), y se continúan con un entramado de alambre que llega hasta el techo. Este último es de chapa galvanizada a dos aguas y puede constar o no de un cielorraso de telgopor o lona como aislante de la temperatura. En el interior de los galpones se encuentran las tolvas con alimento balanceado, los bebederos, y un sistema de riego que permite mojar a las aves y mantenerlas frescas cuando la temperatura ambiente es elevada (Gómez Villafañe 2003). Durante el período de crianza, a las aves se les proporciona agua, alimento y temperatura adecuada. Los pollos son retirados a los 45-50 días, cuando alcanzan un tamaño adecuado para su consumo, en camiones de la misma empresa que los proveyó, se deja de brindar alimento y agua, y el piso o "cama" es retirado. En este período de descanso, que dura entre 15 y 20 días, se preparan los galpones para un nuevo ingreso de pollos. Las granjas son visitadas semanalmente por un supervisor, perteneciente a la empresa proveedora para controlar el estado de los pollos (Gómez Villafañe 2003). En general, los propietarios de las granjas son quienes se encargan de la aplicación de rodenticidas, tanto en el período de cría de las aves como en el de descanso, por lo que se observa una gran variabilidad en los momentos y en la duración de aplicación de este control, como así también en las marcas y por lo tanto en las drogas utilizadas para controlar a los roedores.
Muestreo de roedores
Se determinó la abundancia de roedores en 28 granjas avícolas (Fig. 1b), en primavera (12 granjas) y verano (16 granjas), mediante la instalación de 1082 trampas de captura viva tipo Sherman. En cada granja se muestreó una porción de su perímetro (100 m) y un máximo de tres galpones seleccionados al azar. Las trampas de captura viva fueron ubicadas en los laterales de los galpones y espaciadas cada 20 metros, mientras que en los perímetros estuvieron espaciadas cada 10 metros. Cada granja se muestreó una sola vez y en cada muestreo las trampas fueron revisadas todas las mañanas durante 3 días consecutivos. Para cada individuo capturado se determinó la especie, longitud, peso, sexo, estado reproductivo, fecha y lugar de la captura. La abundancia de roedores en cada granja se estimó utilizando un índice de densidad relativo al esfuerzo de captura, IDR = número de individuos capturados/número trampas x noche.
Variables estudiadas
Las características internas de las granjas consideradas en el análisis y seleccionadas a partir de resultados previos obtenidos por Gómez Villafañe et al. (2001), fueron el número de galpones (muestreados y no muestreados) y su estado de mantenimiento, medido con una escala cualitativa (0 = malo, 1 = regular, 2 = bueno y 3 = muy bueno) basada en la presencia o no de acumulaciones de basura, leña, maquinaria en desuso y el mantenimiento del césped, tanto alrededor de los galpones como en el predio en general. Es importante aclarar que no se incorporaron en este modelo los métodos de control químico de roedores ya que Gómez Villafañe et al (2001) no encontraron asociación entre la aplicación de rodenticidas anticoagulantes y el grado de infestación por roedores, en granjas avícolas localizadas en la misma zona de estudio donde se desarrolló el presente trabajo.
Se ubicaron geográficamente las granjas de la zona de estudio mediante un GPS y el programa GPS Utility versión 4.20.4 (Murphy 2005) (Fig. 1b), y se calculó la distancia entre ellas utilizando el programa OziExplorer versión 3.85.6.2a (Newman & Newman 1999). Las características externas de las granjas consideradas fueron: cantidad de granjas presentes en un radio de 500 m, distancia a la granja más cercana, y distancia promedio de las granjas situadas a una distancia menor de 10 km en cada cuadrante formado por dos rectas perpendiculares que se corten en la granja focal. Para estas tres últimas variables, se tuvo en cuenta la presencia de todas las granjas en el área de estudio, hayan sido o no muestreadas.
Análisis de los datos
Para detectar cual o cuáles de las variables internas y externas se relacionaban con la abundancia se realizaron regresiones con aleatorización mediante el programa RT version 2.1 (Manly 1997). Se realizó una regresión múltiple incluyendo todas las variables a la vez. Para evaluar la existencia de autocorrelación espacial de la abundancia de roedores se realizaron análisis de agrupamiento de Cluster, en base a las diferencias de IDR y en base a las distancias geográficas mediante el programa Statistica versión 5.5 y se realizó un test de Mantel mediante el programa Pop tools version 2.6.6 (Hood 2005). Las matrices utilizadas fueron la de diferencias de IDR, y la de inversas de las distancias geográficas entre granjas, en este análisis se analizaron por separado los datos de primavera y verano.
