










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.17 n.2 Córdoba jul./dic. 2007
TRABAJOS ORIGINALES
La riqueza de artrópodos bentónicos en una cuenca endorreica subtropical (Tucumán, Argentina)
Leticia M Mesa1, * & Hugo R Fernández2
1. CONICET, Instituto Miguel Lillo, San Miguel de Tucumán, Tucumán, Argentina.
2. CONICET, Fac. Ciencias Naturales e Instituto Miguel Lillo, San Miguel de Tucumán, Tucumán, Argentina.
* CONICET. Instituto Miguel Lillo. Miguel Lillo 205. 4000. San Miguel de Tucumán. Tucumán. Argentina.Email: leticiamesa@yahoo.com.ar
Recibido: 25 de enero de 2006;
Fin de arbitraje: 23 de mayo de 2007;
Revisión recibida: 29 de junio de 2007;
Aceptado: 7 de julio de 2007.
Este trabajo fue arbitrado durante el proceso editorial de Marcelo Cassini actuando Alfredo Salibian como co-editor.
RESUMEN. La disminución de la biodiversidad de los ecosistemas dulceacuícolas resulta en la necesidad de desarrollar bases empíricas que describan los patrones de distribución de organismos acuáticos. El objetivo de este trabajo fue estimar la riqueza de artrópodos bentónicos de subcuencas de la provincia de Tucumán. Utilizamos diferentes estimadores (Sobs, ICE, Jack1 y Cole) para estudiar la riqueza de artrópodos de cuatro subcuencas de la cuenca río Salí-Dulce. La información tratada proviene de una base de datos que consta de 2300 registros de macroinvertebrados acuáticos hasta el momento. La comparación de riqueza (Jack1) entre las dos subcuencas mejor muestreadas (>300 registros) no mostró diferencias. Los datos (de presencia/ausencia) fueron explorados mediante un análisis multivariado que reveló que el número total de ríos y su caudal explicaron la mayor proporción de la variación documentada. Los análisis demostraron las diferencias en relación al conocimiento de la comunidad bentónica de las subcuencas estudiadas.
Palabras clave: Biodiversidad; Yungas; Sobs; ICE; Jack1; Cole; Ríos de montaña.
ABSTRACT. Richness of arthropods benthic fauna in a subtropical endorheic basin (Tucumán, Argentina): The reduction of biodiversity in freshwater ecosystems makes necessary the development of empirical basis that describe the patterns of distribution of aquatic organisms. With this purpose, the richness of arthropods from benthic zones was studied using different estimators: Sobs, ICE, Jack1 and Cole. We used a data base of aquatic macroinvertebrate fauna from streams of Yungas biodiversity project including 2300 enters. The richness comparison (Jack1) between the better sub-basins sampled (>300 enters) did not detect differences between them. Besides, a multivariate analysis revealed that the total number of streams and discharge were the variables that accounted for most variation. This study showed the differences in knowledge of the benthic communities from the studied sub-basins.
Keywords: Biodiversity; Yungas; Sobs; ICE; Jack1; Cole; Mountain streams.
INTRODUCCIÓN
El rápido crecimiento de la población y la explotación de los recursos naturales en América del Sur conlleva una rápida disminución de la biodiversidad de los ecosistemas acuáticos. Se reconoce hoy que existen extinciones causadas indirectamente por la destrucción de habitats, así como por alteraciones profundas derivadas directamente de la actividad humana (Allan & Flecker 1993). La falta de bases empíricas que describan los patrones de distribución de la diversidad de insectos acuáticos resulta llamativa, ya que los ecosistemas dulceacuícolas se encuentran entre los ambientes más amenazados de la tierra (Vinson & Hawkins 1998). Además, los escenarios de diversidad pronosticados para los ecosistemas acuáticos continentales aparecen desalentadores (Sala et al. 2000).
