










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.17 n.2 Córdoba jul./dic. 2007
TRABAJOS ORIGINALES
Composición, distribución y relaciones tróficas de la ictiofauna del río Negro, Patagonia Argentina
Pablo A Alvear1, *, Magalí Rechencq1, 2, Patricio J Macchi1, Marcelo F Alonso1, Gustavo E Lippolt1, María A Denegri1, 2, Guillermo Navone1, Eduardo E Zattara1, Martín I García Asorey2, 3 & Pablo H Vigliano1
1. Grupo de Evaluación y Manejo de Recursos Ícticos, Centro Regional Universitario Bariloche, Universidad Nacional del Comahue, Bariloche, Río Negro, Argentina.
2. CONICET, Centro Regional Universitario Bariloche, Univ. Nac. del Comahue, Bariloche, Río Negro, Argentina.
3. Centro Nacional Patagónico-CONICET, Puerto Madryn, Chubut, Argentina.
* Quintral 1250, 8400 San Carlos de Bariloche, Río Negro Argentina. Email: alvearpabloa@yahoo.com.ar
Recibido: 14 de septiembre de 2005;
Fin de arbitraje: 10 de marzo de 2006;
Revisión recibida: 2 de mayo de 2007;
Segunda revisión recibida: 31 de mayo de 2007:
Aceptado: 1 de junio de 2007.
Este trabajo fue arbitrado durante el proceso editorial de Marcelo Cassini.
RESUMEN. La provincia ictiogeográfica Patagónica se caracteriza por una llamativa pobreza de especies. El río Negro es el curso de agua más importante de esta región y a pesar de sus dimensiones e importancia socioeconómica su fauna íctica ha sido poco estudiada. Este trabajo describe algunos aspectos biogeográficos y características ecológicas de su ictiofauna. Se muestrearon estacionalmente cinco sitios a lo largo del río, por medio de redes de enmalle, pesca eléctrica, redes de arrastre costero y atarraya. Se analizó la composición y distribución de peces y se estudió la dieta para determinar los roles tróficos y los niveles de competencia y de piscivoría. Se capturaron ejemplares de 13 especies: siete de origen nativo y seis exóticas, de las cuales se destaca la novedosa presencia de la coridora (Corydoras paleatus) y la carpa común (Cyprinus carpio), ampliando su distribución austral. Las especies más importantes fueron las nativas pejerrey patagónico (Odontesthes hatcheri), perca bocona (Percychthis colhuapiensis), perca de boca chica (P. trucha) y madrecita (Jenynsia multidentata). Los índices ecológicos indican que el curso inferior es diferente al resto del río presentando una dominancia de pejerrey patagónico y madrecita. En este sector se observó una mayor importancia de especies exóticas. Se encontraron ocho grupos tróficos, existencia de solapamientos en las dietas entre componentes nativos y exóticos, y evidencia de piscivoría importante entre las especies. La presencia y distribución de los peces en el río Negro está fuertemente influenciada por fenómenos de antropocoria accidental o intencional. Es necesario desarrollar estudios que analicen el impacto generado por las introducciones y la evolución de las relaciones entre los componentes nativos y exóticos, teniendo en cuenta que la trucha arco iris y la carpa común han influido negativamente en otros lugares del mundo.
Palabras clave: Peces nativos; Peces exóticos; Índices ecológicos; Distribución de peces.
ABSTRACT. Composition, distribution and trophic relationships of the fish fauna of the Negro river, Patagonia Argentina: The Patagonic ictiogeographic region is characterized by a low species diversity. Despite that the Negro river is the most important water body in the region, both in terms of the size of its drainage basin and of its socioeconomic importance, its fish fauna has scarcely been studied. The present paper addresses biogeographic and ecological characteristics related to its fish. Gill net gangs, electro fishing and beach seine nets were used to sample seasonally five stations along the river. Fish fauna composition and distribution were analyzed throughout different ecological indexes. Diet composition was also analyzed to establish trophic roles, competition and piscivory. We caught seven native and six exotic species; two of them (Corydoras paleatus and Cyprinus carpio) represent a widening of their austral distribution. The natives Odontesthes hatcheri, Percychthis colhuapiensis, P. trucha and Jenynsia multidentata were more important in terms of numbers in catches. Ecological indexes show that the lower part of the Negro river differs from the rest of the river with higher numbers of O. hatchery and J. multidentata. Eight trophic groups were found as well as evidence of diet overlapping and considerable piscivory. Presence and distribution of fish in the Negro river is strongly influenced by introduction by men, either accidentally or intentionally. Impacts by introduced species need to be addressed, specially in terms of emerging processes between native and exotic fish, taking into special consideration the negative impacts that the rainbow trout and the carp have already had in different parts of the world.
Keywords: Native fish; Exotic fish; Ecological indexes; Fish distribution.
INTRODUCCIÓN
La provincia icitiogeográfica Patagónica, ubicada dentro de la subregión Austral, presenta la mayor pobreza específica de toda la región Neotrópica. Abarca la parte sur de la Argentina, desde el río Colorado hasta Tierra del Fuego, y el sur de Chile desde Puerto Mont hasta Fuegia (Ringuelet 1975). Se caracteriza por el bajo número de especies de peces presentes, que no supera las 29, entre nativas y exóticas (Baigún & Ferriz 2003), contrastando notablemente con las más de 350 especies presentes en la vecina provincia Parano-Platense (López et al. 2005).
En la provincia Patagónica se distinguen dos regiones ictiofaunísticas: 1) la región del Sur, o de Santa Cruz, que se extiende desde los 45º latitud sur hasta el estrecho de Magallanes (Arratia et al. 1983). Los cuerpos de agua de esta región, se caracterizan por la presencia de especies nativas de origen patagónico y salmónidos exóticos introducidos desde principios del siglo XIX; y 2) la región del Norte o Patagonia antigua que se extiende entre los ríos Colorado y Chubut, incluyendo aguas de las Provincias de Neuquén, Río Negro y Chubut. Además de las especies mencionadas para la Región del Sur, esta región muestra un enriquecimiento dado por la presencia de representantes de origen brasílico y exóticos no salmónidos. La zona comprendida entre los ríos Colorado y Negro constituyen el límite entre las provincias Patagónica y Parano-Platense, abarcando una extensa área de transición considerada un ecotono cuyo límite sur es el río Negro (Arratia et al. 1983; Almirón et al. 1997).