RESULTADOS
Se capturó un total de 148 individuos pertenecientes a la especie M. musculus (141 individuos se capturaron en los galpones de cría, y sólo siete en los perímetros) y 30 individuos de A. azarae, de los cuáles sólo nueve fueron capturados en los galpones. El IDR de M. musculus de las granjas varió entre 0 y 0.49. El número máximo de individuos de M. musculus por granja fue de 45 (equivalente a 500 individuos por hectárea), mientras que la densidad promedio fue de 17.3 individuos/ha, considerando exclusivamente el área de los galpones (nº individuos capturados / área promedio galpón x nº galpones).
El número de galpones por granja varió entre uno y 10, siendo tres la cantidad más frecuente. De las 28 granjas muestreadas, a seis se les asignó el estado malo, a seis regular, a 10 estado bueno y a seis estado muy bueno.
La mayor distancia entre granjas fue de 17.7 km, y la menor de 0.1 km, el número de granjas en un radio menor a 500 m varió entre cero y dos, y la distancia promedio de las granjas situadas a una distancia menor de 10 km varió entre 0.36 y 9 km.
Se encontró una relación inversa entre el "estado de la granja" y el IDR de M. musculus; las otras variables consideradas no fueron significativas (Tabla 1). Los grupos de granjas similares de acuerdo al IDR no se corresponden con los grupos observados para distancia geográfica (Figura 2). Coincidentemente, de acuerdo al test de Mantel, no hay una relación significativa entre las diferencias de abundancia de M. musculus entre granjas y su separación espacial para primavera y verano (R = -0.13 y P = 0.4, R = -0,003 y P = 0.168, respectivamente).
Tabla 1. Resultado de la regresión múltiple entre el IDR de M. musculus y las variables internas y externas. IDR = número de individuos capturados/número trampas x noche.
Table 1. Results of the multiple regression between the IDR of M. musculus and internal and external variables. IDR= captured individuals per trap and per night.

DISCUSIÓN
La ausencia de una autocorrelación espacial de la abundancia y la ausencia de efectos significativos de las variables relacionadas con la probabilidad de recolonización, sugerirían que para el mantenimiento de las poblaciones de M. musculus no sería determinante la recolonización a través de desplazamientos de los individuos entre las granjas estudiadas. Las granjas de la zona presentarían poca migración debido a la baja densidad y, por lo tanto, no estarían formando parte de una única metapoblación. Este resultado contrasta con lo encontrado por Gómez Villafañe (2003), quién sí encontró una autocorrelación espacial de la abundancia para R. norvegicus, lo que estaría relacionado al hecho de que R. norvergicus tiene un área de acción mucho mayor (entre 24 y 525 m2) que la correspondiente a M. musculus. Estudios realizados por Valenzuela (2001) mostraron que M. musculus muestra un rango muy estrecho de movimientos, y principalmente entre los galpones de cría, coincidiendo con la pequeña área de acción (10 m2), citada por Chambers et al (2000). A su vez, el movimiento desde o hacia el perímetro de la granja se vería limitado por la interferencia con A. azarae, especie dominante en los límites de las granjas (Miño et al. 2001). Nuestros resultados concuerdan con los obtenidos por Miñarro (2002), quién observó que controlando la cobertura vegetal baja en los límites de la granja se observa una disminución de roedores, debida principalmente al efecto sobre A. azarae y no a M. musculus, mientras que esta última sí fue afectada por la aplicación de rodenticida en los galpones de cría. Por otro lado, Miño (2003) encontró que granjas con interfase (granjas que presentan galpones en contacto con bordes de campos de cultivo) presentaron mayor proporción de especies silvestres que granjas sin interfase, pero no hubo diferencias entre estos grupos de granjas en cuanto a M. musculus.