La ecoregión de las Yungas se extiende en Argentina entre los 22° y 28° de latitud Sur, con una superficie de aproximadamente cuatro millones de hectáreas. Representa un 9% de la superficie total del Noroeste de Argentina (NOA) constituyendo, junto con la selva misionera, las dos áreas más biodiversas de la Argentina (Grau & Brown 2000). Esta región biogeográfica es además el área donde se originan algunos de los ríos más importantes de Argentina, pero su biodiversidad se encuentra amenazada actualmente por diferentes tipos de impactos antrópicos. La provincia de Tucumán, ubicada en el centro del NOA, no está libre de estos efectos (Dominguez & Fernández 1998; Fernández et al. 2002). Dicha región se caracteriza por poseer un régimen monzónico de lluvias en verano que son drenadas por numerosos ríos desde el oeste de su territorio hasta la cuenca Salí-Dulce.
Una forma de prevenir las pérdidas de biodiversidad y tomar medidas para su protección, y quizá manejo, es entender los patrones de riqueza de especies en el espacio (Wu & Gao 1995; Ward & Tockner 2001). En ese sentido, tomamos la cuenca superior del sistema Salí-Dulce como unidad regional de análisis, y seleccionamos cuatro subcuencas para su estudio. Teniendo en cuenta la diferente intensidad de muestreo en las unidades elegidas, el objetivo de este trabajo es estimar la riqueza de artrópodos bentónicos de subcuencas de la provincia de Tucumán. A través de estimadores no paramétricos (Melo & Froehlich 2001), comparamos los valores de riqueza de las subcuencas y exploramos la influencia de diferentes variables sobre las unidades elegidas (ter Braak 1995).
MATERIALES Y MÉTODOS
Se seleccionaron cuatro subcuencas de la cuenca alta del sistema Salí-Dulce ubicadas en la provincia de Tucumán (Fig. 1), sobre la base del estado de conocimiento de la comunidad bentónica en la provincia (Dominguez & Ballesteros Valdez 1992; Fernández & Dominguez 2001; Fernández et al. 2001). Las subcuencas seleccionadas fueron: subcuenca río Calera (SCL), río Lules (SCL), río Medinas (SCM) y río Marapa (SCMa). Se eligieron siete órdenes de artrópodos: Ephemeroptera, Trichoptera, Prostigmata (Acari), Coleoptera, Lepidoptera, Megaloptera y Diptera (excluído Chironomidae) por tener identificación del más bajo nivel alcanzable. La zona muestreada en cada caso se encuentra en buen estado de conservación, registrándose sólo alguna contaminación difusa de poblaciones pequeñas (<1000 habitantes) y efectos provenientes del uso del suelo (Dominguez & Fernández 1998; Fernández et al. 2002).
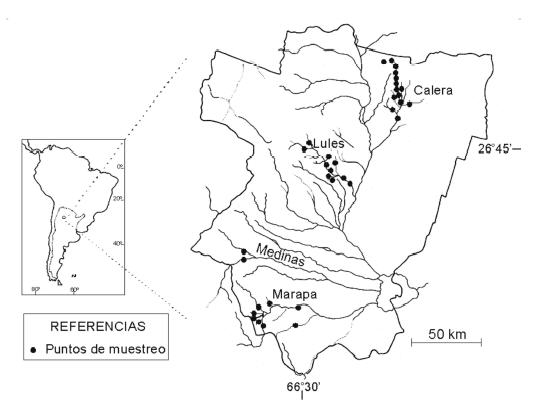
Figura 1. Mapa de la provincia de Tucumán con la ubicación de las subcuencas, los sitios de muestreo y su posición relativa en Sudamérica y Argentina.
Figure 1. Map of Tucumán province including the location of the sub-basins, the sampling sites and its relative position in South America and Argentina.