A pesar de su gran caudal, importancia e impacto socioeconómicos el río Negro, y particularmente su ictiofauna, han sido poco estudiados. Este curso de agua es el más importante de la región Patagónica. En su cuenca se desarrolla una importante actividad económica, principalmente la fruti horticultura y la cría de ganado bovino, con una alta densidad poblacional humana concentrada principalmente en centros urbanos. Además, sus aguas se utilizan para irrigación, turismo, recreación, industrias, abastecimiento doméstico y descarga de deshechos (Zattara, obs. pers.). Su ictiofauna incluye especies de valor deportivo, que motivan la existencia de numerosos clubes y asociaciones de pesca y una gran potencialidad de dicha actividad. Sin embargo, hay escasos trabajos específicos que incluyen estudios de carácter descriptivo (AIC - UNS 1995), taxonómico (López-Arbarello 2004) y de distribución (Liotta 2006). El presente trabajo señala algunos aspectos biogeográficos de la ictiofauna del río Negro documentando la composición, distribución, características ecológicas y relaciones tróficas de las especies de peces presentes en el río.
MATERIALES Y MÉTODOS
El río Negro nace de las confluencias de los ríos Neuquén y Limay y recorre 700 km en sentido Oeste – Este a través de la meseta Patagónica hasta su desembocadura en el océano Atlántico (Fig. 1). Es un río típicamente alóctono que recibe aportes de los hielos, nevadas y lluvias de la cordillera de los Andes (De Aparicio & Difrieri 1958). Su caudal está condicionado fuertemente por una cadena de represas hidroeléctricas ubicadas sobre los ríos Limay y Neuquén. El cauce es meandroso y su valle tiene gran cantidad de curvas y brazos secundarios que se activan estacionalmente. Atraviesa una región de gran pobreza hídrica, sin el aporte de afluentes a lo largo de toda su extensión (De Aparicio & Difrieri 1958). Sus riberas presentan mayormente cobertura de sauces (Salix fragilis L.) y vegetación típica de la provincia de monte. Existen asentamientos humanos ubicados a lo largo del río, con una mayor densidad poblacional en el Alto Valle (curso superior). Las actividades productivas varían a lo largo del río siendo la fruti-horticultura más importante en el curso superior y la ganadería extensiva en los cursos medio e inferior.
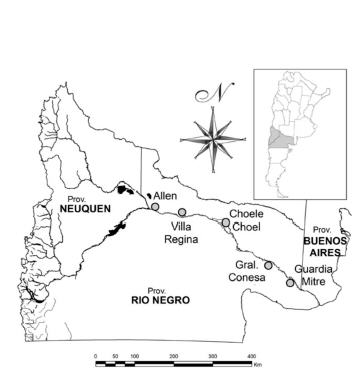
Figura 1. Curso del Río Negro: Ubicación de los sitios de muestreo.
Figure 1. Negro river course: Location of sampling sites.
Para la captura de peces se seleccionaron cinco sitios de muestreo entre la naciente y las cercanías de la desembocadura, en las localidades de Allen (Allen), Villa Regina (Regina), Choele Choel (Choele), Gral. Conesa (Conesa) y Guardia Mitre (GMitre) (Fig. 1). Cada sitio fue visitado en cuatro ocasiones con frecuencia estacional entre Septiembre 2003 y Agosto 2004. Se utilizaron cuatro trenes de redes de enmalle compuestos por paños de: 15, 20, 30, 40, 50, 60 y 70 mm de apertura de malla entre nudos contiguos. La superficie promedio de cada tren fue de 180 m2. Las redes se calaron al atardecer y fueron levantadas a primeras horas de la mañana siguiente (15 h de calado efectivo aproximadamente). Los hábitats muestreados incluyeron pozones, zonas de remanso, brazos secundarios de corriente lenta y sectores costeros del brazo principal. Para la fracción de peces pequeños y juveniles se utilizaron artes de pesca complementarias: una atarraya de 8 mm de luz de malla (esfuerzo estandarizado en diez lances por sitio, cubriendo aproximadamente 2.5 m2 por lance), una red de arrastre costero con copo de 10 mm de luz de malla (esfuerzo estandarizado en 300 m2 de superficie barrida) y un equipo de pesca eléctrica (300 m2 barridos por sitio). Estos artes se utilizaron en zonas de baja profundidad, brazos secundarios, zonas costeras del brazo principal o lagunas someras anegadas durante parte del año. De los ejemplares capturados se determinó la especie y su origen biogeográfico (Ringuelet & Aramburu 1961; Ringuelet 1975), conservando en formol los tractos digestivos para su posterior análisis en el laboratorio. Para la clasificación de las percas, cuya taxonomía se halla actualmente en revisión (Cussac et al. 1998, Ruzzante et al. 2003, López-Arbarello 2004), se siguió el criterio sugerido por López-Arbarello (2004) quien propone como especies válidas Percichthys trucha (perca de boca chica, incluyendo junto con P. trucha las anteriormente denominadas P. altispinnis y P. vinciguerrai) y P. colhuapiensis (perca bocona) con referencia a las especies encontradas en la cuenca del río Negro.
Se determinaron las abundancias relativas por especie, sitio y estación del año para cada arte de pesca calculando las capturas por unidad de esfuerzo en número (CPUEN). La unidad de esfuerzo para las redes de enmalle se fijó en 100 m2 de red / 15 horas de pesca y para las artes complementarias en 1 m2 barrido. Las características de la ictiofauna se estudiaron mediante los índices de dominancia de Simpson, diversidad de Shannon – Wiener y equitatividad de Pielou (1975). El grado de representación de las especies exóticas con respecto a las nativas se estudió por medio del coeficiente de integridad zoogeográfica (C.I.Z.) propuesto por Bianco (1990). En el grupo "exótico" se incluyeron salmónidos, ciprínidos y especies de origen brasílico. La expresión del índice es C.I.Z. = número de especies nativas / número total de especies. Los valores cercanos a cero indican un alto grado de importancia de elementos exóticos, en tanto que valores cercanos a uno indican mayor presencia de elementos nativos (Elvira 1995).
Para determinar las relaciones tróficas entre las especies se estudió la alimentación determinando la existencia de grupos con distintos roles tróficos, la competencia y los niveles de piscivoría. Los análisis se realizaron separando los peces en dos grupos, uno denominado "grandes peces", conformado por peces adultos de mayor porte, y el otro denominado "pequeños peces", formado por juveniles de perca de boca chica, bocona y Odontesthes hatcheri (pejerrey patagónico) junto con los adultos de las especies de pequeño tamaño. Los contenidos estomacales se clasificaron hasta el mínimo nivel taxonómico posible registrándose en cada caso el peso húmedo (precisión 0.001 g) de cada ítem de alimento (Wallace 1981). Las fracciones vegetales y el detrito se encontraron como masas discretas fácilmente separables y fueron pesadas.