Mus musculus es una especie territorial, y en condiciones de baja densidad los individuos tienden a permanecer en sus territorios, pero cuando la densidad aumenta, los individuos subordinados son forzados a dispersarse para poder acceder a su propio territorio. La densidad promedio en las granjas estudiadas fue baja, y probablemente no llegó a esos niveles en los cuales los individuos subordinados se ven obligados a emigrar (Petras 1967; Anderson 1970; Macintosh 1981), y por lo tanto las poblaciones de las distintas granjas tendrían una dinámica independiente, determinada por sus características internas. A su vez, la competencia interespecífica en los alrededores de las granjas tendría mayor efecto que la intraespecífica dentro de estas, debido a los controles efectuados permanentemente que mantienen la abundancia de roedores baja. La permanente remoción de individuos daría oportunidad a los individuos más jóvenes o subordinados de ocupar territorios vacantes, sin necesidad de dispersarse.
El efecto del estado de la granja (que está relacionada con el estado de los galpones y el mantenimiento de la cobertura vegetal baja), concuerda con la relación encontrada por Gómez Villafañe et al (2001) entre el grado de infestación por roedores, el estado de los galpones y una cobertura vegetal alta.
En resumen, la abundancia de M. musculus presenta una relación inversa con las características internas de las granjas. Este hecho estaría relacionado a que granjas en peor estado tienen mayor cantidad de refugios (alta cobertura de césped) y más fuentes de alimento (por ejemplo, acumulaciones de residuos orgánicos) que granjas en buen estado. A su vez, la posibilidad de reinvasión desde granjas vecinas, al menos con las densidades encontradas en las granjas estudiadas, sería relativamente baja. La persistencia de las poblaciones podría deberse a su recuperación a partir de los individuos que sobreviven al tratamiento que se realiza permanentemente en las granjas, favorecida tal vez por una respuesta densodependiente de la reproducción (Pelz 2003) o a la recolonización pasiva a través del hombre. Esto reafirma lo propuesto por Miño (2003), que las medidas de control efectivas para roedores silvestres y para M. musculus son diferentes. Sin embargo, para evaluar el papel de la recolonización en el mantenimiento de las poblaciones de M. musculus, es necesario realizar estudios de movimientos.
AGRADECIMIENTOS
A los pobladores de Diego Gaynor, Capilla del Señor, La Lata y Puerto Chenaut por su permanente colaboración. A C. González Fischer, C. Guerra Navarro, M. Escolar y M. Totino por su colaboración en los muestreos. A R. Cavia por su ayuda con los análisis estadísticos.
Estos estudios han sido financiados por la Agencia Nacional de Promoción Científica y Tecnológica (PICT 12303) y la Universidad de Buenos Aires (UBACyT X098). El trabajo se realizó en el marco de una beca doctoral otorgada a Léon Vanina por la Agencia Nacional de Promoción Científica y Tecnológica.
BIBLIOGRAFÍA
ANDERSON, PK 1970. Ecological structure and gene flow in small mammals. Symposia of the Zoological society of London, 26:299- 325. [ Links ]
BERRY, RJ. 1986. Genetical processes in wild mouse populations. Past myth and present knowledge. Curr. Top. Microbiol. Immunol., 127:86-94. [ Links ]
BILENCA, D; EA CITTADINO & FO KRAVETZ. 1995. Influencia de la actividad de Cavia aperea sobre la estructura del hábitat y la distribución de Akodon azarae y Oryzomys flavescens (Rodentia: Caviidae, Muridae) en bordes de cultivos de la región pampeana (Argentina). Iheringia, Série Zoología, 79:67-65. [ Links ]
BUSCH, M; D BILENCA; EA CITTADINO & G CUETO. 2005. Effect of removing a dominant competitor, Akodon azarae (Rodentia, Sigmodontinae) on community and population parameters of small rodent species in Central Argentina. Austral Ecology, 30:168-178. [ Links ]