La riqueza de cada subcuenca se estimó usando Estimates 5.0.1 (Colwell 1997), seleccionando la opción por defecto. Se utilizaron los datos de presencia/ausencia de los órdenes de artrópodos elegidos (Apéndice 1) con identificaciones de especies confirmadas. La información se obtuvo de una base de datos de macroinvertebrados bentónicos de las Yungas de Argentina, con 2300 registros al momento del análisis. La información referente a la SCM no pudo ser procesada por el programa a causa del bajo número de sitios de muestreo. Asimismo, se seleccionaron cuatro estimadores: el estimador de especies observadas (Sobs), el estimador de Rarefacción o Coleman (Cole), el estimador basado en la incidencia de las especies (ICE) y el estimador de Jacknife (Jack1) teniendo en cuenta sus ventajas (Melo & Froehlich 2001). Mediante la prueba de t Student se comparó la riqueza estimada por Jack1 para ocho sitios de muestreo, con α= 0.05. La comparación de t se efectúo con un valor tabulado de 2.365, para siete grados de libertad. Las curvas de los estimadores y su relación con la riqueza observada (Sobs) se interpretaron siguiendo a Heyer et al. (1999) y Melo & Froehlich (2001). Para explorar la relación entre la distribución de los taxones y diferentes variables propias de las subcuencas (Tabla 2), se empleó un Análisis de Correspondencia Canónica (ACC) (programa PC-ORD 4.25). Área (A), número total de ríos (N) y caudal (Q) fueron las características elegidas entre 17 disponibles mediante un Análisis de Componentes Principales (ACP). La influencia de estas variables sobre los ejes se evaluó mediante los valores de correlación intraset del ACC según ter Braak (1995).
Tabla 2. Variables y códigos utilizados en el ACC. SCC = subcuenca río Calera, SCL = subcuenca río Lules, SCM = subcuenca río Medinas, SCMa = subcuenca río Marapa.
Table 2. Variables and codes used in CCA. SCC = Calera river sub-basin, SCL = Lules river sub-basin, SCM = Medinas river sub-basin, SCMa = Marapa river sub-basin.

RESULTADOS
Se seleccionaron 960 registros de la base de datos relacionados con las subcuencas de interés y grupos de artrópodos elegidos (Tabla 1). Del total, 495 registros pertenecían a SCC con 73 especies. El número de registros correspondientes a SCL fue de 307 con 71 especies. A SCM le correspondieron 109 registros con 34 especies, mientras que la SCMa estuvo representada por 49 registros con 24 especies.
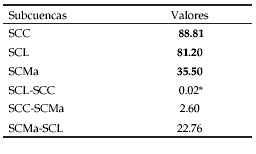
Tabla 1. Valores de riqueza estimada por Jack1 para ocho sitios de muestreo (en negrita) y resultados derivados de la prueba de t Student. (*P < 0.05). SCC = Subcuenca río Calera, SCL = Subcuenca río Lules, SCMa = Subcuenca río Marapa.
Table 1. Jack1 richness estimation for eight sampled sites (in bold) and results of t Student test. (*P < 0.05). SCC = Calera river sub-basin, SCL = Lules river sub-basin, SCMa = Marapa river sub-basin.
En relación con la distribución de la riqueza por grupo y subcuenca (Fig. 2), se destacó la riqueza de hidrácaros en SCC y SCL, y también resultó notable la ausencia del orden Lepidoptera en SCL y SCMa.
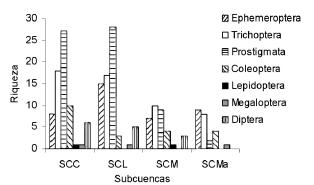
Figura 2. Importancia relativa de cada orden (como número de especies) en cada subcuenca. SCC = subcuenca río Calera, SCL = subcuenca río Lules, SCM = subcuenca río Medinas, SCMa = subcuenca río Marapa.
Figure 2. Relative importance of each order (as species number) in each sub-basin. SCC = Calera river sub-basin, SCL = Lules river sub-basin, SCM = Medinas river sub-basin, SCMa = Marapa river sub-basin.