Para el estudio del rol trófico de los peces "grandes" y "pequeños", se realizó un Análisis Discriminante (Norusis 1986) considerando a cada una de las especies como un grupo natural. Sus dietas fueron definidas sobre la base del porcentaje del peso total de cada categoría de alimento (Wallace 1981). Se estudió la competencia en la alimentación entre especies mediante el índice de solapamiento de Schöener (Schöener 1970), considerando superposición de dietas biológicamente significativas con valores de solapamiento iguales o mayores a 0.60 (Wallace 1981). Este análisis se realizó por especie, estación del año y sitio de muestreo. Para el estudio de la piscivoría se utilizaron los contenidos estomacales de los peces grandes, considerando el peso húmedo de cada especie de pez-presa por separado.
RESULTADOS
Se pescaron 1984 peces de trece especies, representando nueve órdenes y 11 familias (Tabla 1). De éstos, 1375 se capturaron con redes de enmalle y 609 con artes complementarios. Cabe destacar la aparición de dos nuevas especies no citadas anteriormente para este río: Corydoras paleatus (coridora) y Cyprinus carpio (carpa común).
Tabla 1. Especies de peces que componen la ictiofauna del río Negro. El asterisco indica las especies de origen brasilico.
Table 1. Fish species composition of the Negro River. The asterisk indicates a Brazilian origin.

En las capturas con redes de enmalle la abundancia de pejerrey patagónico fue importante a lo largo del río y principalmente durante el verano y la primavera, seguidas por las percas de boca chica y bocona (Fig. 2). La perca de boca chica mostró las mayores capturas en el verano, siendo más numerosa en el sector superior del río. Por el contrario la perca bocona fue más abundante en los sectores medio e inferior, con mayores capturas en verano y primavera. Oncorhynchus mykiss (trucha arco iris) se capturó principalmente en los sectores superior y medio, encontrándose un gradiente en la abundancia en este mismo sentido. La coridora apareció en casi todos los sitios de muestreo pero con capturas poco importantes. La carpa fue capturada en los cursos medio e inferior, siendo más abundante en este último sector donde apareció durante todo el año. Mugil lisa (lisa) fue capturada principalmente en el sector inferior del río en primavera y verano. El pejerrey bonaerense (O. bonariensis) fue capturado, en bajos números, en este mismo sector en otoño e invierno. Por último Diplomystes viedmensis (bagre aterciopelado) apareció en verano en la localidad de Allen también en números poco importantes. Estas dos especies no se incluyeron en los análisis debido a la baja captura.
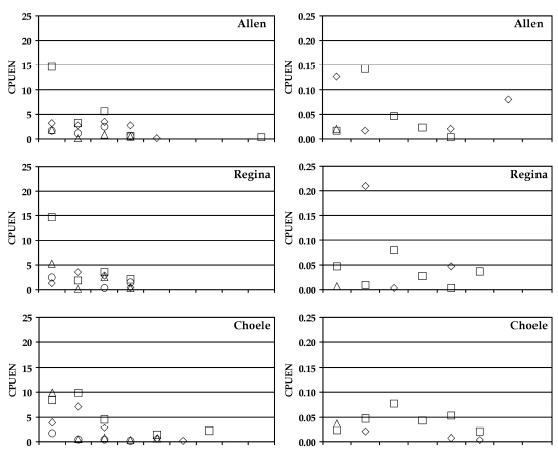
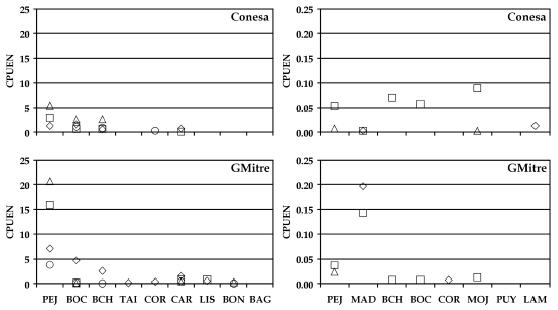
Figura 2. Abundancias relativas por sitios de muestreo, especie y estación del año. La columna de la izquierda indica capturas por unidad de esfuerzo en número (CPUEN) para capturas con redes de enmalle; la de la derecha para capturas con artes complementarias. Simbología: ![]() primavera;
primavera; ![]() verano;
verano; ![]() otoño;
otoño;![]() invierno. PEJ = pejerrey patagónico; BOC = perca bocona; BCH = perca de boca chica; TAI = trucha arco iris; COR = coridora; CAR = carpa común; LIS = lisa; BON = pejerrey bonaerense y BAG = bagre aterciopelado.
invierno. PEJ = pejerrey patagónico; BOC = perca bocona; BCH = perca de boca chica; TAI = trucha arco iris; COR = coridora; CAR = carpa común; LIS = lisa; BON = pejerrey bonaerense y BAG = bagre aterciopelado.
Figure 2. Relative abundances by sample site, species and season. The left column shows gill nets CPUEN values (captures by effort measured in numbers); the right column shows complementary methods CPUEN values. Simbology: ![]() spring;
spring; ![]() summer;
summer; ![]() autumn;
autumn; ![]() winter. PEJ = patagonian silverside; BOC = big mouth perch; BCH = small mouth perch; TAI = rainbow trout; COR = coridora; CAR = common carp; LIS = liza; BON = bonaerense silverside y BAG = belvet catfish.
winter. PEJ = patagonian silverside; BOC = big mouth perch; BCH = small mouth perch; TAI = rainbow trout; COR = coridora; CAR = common carp; LIS = liza; BON = bonaerense silverside y BAG = belvet catfish.
En los artes de pesca complementarios Jenynsia multidentata (madrecita) presentó valores importantes de CPUEN en Regina, Allen y Guardia Mitre. Para los otros sitios, la importancia mayoritaria en las capturas varió entre los juveniles de pejerrey patagónico y los de percas de boca chica y bocona (Fig. 2). Galaxias maculatus (puyen chico) y larvas ammocetes de Geotria australis (lamprea) fueron capturadas en bajo número sólo en Allen y Conesa respectivamente, durante la primavera. Cheirodon interruptus (mojarra) apareció principalmente en verano en casi todos los sitios de muestreo. La coridora fue capturada en bajo número en todos los sitios exceptuando Conesa, en donde la abundancia relativa fue importante.
Los índices utilizados para señalar características ecológicas se presentan en la Tabla 2. Guardia Mitre mostró los mayores valores de dominancia de Simpson, siendo el pejerrey patagónico y la madrecita las especies más importantes en las capturas de redes agalleras y de artes complementarios respectivamente. Los otros sitios de pesca no evidenciaron dominancia clara por parte de ninguna especie. Un resultado consistente se obtuvo con los índices de diversidad y equitatividad, los que mostraron menores valores en Guardia Mitre que para el resto de los sitios (Tabla 2).