BUSCH, M & FO KRAVETZ. 1992. Competitive interactions among rodents (Akodon azarae, Calomys laucha, Calomys musculinus y Oligoryzomys flavescens) in a two- habitats system. Spatial and numerical relationship. Mammalia, 56:45-46. [ Links ]
CRESPO, JA. 1966. Ecología de una comunidad de roedores silvestres en el Partido de Rojas, Provincia de Buenos Aires. Revista del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia", 1:79-134. [ Links ]
CHAMBERS, LK; G SINGLETON & JK CHARLES. 2000. Movements and social organization of wild house mice (Mus musculus) in the weatlands of northwestern Victoria, Australia. J. Mammal., 81(1):59-69. [ Links ]
GÓMEZ VILLAFAÑE, IE. 2003. Estudio de la Ecología y epidemiología de Rattus sp. en un ambiente urbano y granjas avícolas del Partido de Exaltación de la Cruz, Buenos Aires, Argentina. Tesis doctoral, Universidad de Buenos Aires, Argentina. [ Links ]
GÓMEZ VILLAFAÑE, IE; DN BILENCA; R CAVIA; MH MIÑO; EA CITTADINO & M BUSCH. 2001. Environmental factors associated with rodent infestations in Argentine poultry farms. Br. Poult. Sci., 42:300-307. [ Links ]
HOOD, GM. 2005. PopTools version 2.6.6. http://www.cse.csiro.au/poptools [ Links ]
JACOB, J; H YLÖNEN; MJ RUNCIE; DA JONES & GR SINGLETON. 2003. What affects bait uptake by house mice in Australian grain fields? J. Wildl. Manag., 67:341-351. [ Links ]
KRAVETZ, FO; MI BELLOCQ; M BUSCH; SM BONAVENTURA & A MONJEAU. 1987. Efecto de la aplicación de un anticoagulante sobre la comunidad de roedores de campos de cultivo. Anales del Museo de Historia Natural, Valparaíso, 18:153-156. [ Links ]
MACINTOSH, JH. 1981. Behaviour of the house mouse. Symposia of the Zoological Society of London, 47:337-365. [ Links ]
MANLY, BFJ. 1997. RT a program for randomization testing. Version 2.1. [ Links ]
MILLS, JN; B ELLIS; K MCKEE; J MAIZTEGUI & JE CHILDS. 1991. Habitat associations and relative densities of rodent population in cultivated areas of Central Argentina. J. Mammal., 72: 470-479. [ Links ]
MIÑARRO, FO. 2002. Evaluación experimental de métodos de ordenamiento del hábitat para el control de roedores en granjas avícolas de la Provincia de Buenos Aires. Tesis de Licenciatura, Universidad de Buenos Aires, Argentina. [ Links ]
MIÑO, MH. 2003. Caracterización de las comunidades de roedores en granjas avícolas del partido de Exaltación de la Cruz (Provincia de Buenos Aires). Tesis Doctoral, Universidad de Buenos Aires, Argentina. [ Links ]
MIÑO, MH; R CAVIA; IE GÓMEZ VILLAFAÑE; DN BILENCA; EA CITTADINO & M BUSCH. 2001. Estructura y diversidad de dos comunidades de pequeños roedores en agroecosistemas de la provincia de Buenos Aires, Argentina. Boletín de la Sociedad Biológica de Concepción, Chile, 72:67-75. [ Links ]
MURPHY, AS. 2005. GPS Utility versión 4.20.4. http://www.gpsu.co.uk [ Links ]
NEWMAN, D & L NEWMAN 1999. OziExplorer version 3.85.6.2a. [ Links ]
PELZ, HJ. 2003. Management – Field [4] Pp. 233-237 in: GR Singleton; LA Hinds; CH Krebs & D Spratt. Rats, mice and people: rodent biology and management. Australian Centre for International Agricultural Research. [ Links ]
PETRAS, ML. 1967. Studies of natural populations of Mus. Biochemical polymorphisms and their bearing on breeding structure. Evolution, 21:259-74. [ Links ]
RYAN, AW; EJ DUKE & JS FAIRLEY. 1993. Polymorphism, localization and geographical transfer of mithocondrial DNA in Mus musculus domesticus (Irish house mice). Heredity, 70:75-81. [ Links ]
SELANDER, R.K. 1970. Biochemical polymorphism in populations of the house mouse and old-field mouse. Symp. Zool. Soc. Lond., 26:73-91. [ Links ]
VALENZUELA, L. 2001. Variación estacional y desplazamientos de poblaciones de pequeños roedores en dos granjas avícolas. Tesis de Licenciatura, Universidad de Buenos Aires, Argentina. [ Links ]
YLÖNEN, H; J JACOB; MJ DAVIES & GR SINGLETON. 2002. Predation risk and habitat selection of Australian house mice Mus musculus during an incipient plague: Behaviour due to food depletion. Oikos, 99:284-289. [ Links ]