El análisis de las curvas producidas por los diferentes estimadores de riqueza para la SCC (Fig. 3c) mostró que la curva de Cole se asemeja en gran medida a la estimación de Sobs; el estimador Jack1 sobreestimó la riqueza de especies total, mientras que ICE subió de un modo escalonado. Por otra parte, en SCL (Fig. 3a) el estimador Jack1 estimó con menor rango de error la verdadera riqueza, ya que la curva se acerca en gran medida a la de Sobs, mientras que ICE tendió a aproximarse a Sobs en los últimos sitios de muestreo. En SCMa (Fig. 3b) el estimador Jack1 sobreestimó la riqueza de especies, mientras que el estimador ICE describió una curva singular, manteniéndose lejos de Sobs.
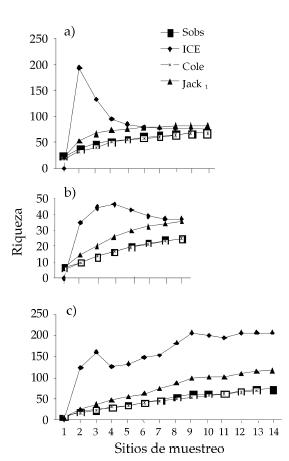
Figura 3. Estimación de la riqueza de artrópodos bentónicos para las diferentes subcuencas mediante cuatro estimadores no paramétricos: Sobs = estimador de especies observadas, ICE = estimador basado en la incidencia de las especies, Cole = estimador de rarefacción o Coleman y Jack1 = estimador de Jacknife 1. a) = subcuenca río Lules, b) = subcuenca río Marapa, c) = subcuenca río Calera.
Figure 3. Richness estimation of benthic arthropods for the different subbasins using four non-parametric estimators: Sobs = Observed-based coverage estimator, ICE = Incidendent-based coverage estimator, Cole = Coleman estimator, Jack1 = First order Jackknife estimator. a) = Lules river sub-basin, b) = Marapa river sub-basin, c) = Calera river sub-basin.
Las dos subcuencas mejor muestreadas (>300 registros, SCC y SCL) no presentaron diferencias en la riqueza estimada por Jack1 para ocho sitios de muestreo (P < 0.05) (Tabla 1). Por otra parte, la riqueza estimada fue distinta entre las SCMa y SCL, SCC y SCMa según la prueba de t Student (Tabla 1).
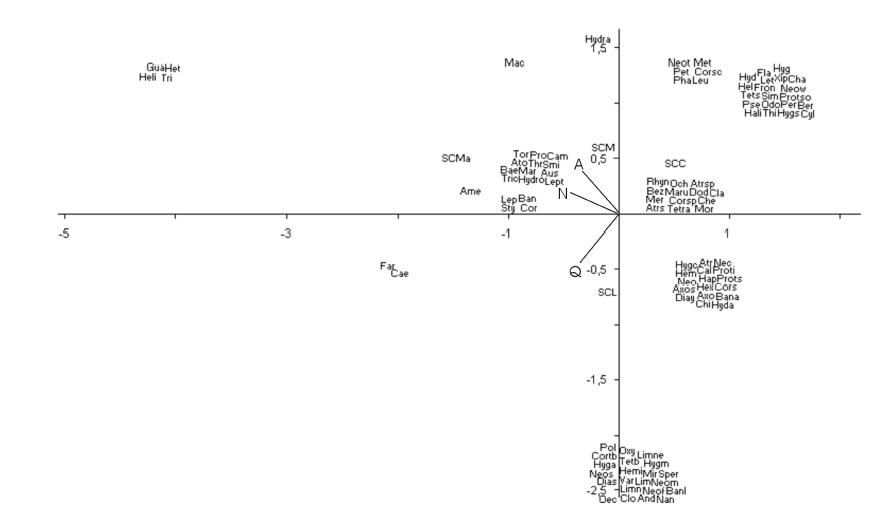
Se seleccionaron los dos primeros ejes del ACC: el eje 1 (λ = 0.35) explicó 39.7% de la variación total, mientras que el eje 2 (? = 0.34) explicó 37.8 %. En un extremo del eje 1 se ubicó SCMa (cuadrante superior izquierdo; Fig. 4) definida por Triplectides sp., Heterelmis sp., Guajirolus sp. y Helichus sp. En el otro extremo del mismo eje, se ubicó SCC (cuadrante superior derecho), descripta por un conjunto de 20 taxones (Fig. 4). Por otra parte, la SCL y SCM aportaron poco a la explicación de este eje. La variable N explicó el gradiente existente en el eje 1 (r2 = 0.75). En el análisis del eje 2 se observa una clara diferenciación entre la SCL y SCM definida por Q (r2 = 0.76), y en menor medida por A (r2 = 0.68). Un conjunto de 22 taxones caracterizó a la SCL (cuadrante inferior, Fig. 4), mientras que en el otro extremo (cuadrante superior izquierdo) SCM resultó descripta por solo un taxón (Hydramara argentina).