Tabla 2. Índices de dominancia, equitatividad, diversidad, riqueza por origen biogeográfico y coeficiente de integridad zoogeográfico (C.I.Z.) por sitio de muestreo y arte de pesca. RE = redes de enmalle; AA = artes alternativos.
Table 2. Dominance, evenness, diversity, richness indexes by biogeographical origin and Zoogeographical Integrity Coefficient (C.I.Z.) for each sampling site and fish modality. RE = guillnets AA = alternative arts.
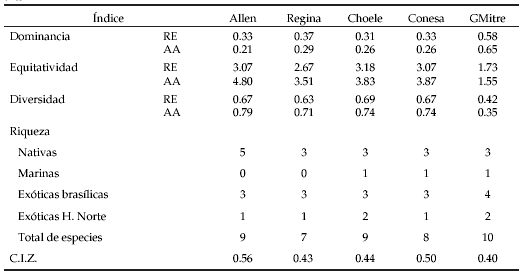
Considerando el número total de especies capturadas con todos los artes de pesca por sitio, la riqueza específica varió entre siete especies en Villa Regina y diez en Guardia Mitre (Tabla 2). De acuerdo con el origen biogeográfico, se obtuvo el siguiente agrupamiento: cinco nativas de origen patagónico, dos nativas marinas con ingresiones en el agua dulce y seis exóticas. De éstas, cuatro son de origen brasílico transplantadas desde otras cuencas argentinas y dos son originarias del Hemisferio Norte (Tabla 1). En el curso superior se observó una mayor riqueza de especies nativas que decrece levemente hacia la desembocadura del río, resultando la tendencia inversa a la que se observa en los grupos de especies exóticas.
Para calcular el coeficiente de integridad zoogeográfico se conformaron los siguientes grupos: a) especies nativas: pejerrey patagónico, percas de boca chica y bocona, puyen chico, bagre aterciopelado, lisa y lamprea, y b) especies exóticas: madrecita, mojarra, coridora y pejerrey bonaerense (brasílicas), carpa común y trucha arco iris (del Hemisferio Norte). Los valores hallados del C.I.Z. se presentan en la Tabla 2. Se observa una tendencia decreciente de los valores en el sentido naciente - desembocadura, con una menor importancia de especies exóticas en Allen. Los aportes de componentes nativos y exóticos variaron a lo largo del río, con mayor representatividad de especies brasílicas hacia los cursos medio e inferior.
Para los "peces grandes", considerando todos los sitios de muestreo y estaciones del año agrupados, el análisis discriminante formó cuatro grupos (DISCR, P < 0,0001, n = 520). Las carpas fueron discriminadas por su alto consumo de macrófitas, las lisas por una importante ingesta de algas filamentosas y detritos, y la perca bocona por un importante consumo de peces. La trucha arco iris, la perca de boca chica y el pejerrey patagónico formaron un grupo caracterizado por un amplio espectro trófico, aunque con diferencias en sus dietas. Las variaciones estacionales de los tipos de alimento consumidos por sitio de muestreo fueron importantes para la mayoría de las especies (Fig. 3), aunque no existe un patrón claro en el consumo de presas. Para los "peces pequeños" el análisis discriminante formó cuatro grupos (DISCR, P < 0,0001, n = 336). El puyen chico fue discriminado por el consumo de plancton, la mojarra por una importante ingestión de detritos, la madrecita por la incorporación de insectos adultos y algas. La coridora y los juveniles de ambas percas y pejerrey patagónico formaron un solo grupo altamente homogéneo en cuanto al consumo de larvas de insectos. Las variaciones estacionales de los tipos de alimento consumidos por sitio de muestreo fueron elevadas para la mayoría de las especies (Fig. 4). En general no se encontraron patrones de alimentación dependientes de la estación del año ni del sitio de muestreo, aunque los detritos y material vegetal cobraron mayor importancia hacia la desembocadura del río, particularmente en Guardia Mitre.

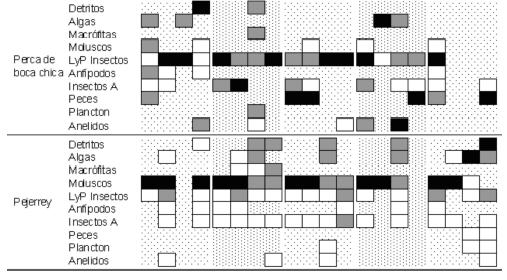
Figura 3. Porcentaje del consumo de ítems de alimento por grandes peces por sitio de muestreo, estación del año y especie. ![]() entre 0 y 10%;
entre 0 y 10%; ![]() entre 11 y 50%;
entre 11 y 50%;![]() > 51%.
> 51%.
Figure 3. Food consumption percentage by big fishes by species, sample site and season. ![]() between 0 and 10%;
between 0 and 10%;![]() between 11 and 50%;
between 11 and 50%; ![]() > 51%.
> 51%.

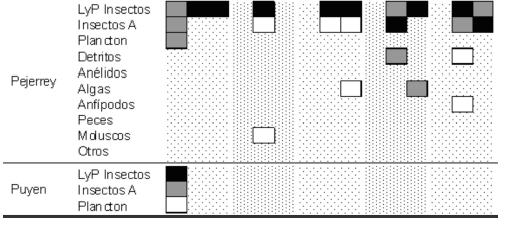
Figura 4. Porcentaje del consumo de ítems de alimento por pequeños peces por sitio de muestreo, estación del año y especie. ![]() entre 0 y 10%;
entre 0 y 10%; ![]() entre 11 y 50%;
entre 11 y 50%; ![]() > 51%.
> 51%.
Figure 4. Food consumption percentage by small fishes by species, sample site and season. ![]() between 0 and 10%;
between 0 and 10%; ![]() between 11 and 50%;
between 11 and 50%; ![]() > 51%.
> 51%.
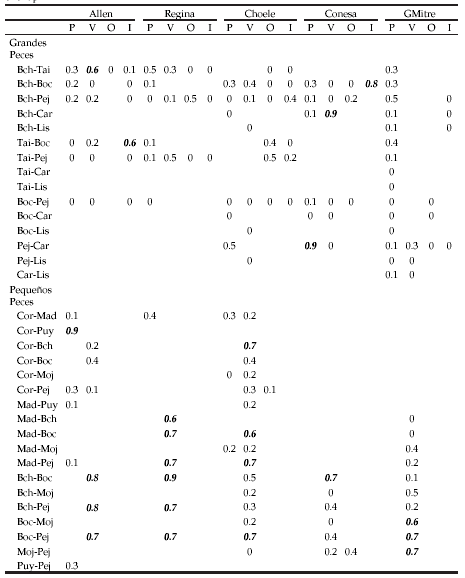
Considerando los grupos tróficos y la amplia diversidad de las dietas de varias de las especies, se encontraron pocos solapamientos significativos en las distintas épocas del año entre los peces grandes (Tabla 3). Las principales similitudes se observaron entre las dietas de ambas percas y con la trucha arco iris y entre las de la perca de boca chica, pejerrey patagónico y la carpa. Dada las bajas capturas de esta última especie, este resultado solo debe considerarse como indicativo. Para los peces pequeños se encontró mayor similitud en la alimentación, dando como resultado una mayor cantidad de solapamientos significativos (Tabla 3). Se encontraron similitudes entre las dietas de los juveniles de pejerrey, de ambas especies de perca y la madrecita; entre la mojarra y la perca bocona, y por último entre la coridora con el puyen y la perca de boca chica. El ictiófago más importante fue la perca bocona, que consumió peces de pequeño porte, incluyendo especies exóticas y juveniles de su misma especie. Las truchas arco iris, percas de boca chica y pejerreyes patagónicos también consumieron peces, aunque no fue un item de aparición continua en el tiempo ni a lo largo del río (Fig. 3). La trucha arco iris incorporó en su dieta juveniles de percas, pejerreyes y coridoras. La perca de boca chica consumió estas mismas especies y larvas de lamprea. El pejerrey patagónico consumió madrecitas, percas y pejerreyes.
Tabla 3. Solapamiento de dietas de grandes y pequeños peces por sitio de muestreo y estación del año. Los valores en negrita y cursiva indican solapamientos significativos.
Table 3. Big and small fish diet overlap by sample site and season. Values in bold italic indicate a significant overlap.
DISCUSIÓN
La captura estuvo compuesta por 13 especies: cinco nativas de origen patagónico, dos nativas de ingresión marina y seis exóticas, de las cuales cuatro son de origen brasílico y dos del hemisferio Norte. Las especies capturadas son de aparición común en los cuerpos de agua lóticos y lénticos de la región patagónica (Bello 2002; Baigún & Ferriz 2003). Nuestros resultados son similares a los de trabajos anteriores realizados en el río Negro (AIC-UNS 1995; Almirón et al. 1997). La principal diferencia con respecto al primero de los trabajos consiste en la aparición de dos especies de origen exótico, la coridora y la carpa común. Este dato es novedoso ya que no han sido citadas hasta el momento para este cuerpo de agua. La aparición de mojarras, puyen chico, larvas de lamprea y madrecitas en nuestro trabajo, dada por el uso de artes de pesca complementarios, brindó una imagen más completa de la composición de la ictiofauna, permitiendo reafirmar los resultados encontrados por Almirón et al. (1997), particularmente con respecto a las especies de pequeño porte.
Las especies nativas fueron las más abundantes a lo largo del río durante todo el año, considerando los dos métodos de pesca. Teniendo en cuenta las capturas con redes de enmalle, se distinguieron patrones particulares de abundancia para las distintas especies. El pejerrey patagónico fue más abundante hacia Guardia Mitre, la perca de boca chica en los cursos superior y medio del río y la perca bocona en el curso medio. La trucha arco iris fue mas abundante en el curso superior, resultado coincidente con el hallado por AIC - UNS (1995). La aparición de truchas arco iris en el curso inferior podría deberse a las siembras realizadas por organismos provinciales en los últimos años. Si bien se han practicado siembras regulares de esta especie a partir de 1920 y hasta la actualidad (Macchi 2004), la distribución observada podría estar más asociada a una dispersión natural desde las sub-cuencas, ubicadas aguas arriba, de los ríos Limay y Neuquén. Las exigencias en cuanto a calidad ambiental necesarias para el establecimiento de poblaciones de salmónidos auto-sostenidas en el tiempo son de difícil cumplimiento en el ecosistema del río Negro. Las más relevantes son la escasez de lechos aptos para desove y las altas temperaturas durante el verano, afectando en forma desfavorable el desarrollo embrionario y crecimiento de alevinos. La carpa común tuvo una presencia relativamente importante en Guardia Mitre, apareciendo hasta Choele Choel. Sin embargo, cabría esperar una expansión en su distribución hacia el curso superior en los próximos años dada la fácil adaptación de esta especie a distintas condiciones ambientales (Kohen 2004). La carpa ha sido citada para numerosos cuerpos de agua de la planicie pampeana, incluyendo el río Colorado, límite entre las provincias de Río Negro y la Pampa (Liotta 2006). Se cree que allí fue introducida a mediados de 1980 para control de la vegetación acuática (Cazzaniga 1981). Es posible que la aparición de esta especie en el río Negro se deba a las mismas causas. La lisa apareció en los meses de verano y primavera hasta el curso medio, resultado coincidente con el de AIC - UNS (1995). Estos movimientos podrían estar relacionados con los hábitos alimentarios de la especie, descartándose movimientos reproductivos ya que la lisa desova en aguas estuarinas o con altas salinidades (Lasta 1995). Por último, el bagre aterciopelado y el pejerrey bonaerense aparecieron ocasionalmente con abundancias poco importantes en las localidades de Allen y Guardia Mitre respectivamente, resultado también coincidente con el estudio de AIC - UNS (1995).
En las capturas con artes complementarios, las especies más abundantes fueron la madrecita, los juveniles de pejerreyes y percas, y la coridora. Estas especies aparecieron en todos los sitios de muestreo, particularmente durante la época de estiaje y variaron su importancia en las capturas de acuerdo con el sitio de muestreo. El puyen chico fue capturado solo en Allen, aunque ha sido citado para el curso inferior y medio del río por Azpelicueta et al. (1996). Es una especie de aparición común en los cuerpos de agua norpatagónicos y constituye un recurso alimentario importante para peces ictiófagos (Macchi et al. 1999). Las larvas de lamprea se capturaron en Conesa en números muy bajos, aunque la especie es de aparición común en el río Negro e incluso ha sido citada en el curso inferior del río Limay (Baigún et al. 2002). La lamprea realiza migraciones reproductivas de adultos desde el mar para desovar en aguas dulces (Cushing et al. 1995). A diferencia de lo observado en las capturas con redes agalleras, no fue posible hallar patrones particulares de distribución para los peces de pequeño porte. El bajo número de capturas hacia el otoño e invierno podría estar asociado a un cambio de hábitat producido por el incremento en el caudal del río, utilizándose los brazos y canales secundarios como refugio, o a una disminución en la efectividad del arte de pesca dado por ese incremento. Esta última idea no debería subestimarse, dado que la complejidad estructural del ambiente tiene una relación directa con el éxito de las capturas (Harvey & Cowx 1996). Teniendo en cuenta esta idea, es importante considerar la composición específica de la comunidad íctica que muestra la presencia de percas y pejerreyes juveniles a lo largo del río, lo que indicaría la existencia de condiciones apropiadas para la reproducción de estas especies en todo el curso del río.