Figura 4. Ordenamiento de las cuatro subcuencas y 103 taxones mediante análisis de correspondencia canónica (ACC) (códigos en Tabla 1). A = área, N = número total de ríos y Q = caudal.
Figure 4. Order of the four sub-basins and 103 taxa using canonical correspondence analysis (CCA) (codes in Table 1). A = area, N = total number of streams and Q = discharge.
DISCUSIÓN
Los bajos valores de riqueza de las SCM y SCMa se relacionaron con la falta de muestreos, posiblemente debido a la lejanía y dificultad de acceso a estas subcuencas. Sin embargo, la magnitud de ambas subcuencas, su heterogeneidad espacial y el bajo impacto antrópico nos llevan a pensar que, con estudios intensivos, sus valores de riqueza superarán a los de las unidades mejores conocidas.
La riqueza relativa por taxón en cada sub-cuenca reveló la importancia de los hidrácaros, un grupo que frecuentemente no es considerado en estudios de comunidades bentónicas de ríos y arroyos (Fernández 2003). Basándonos en las curvas de los estimadores podemos decir que en SCC (Fig. 3c) la riqueza real es superior a la observada. Las curvas de Sobs y Jack1 son aproximadamente lineales con tendencia ascendente y, además, la separación entre la curva de Sobs e ICE es grande. Esto sugiere que se necesitaría un mayor número de muestreos para una riqueza más ajustada a la real (Heyer et al. 1999) ya que la heterogeneidad en esta unidad hidrográfica es alta (Wu & Gao 1995), considerando que su hidrografía está poco desarrollada (Tabla 2). En contraste con lo anterior, SCL se encuentra bien descripta por los datos (Fig. 3a). Las curvas de riqueza de Sobs, Cole, Jack1 y ICE son coincidentes y se aproximan a una meseta en los últimos sitios de muestreo. Además, la curva de ICE tiene la forma esperada para regiones que poseen un número de muestreos suficientes y confiables (Heyer et al. 1999). SCL posee una gran intensidad de muestreos (por sitio) y en diferentes estaciones del año (Fernández et al. 2002) con lo cual se consiguió una gran validez en los datos. En relación con lo anterior, podemos asegurar que existe un buen conocimiento de la fauna bentónica de artrópodos (y probablemente también de macroinvertebrados).
En SCMa (Fig. 3b) las curvas de los estimadores describieron un comportamiento menos claro que el exhibido por SCL y SCC (Heyer et al. 1999). La distancia entre Sobs y los demás estimadores resultó grande en comparación con la subcuenca bien muestreada (Fig. 3a), mientras que las tendencias de las curvas de los estimadores considerados fueron ascendentes. Esto supone la necesidad de incrementar el número (otros sitios) y la intensidad (distintas épocas) de los muestreos. Del mismo modo, se considera que la similitud existente en la riqueza estimada por Jack1 para ocho sitios de muestreo entre SCC y SCL variará cuando se incremente la información biológica de la primera. Esta suposición se basa en diferencias significativas encontradas al comparar la taxocenosis de hidrácaros usando datos de abundancia de ambas subcuencas (Fernández 2003). Además existen otras diferencias importantes entre éstas como ser orientación, disposición, características físicas locales, entre otros factores que deberán ser tenidos en cuenta.
En el ACC, SCM estuvo caracterizada por el hidrofílido monotípico Hydramara argentina. Ésta especie es típica de ríos de montaña del NOA y habita en zonas con restos de vegetación de deriva (Archangelsky 1997). Por otra parte, SCMa se caracteriza por los taxones Heterelmis sp., Guajirolus sp., Triplectides sp. y Helichus sp. Esta subcuenca, a pesar de no estar bien muestreada, posee un conjunto de taxones propios atribuibles a ciertas particularidades de la unidad (presencia de un embalse, monto pluviométrico, etc). La influencia de SCL y el conjunto de taxones propios (extremo inferior del eje 2, Fig. 4) que constituyen, posiblemente, el grupo de especies raras predichas en la distribución log-normal (Magurran 1989), resultaron destacables.
Es importante destacar que otras variables no consideradas por lo fragmentarias en su disponibilidad para el ACC, podrían ajustar mejor como explicación de la distribución del bentos. Así, consideramos que estudios posteriores deberán incluir otras variables no medidas aún en las subcuencas, como ser, proporción de los distintos estratos de vegetación, relación vegetación natural-cultivos y orientación de las mismas, entre otras.
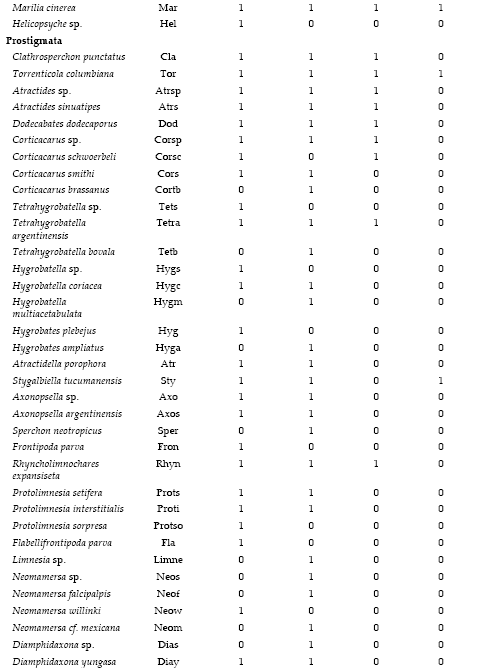
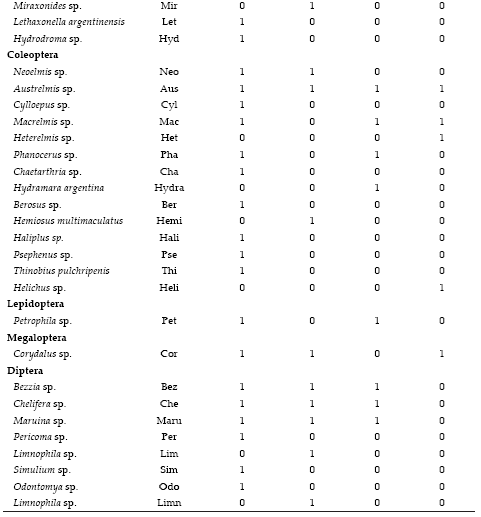
Apéndice 1. Composición taxonómica y distribución del bentos en cuatro subcuencas del noroeste argentino. Los códigos corresponden al ACC.
Appendix 1. Taxonomic composition and distribution of benthic fauna in four sub-basins of northwestern Argentina. The codes are used in CCA.

AGRADECIMIENTOS
Al Dr. E. Dominguez por su colaboración incondicional para realizar este trabajo. Al Ing. R. A. Fernández por su ayuda con la cartografía. Al equipo de trabajo por el apoyo en tareas de campo, identificaciones y valiosos comentarios. A la Agencia de Promoción Científica y Tecnológica, Pict: 01-12529 y al CONICET al cual pertenecen los autores. A las Universidades Nacionales de Tucumán y del Litoral.