Los índices ecológicos, en conjunto con las abundancias relativas, permitieron definir características de la comunidad de peces a lo largo del río. En particular, Guardia Mitre mostró los mayores índices de dominancia y los menores índices de equitatividad y diversidad. El resto del río mostró valores similares entre los sitios de muestreo. Los altos valores de dominancia y los coincidentes resultados observados en los bajos índices de equitatividad y diversidad estarían indicando condiciones ambientales que favorecen particularmente a una o dos especies (Camargo 1992). En general estas diferencias podrían atribuirse a una distribución heterogénea de la ictiofauna en los sistemas fluviales. Estas distribuciones están principalmente influenciada por factores físicos (forma del canal, pendientes de los taludes, ancho del valle) y biológicos (vegetación riparia, productividad acuática) (Naiman & Bilby 1998), aunque no debe descartarse el efecto modificador de las siembras efectuadas por el hombre. Nuestros resultados coinciden parcialmente con la idea de continuo del río de Vannote et al. (1980) y Welcomme (1985) con respecto al aumento de la diversidad y de la abundancia de peces en el sentido de la corriente. La dominancia de una o dos especies en el curso inferior es llamativa, pues no coincide con lo propuesto por los autores mencionados anteriormente. La práctica de liberación de peces, tanto oficial como accidental, puede influenciar la interpretación de estos índices, por lo que deben considerarse cuidadosamente.
Un aspecto importante en la descripción de la comunidad de peces del río Negro es la presencia de especies de origen exótico, dado que la composición de la comunidad podría estar determinada, entre otras causas, por la interacción entre especies nativas y exóticas (Taylor et al. 1984). Los valores de C.I.Z. indican una proporción relativamente equilibrada entre los grupos de ambos orígenes, aunque el componente "exótico" difiere a lo largo del río. La fauna de origen brasílico tiene en general una mayor importancia que las especies del hemisferio Norte. Sin embargo, estas diferencias en la distribución son de difícil interpretación ya que podrían deberse a causas ambientales y/o diferencias en los procesos de dispersión desde los sitios de introducción. (Ringuelet 1975; Baigún et al. 2002). La escala temporal de dispersión y establecimiento de una especie es difícil de medir y principalmente depende de la habilidad de la especie para ocupar nuevos ambientes (D'Antonio 1993). Por otra parte, la situación actual en el río Negro puede considerarse dinámica, en función de las respuestas de las comunidades ícticas a la dispersión de las especies exóticas registradas en los últimos años, que dependerán principalmente de características de la especie introducida y de la estructura de la comunidad afectada (Ross 1991).
Los estudios de alimentación permitieron determinar relaciones tróficas entre las especies presentes. Estas muestran amplios espectros tróficos, aunque hay cierto grado de preferencia en el consumo de alimento. Los grupos tróficos individualizados explotan una importante variedad de recursos, que incluyen desde el detrito hasta los peces. En el río Negro el rol de depredador tope es cumplido por la perca bocona, el micro y macrozoobentos son consumidos por la mayoría de las especies, el rol de planctófago es cumplido por las especies de menor tamaño que viven en las zonas de aguas tranquilas. Aparece un consumidor de macrófita, la carpa, y la materia orgánica es incluida en la dieta de varias especies, principalmente en los cursos medio e inferior del río. Estos grupos tróficos fueron descriptos por otros autores en distintos cuerpos de agua de la cuenca del río Limay (Ferriz 1988, 1989, 1993/1994; Macchi et al. 1999; Milano et al. 2002; Macchi 2004). En estos ambientes el rol de depredador tope es cumplido principalmente por la perca bocona y por la trucha marron (Salmo trutta), en tanto que la perca de boca chica, las truchas arco iris y la trucha de arroyo (Salvelinus fontinalis), el pejerrey patagónico y los galáxidos son principalmente depredadores de micro y macrozoobentos. Estacionalmente los adultos de pejerrey se vuelven zooplanctófagos y en los embalses incorporan gran cantidad de algas filamentosas. Finalmente, solo Galaxias platei (puyen grande) incorpora materia orgánica a su alimentación.
Nuestros resultados indican solapamientos de dietas principalmente entre especies nativas y exóticas, tanto en peces grandes como en pequeños. El solapamiento en las dietas podría resultar en competencia por recursos, dependiendo de la abundancia de presas (Matthews 1998). Este hecho resalta la importancia de las interrelaciones entre los integrantes de la comunidad íctica cuando ésta es modificada a través de la introducción de nuevas especies. La trucha arco iris y la carpa común han tenido influencias negativas en otros lugares del mundo donde han sido introducidas (McDowell 2003; Kohen 2004). Aunque las abundancias relativas de estas especies son bajas, no es posible descartar una interacción negativa más fuerte al ampliarse el rango de distribución actual o aumentar su importancia relativa.
La piscivoría aparece como una relación importante en la comunidad de peces del río Negro y ha sido observada en otros cuerpos de agua de la región Patagónica (Macchi et al. 1999; Macchi et al. 2007). La mayoría de las especies incluyen peces en sus dietas, aunque con importancias relativas diferentes. La perca bocona se comporta como piscívora casi estricta, en tanto que la perca de boca chica, el pejerrey patagónico o la trucha arco iris consumen peces facultativamente. Estos resultados coinciden con los hallados por Macchi et al. 1999. Es importante destacar que la mayoría de las especies son consumidas, resaltando la importancia de la piscivoría como relación ecológica en la comunidad. Los peces grandes podrían cumplir un importante rol en la regulación de la composición de dicha comunidad ya que la depredación es un evento de inmediata repercusión en los números poblacionales de las especies presas (Moyle & Light 1996).