BIBLIOGRAFÍA
ALLAN, DA & AS FLECKER. 1993. Biodiversity conservation in running waters. BioScience, 43:32-43. [ Links ]
ARCHANGELSKY, M. 1997. Studies on the biology, ecology and systematics of the inmature stages of New World Hydrophiloidea (Coleoptera: Staphyliniformia). Bull. Ohio Biol. Surv., 12(1):58-59. [ Links ]
TER BRAAK, CJF. 1995. Ordination. Pp. 91-173 in: RHG Jongman; CJF ter Braak; OFR Van Tongeren (eds). Data analysis in community and landscape ecology. Cambridge University Press. USA. [ Links ]
COLWELL, R. 1997. EstimateS 5.0.1. Statistical estimation of species richness and shared species from samples. Department of Ecology and Evolutionary Biology, University of Connecticut. USA. [ Links ]
DOMINGUEZ, E & HR FERNÁNDEZ. 1998. Calidad de los ríos de la cuenca Salí (Tucumán, Argentina) medida por un índice biótico. Serie conservación de la naturaleza Nº 12. Fundación Miguel Lillo, Tucumán, Argentina. [ Links ]
DOMINGUEZ, E & JM BALLESTEROS VALDEZ. 1992. Altitudinal replacement of Ephemeroptera in subtropical river. Hidrobiología, 246:83-88. [ Links ]
FERNÁNDEZ, HR & E DOMINGUEZ. 2001. Guía para la determinación de artrópodos bentónicos sudamericanos. Editorial Universitaria de Tucumán. Universidad Nacional de Tucumán. Argentina. [ Links ]
FERNÁNDEZ, HR; F ROMERO; M PERALTA & L GROSSO. 2001. La diversidad del zoobentos en ríos de montaña del noroeste de Argentina: comparación entre seis ríos. Ecol. Austral, 11:9-16. [ Links ]
FERNÁNDEZ, HR; F ROMERO; MB VECE; V MANZO; C NIETO & M ORCE. 2002. Evaluación de tres índices bióticos en un río subtropical de montaña (Tucumán-Argentina). Limnetica, 21(1-2):1-13. [ Links ]
FERNÁNDEZ, HR. 2003. Structure of water mite taxocoenoses in two northwestern Argentinean subtropical sub-catchments. Systematic & Applied Acarology, 8:55-66. [ Links ]
GRAU, A & A BROWN. 2000. Development threats to biodiversity and opportunities for conservation in the mountain ranges of the upper Bermejo River basin, NW Argentina and SW Bolivia. Ambio, 29:445-450. [ Links ]
HEYER, WR; J CODDINGTON; WJ KRESS; P ACEVEDO; D COLE ET AL. 1999. Amazonian biotic data and conservation decisions. Cienc. Cult. (Sao Paulo), 51(5/6):372-385. [ Links ]
MAGURRAN, AE. 1989. Diversidad ecológica y su medición. Ediciones Vedrá. Barcelona, España. 200 pp. [ Links ]
MELO, AS & CG FROEHLICH. 2001. Evaluation of methods for estimating macroinvertebrate species richness using individual stones in tropical streams. Freshw. Biol., 46:711-721. [ Links ]
SALA, OE; F STUART; CHAPIN III; JJ ARMESTO ET AL. 2000. Global Biodiversity scenarios for the year 2100. Science, 287:1770-1774. [ Links ]
VINSON, MR & CP HAWKINS. 1998. Biodiversity of stream insects: variation al local, basin and regional scales. Annu. Rev. Entomol., 43:271-93. [ Links ]
WARD, W & K TOCKNER. 2001. Biodiversity: towards a unifying theme or river ecology. Freshw. Biol., 46:807-819. [ Links ]
WU, J & W GAO. 1995. Spatial patterns of species richness: a hierarchical perspective. Chinese biodiversity, 3:12-21. [ Links ]


{kind=link}