De la novedosa aparición de dos especies nuevas para este río, la coridora parece estar presente en todo el curso a diferencia de la carpa común que sólo apareció hasta el sector medio. La coridora ha sido observada en canales interurbanos de varias localidades en la zona del alto valle (Alvear, obs. pers.), demostrando la gran adaptación de la especie a las condiciones ambientales de la zona. Si bien fue citada para el río Limay (Baigún et al. 2002) no se encontraron referencias de su presencia en el río Negro anteriores a este trabajo. Por su parte la carpa, distribuida ampliamente en todo el mundo (Welcomme 1988), es encontrada en varias provincias argentinas (Liotta 2006). En Patagonia ha sido mencionada para el río Colorado por este último autor. La presencia de coridora y carpa en el río Negro no incrementa el número de especies citadas para la Patagonia (Pascual et al. 2002; Baigún & Ferriz 2003) pero amplía los rangos de distribución de las mismas hacia el sur. El rango de distribución de una especie depende de factores ambientales, fisiológicos y humanos. Los factores ambientales y fisiológicos se relacionan con los niveles de tolerancia físico-químicos y la eficacia del método de dispersión natural de cada especie, entre otras razones (Moyle & Light 1996). El traslado de especies fuera de su rango de distribución natural hecho por el hombre es denominado antropocoria. Esta acción ha sido citada como una de las actividades ecológicas más dañinas realizada por el hombre (Lodge 1993; Wilcove et al. 1998).
La comunidad de peces del río Negro es el resultado de la condición geográfica ecotonal del río (López 2001) y de la fuerte influencia de las actividades humanas asociada a este curso de agua. La antropocoria, tanto accidental como intencional, podría tener un importante efecto sobre la ictiofauna. Esta actividad no siempre está documentada y muchas veces es llevada a cabo sin profundizar el análisis de necesidades sociales o económicas, ni el impacto sobre el ecosistema. Existen escasos estudios en Patagonia que analizan la interacción entre las especies de peces exóticos y nativos (Ortubay & Cussac 2000; Macchi et al. 1999; Macchi 2004; Macchi et al. 2007). Estas interacciones cobran especial importancia considerando la baja diversidad de especies para la región patagónica y debe profundizarse su estudio para reducir o evitar efectos negativos, como los hallados en otros ambientes, que han llevado incluso a la extinción de especies (Brown & Moyle 1997). El estudio del impacto generado por las introducciones de peces y de la evolución de las mismas permitirá entender el funcionamiento de la comunidad íctica. Este conocimiento, junto con el desarrollo de acciones efectivas para evitar nuevas introducciones, será necesario para proteger, preservar o explotar la valiosa ictiofauna del río Negro.
AGRADECIMIENTOS
Deseamos agradecer a M. Bello y A. López Cazorla por el aporte bibliográfico, a L. Protogino del Instituto de Limnología R. Ringuelet de la Plata por su colaboración en la determinación taxonómica de mojarras. A J. De Giusto y M. Uguccioni por su invalorable apoyo y colaboración en las tareas de campo y por último al Centro Regional Universitario Bariloche. Este estudio fue financiado por la Autoridad Interjurisdiccional de las cuencas de los ríos Limay, Neuquén y Negro.
BIBLIOGRAFÍA
AIC - UNS. 1995. Relevamiento de la ictiofauna del Río Negro. Convenio complementario entre la Autoridad Interjurisdiccional de las cuencas de los ríos Limay, Neuquén y Negro. Universidad Nacional del Sur. Informe Técnico. 70 pp. [ Links ]
ALMIRÓN, A; M AZPELICUETA; JR CASCIOTTA & A LÓPEZ CAZORLA. 1997. Ichthyogeographic boundary between the Brazilian an Austral subregions in South América. Biogeographic, 73(1):23-30. [ Links ]
ARRATIA, G; MB PEÑAFORT & S MENU MARQUE. 1983. Peces de la región sureste de los Andes y sus probables relaciones biogeográficas actuales. Desert, 7:48-107. [ Links ]
AZPELICUETA, MM; A ALMIRÓN; A LÓPEZ-CAZORLA & J CACIOTTA. 1996. Geographical distribution of Galaxias maculatus (Jenyns) (Pisces: Osmeriformes: Galaxidae) in Patagonia. Biogeographic, 72:157-160. [ Links ]
BAIGÚN, C & R FERRIZ. 2003. Distribution patterns of native freshwater fishes in Patagonia (Argentina). Org. Divers. Evol., 3:151-159. [ Links ]
BAIGÚN, C; G LÓPEZ; A DOMÁNICO; R FERRIZ; S SVERLIJ & RD SCHENKE. 2002. Presencia de Corydorass paleatus (Jenyns, 1842), una nueva especien brasílica en el norte de la Patagonia (Río Limay) y consideraciones ecológicas relacionadas con su distribución. Ecol. Austral, 12:41-48. [ Links ]
BELLO, MT. 2002. Los peces autóctonos de la Patagonia Argentina. Distribución natural. Cuadernos Universitarios. Universidad Nacional del Comahue. Centro Regional Universitario Bariloche. [ Links ]
BIANCO, PG. 1990. Proposta di impiego di indici e di coefficienti per la valutazione dello stato di degrade dell'ittiofauna autoctona delle acque dolci. Rivista di Hidrobiología, 29: 131-149. [ Links ]
BROWN, LR & PB MOYLE. 1997. Invading species in the Eel River, California: successes, failures, and relationships with resident species. Environ. Biol. Fishes, 49:271-291. [ Links ]
CAMARGO, JA. 1992. Structural and trophic alterations in macrobenthic communities downstream from a fish farm outlet. Hidrobiología, 242:41–49 [ Links ]
CAZZANIGA, NJ. 1981. Caracterización químico-faunística de canales de drenaje del Valle Inferior del Río Colorado. Ecosur, 8:24-46. [ Links ]
CUSHING, EC; KW CUMMINS & GW MINSHALL. 1995. Ecosystem of the World. River and Stream Ecosystem, 22:257-293. [ Links ]
CUSSAC, VE; D RUZZANTE; S WALDE; PJ MACCHI; V OJEDA; MF ALONSO ET AL. 1998. Body shape variation of three species of Percichthys in relation to their coexistence in the Limay river basin, in Northern Patagonia. Environ. Biol. Fishes, 53:143-153. [ Links ]
D'ANTONIO, CM. 1993. Mechanisms controlling invasion of coastal plant communities by the alien succulent Carpobrotus eludis. Ecology, 74:83-95. [ Links ]
DE APARICIO, FO & HA DIFRIERI. 1958. La Argentina. Suma de geografía. Peuser. Bs.As, Argentina. [ Links ]
ELVIRA, B. 1995. Native and exotic freshwater fishes in Spanish river basins. Freshw. Biol., 33 (1):103–108. [ Links ]
FERRIZ, RA. 1988. Relaciones tróficas de trucha marrón Salmo fario Linné y trucha arco iris Salmo gairdneri Richardson, (Osteichthyes, Salmoniformes) en un embalse norpatagónico. Stud. Neotrop. Fauna Environ., 23:123-131. [ Links ]
FERRIZ, RA. 1989. Alimentación de Percichthys colhuapiensis (Mac Donagh, 1955) y P. trucha (Girard, 1854) (Osteichthyes, Percichthyidae) en el embalse Ramos Mexía, provincia del Neuquén, Argentina. Iheringia, 69:109-116. [ Links ]
FERRIZ, RA. 1993/1994. Algunos aspecto de la dieta de cuatro especies ícticas del río Limay (Argentina). Revista de Ictiología, 2/3:1-7. [ Links ]
HARVEY, J & IG COWX. 1996. Electric fishing for the assessment of fish stocks in large rivers. [1] Pp. 11-26 in: IG Cowx (ed). Stock assessment in inland fisheries. 1º edn. Fishing New Books. [ Links ]
KOHEN, J. 2004. Carp (Cyprinus carpio) as a powerful invader in Australian waterways. Freshw. Biol., 49:882-894. [ Links ]
LASTA, CA. 1995. La bahía Sanborombón; zona de desove y cría de peces. Tesis nº 636. Facultad de Ciencias Naturales y Museo (UNLP). Argentina. 304 pp. [ Links ]
LIOTTA, J. 2006. Distribución geográfica de los peces de aguas continentales de la República Argentina. ProBiota, nº3. 701 pp. [ Links ]
LODGE, DM. 1993. Biological invasions: lessons for ecology. Trends Ecol. Evol., 8:133-137. [ Links ]
LÓPEZ, HL. 2001. Estudio y uso sustentable de la biota austral: Ictiofauna continental Argentina. Revista Cubana de Investigación Pesquera. Suplemento Especial. 35pp. [ Links ]
LÓPEZ, HL; AM MIQUELARENA & J PONTE GÓMEZ. 2005. Biodiversidad y distribución de la ictiofauna mesopotámica. Temas de la biodiversidad del litoral fluvial argentino. Miscelanea, 14:311-354. [ Links ]
LÓPEZ-ARBARELLO, A. 2004. Taxonomy of the genus Percichthys (Perciformes: Percichthyidae). Ichthyol. Explor. Freshwater, 4:331-350. [ Links ]
MACCHI, PJ. 2004. Mecanismos de comportamiento vinculados a respuestas de escape y ataque entre Galaxias maculatus y sus predadores. Tesis doctoral, Universidad Nacional del Comahue. Bariloche, Argentina. [ Links ]
MACCHI, PJ; V CUSSAC; MF ALONSO & MA DENGRI. 1999. Predation relationships between introduced salmonids and native fish fauna in lakes end reservoirs of northern Patagonia. Ecol. Freshw. Fish., 8:227-236. [ Links ]
MACCHI, PJ; MA PASCUAL & PH VIGLIANO. 2007. Differential piscivory of the native Percichthys trucha and exotic salmonids upon the native forage fish Galaxias maculatus in Patagonian Andean lakes. Limnologica, 37:76-87. [ Links ]
MATTHEWS, WJ. 1998. Patterns in freshwater fish ecology. 2nd edn. Chapmann & Hall. Norwall, Massachusetts. 756 pp. [ Links ]
MCDOWELL, RM. 2003. Impacts of introduced salmonids on native galaxiids in New Zealand upland streams, a new look at an old problem. Trans. Am. Fish. Soc., 132:229-238. [ Links ]
MILANO, D; VE CUSSAC; PJ MACCHI; DE RUZZANTE; MF ALONSO ET AL. 2002. Predator associated morphology in Galaxias platei in Patagonian lakes. J. Fish Biol., 61/1:138-156. [ Links ]
MOYLE, PB & T LIGHT. 1996. Biological invasions of freshwater: empirical rules and assembly theory. Biol. Conserv., 78:149-161. [ Links ]
NAIMAN, RJ & RE BILBY. 1998. River ecology and management. Lesson from the Pacific Coastal Ecoregion. Springer- Verlag. New York, Inc. 705 pp. [ Links ]
NORUSIS, MJ. 1986. SPSS/PC+ Advanced Statistics. SPSS Inc. Chicago. 328 pp. [ Links ]
ORTUBAY, S & V CUSSAC. 2000. Threatened fishes of the world: Gymnocharacinus bergii Steindachner, 1903 (Characidae). Environ. Biol. Fishes, 58:144. [ Links ]
PASCUAL, M; PJ MACCHI; J URBANSKI; F MARCOS; C RIVA ROSSI; M NOVARA ET AL. 2002. Evaluating poential effects of exotic freshwater fish from imcomplete species presence-absence data. Biol. Invasions, 4:101-113. [ Links ]
PIELOU, EC. 1975. Ecological diversity. Wiley. Nueva York. 165 pp. [ Links ]
RINGUELET, RA. 1975. Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur, 2(3):122 pp. [ Links ]
RINGUELET, RA & RH ARAMBURU. 1961. Peces argentinos de agua dulce. Claves de reconocimiento y caracterización de familias y subfamilias, con glosario explicativo. AGRO. Publicación técnica. Año III nº7. 98 pp. [ Links ]
ROSS, ST. 1991. Mechanisms structuring stream fish assemblages: are there lessons from introduced species? Environ. Biol. Fishes, 30:359-368. [ Links ]
RUZZANTE, DE; S WALDE; V CUSSAC; P MACCHI; M ALONSO ET AL. 2003. Resource polymorphism in a Patagonian fish Percichthys trucha (Percichthyidae): phenotypic evidence for interlake pattern variation. Biol. J. Linn. Soc., 78: 497-515 [ Links ]
SCHÖENER, TW. 1970. Non-syncronous spatial overlap of Lizards in patchi habitats. Ecology, 51:408-418. [ Links ]
TAYLOR, JN; J COURTENAY & JA MCCANN. 1984. Known impacts of exotic fishes in the continental United States. Pp 332-373 in: WR Courtney, Jr & JR Stauffer, Jr. (ed). Distribution, Biology and Management of Exotic Fishes. John Hopkins University Press. Baltimore. [ Links ]
VANNOTE, R; G MINSHALL; K CUMMINS; JR SEDELL & CE CUSHING. 1980. The river continuum concept.Canadian J. Fish Aquat. Sci., 37:130-137. [ Links ]
WALLACE, RK. 1981. An assesment of diet-overlap indexes. Trans. Am. Fish. Soc., 110:72-76 pp. [ Links ]
WELCOMME, RL. 1985. River fisheries. FAO Fisheries Technical Paper 262. 330 pp. [ Links ]
WELCOMME, RL. 1988. International Introductions of Inland Aquatic Species. FAO Fisheries Technical paper 294. 318 pp. [ Links ]
WILCOVE, DS; D ROTHSTEIN; J DUBOW; A PHILLIPS & E LOSS. 1998. Quantifying threats to imperiled species in the United States. BioSciense, 48:607. [ Links ]

