










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.17 n.2 Córdoba jul./dic. 2007
TRABAJOS ORIGINALES
Disturbios en una estepa arbustiva del Monte: cambios en la vegetación
Alicia I Kröpfl1, *, Víctor A Deregibus2 & Gustavo A Cecchi3
1. CURZA, Universidad Nacional del Comahue, Viedma, Río Negro, Argentina.
2. Cátedra de Forrajicultura, Universidad de Buenos Aires, Buenos Aires, Argentina.
3. Est. Exp. Agropecuaria Valle Inferior, Viedma, Río Negro, Argentina.
* CURZA, Universidad Nacional del Comahue, Mons. Esandi y Ayacucho, (8500) Viedma, Río Negro, Argentina. Email: akropfl@uncoma.edu.ar
Recibido: 12 de abril de 2007;
Fin de arbitraje: 24 de abril de 2007;
Revisión recibida: 29 de junio de 2007;
Aceptado: 31 de julio de 2007
RESUMEN. En una estepa arbustiva del Monte oriental rionegrino se evaluó el efecto producido por la acción individual y combinada del fuego, desmonte mecánico y pastoreo doméstico sobre la vegetación que sustenta la producción ganadera de la región. Se instalaron clausuras en sitios afectados por grandes disturbios (fuego, desmonte, y ambos), y se tomaron mediciones fuera y dentro de ellas con el fin de evaluar estos disturbios. Se estudiaron diversas características estructurales (densidad y cobertura basal de los arbustos, biomasa aérea y cobertura basal del estrato herbáceo y de la costra biológica) y una funcional (fenología de las cinco gramíneas y los dos arbustos dominantes). El fuego y el desmonte redujeron la densidad y la superficie ocupada por los arbustos, y también la cobertura de costra biológica. A su vez, estos efectos se profundizaron cuando ambos disturbios actuaron en conjunto. El fuego modificó cualitativamente la composición florística del estrato herbáceo al alterar la dominancia de Stipa tenuis Phil. a favor de una especie más palatable como es Stipa papposa Nees. El pastoreo redujo de manera homogénea la biomasa aérea del estrato herbáceo, pero prolongó el período vegetativo de las gramíneas y aumentó el tamaño de ese compartimiento en desmedro del reproductivo, lo cual resultó en una mejor calidad de forraje disponible para los herbívoros. La ausencia general de interacciones entre el fuego o el desmonte con el pastoreo permitiría predecir la dirección de cambio de este sistema frente a cada disturbio en particular.
Palabras clave: Fuego; Desmonte; Pastoreo; Interacciones; Costra biológica.
ABSTRACT. Disturbances on a shrubby steppe of the Monte phytogeographical province: changes in vegetation: We evaluated the effects of individual and joint effects of fire, mechanical removal of shrubs and grazing on natural vegetation of the eastern Monte Phytogeographical Province in Rio Negro, Argentina. Four enclosures were constructed in sites where large disturbances (fire, mechanical removal and both) had taken place. Measurements were made both inside and outside them in order to evaluate not only the grazing effects in each site, but also possible interactions between the latter and large disturbances, which have not usually been reported. Several structural characteristics (shrub density and cover, aerial biomass and basal cover of herbaceous layer, and of biological crust), plus a functional one (phenology of five grasses and two shrubs) were studied. Both the fire and the mechanical removal reduced the density and area covered by the shrubby layer, and also the biological crust cover, with the effects being larger at the site were both had taken place. Bare soil was higher in the disturbed sites respect to the control due to the disappearance of the biological crust. Fire produced a qualitative change in the floristic composition of the herbaceous layer through altering the dominance from Stipa tenuis Phil. to a more palatable species as it is Stipa papposa Nees. Grazing reduced homogeneously aboveground biomass of the herbaceous layer, but extended the vegetative period of grasses and increased the size of that budget though reducing the reproductive one. The last two effects will be beneficial for secondary production since they increase the energy flux to herbivory. Large disturbances, however, could have negative effects through the reduction of the biological crust, which plays an important role in seed germination, seedlings establishment and water balance, thus contributing to ecosystems sustainability.
Keywords: Fire; Mechanical removal; Grazing; Interactions; Biological crust.
INTRODUCCIÓN
Una gran cantidad de ecosistemas áridos y semiáridos del mundo ha experimentado incrementos en la cobertura de leñosas y reducciones en la cobertura de graminoides en el último siglo, y este proceso no parece fácil de revertir (Berlow et al. 2003). Se han utilizado diversas formas de control de arbustos (desmontes, aplicación de herbicidas, quemas controladas) con el propósito de restablecer la vegetación herbácea y reducir la competencia, pero los resultados han sido variables (Morton & Melgoza 1991; Giorgetti et al. 1997).
La cobertura arbustiva, que sería determinante de muchas de las propiedades del sistema, estaría regulada tanto por la disponibilidad de recursos como por los regímenes de disturbios (Sankarán et al. 2005). Oesterheld et al. (1999) citan tres tipos de disturbios como modeladores de la estructura y el funcionamiento de pastizales y sabanas: la herbivoría, el fuego, y las variables climáticas. Si bien el aumento de la cobertura de leñosas en un sistema podría estar asociado a un calentamiento climático, la fuerza conductora parece ser los altos niveles de herbivoría de los animales domésticos (Van Auken 2000). La herbivoría ha reducido la biomasa aérea de los pastos perennes, su cobertura (Bisigato & Bertiller 1997) y su vigor, llevando a una disminución del combustible fino y la consecuente reducción en la frecuencia (o aún la eliminación) de los incendios en los pastizales y estepas arbustivas, donde su ocurrencia habría contribuido a controlar la densidad de plantas leñosas e impedir que invadan nuevas áreas (Walker & Noy Meir 1982). El pastoreo de áreas con un componente arbustivo normalmente lleva a un incremento en la dominancia de los arbustos y la conversión de los pastizales en arbustales menos deseables, sobre los cuales, además, habría un mayor consumo por unidad de producción que en los pastizales (Milchunas & Lauenroth 1993).
Sankaran et al. (2005) sostienen que, en sistemas con precipitaciones inferiores a los 650 mm, la cobertura de leñosas sólo está restringida por la falta de humedad y no por el fuego, los herbívoros o el tipo de suelo. Sin embargo, según la hipótesis de los "dos estratos" de Walter, en estos sistemas limitados por agua los arbustos compiten con los pastos por el agua de los estratos superficiales del suelo pero sólo los arbustos tienen acceso al agua almacenada en los estratos profundos (Soriano & Sala 1983; Knoop & Walker 1985; Sala et al.1989; Kröpfl et al. 2002). El régimen de precipitaciones de nuestro sistema, con las mayores lluvias concentradas en otoño y primavera y sin eventos torrenciales, en años normales permite la recarga profunda del suelo, básicamente a través del flujo por tronco, favoreciendo a los arbustos en detrimento de los pastos (Cecchi et al. 2006).
El fuego puede alterar la composición de una comunidad según la resistencia relativa de cada especie al fuego y su habilidad para rebrotar (Boó et al. 1997): las plantas resistentes aumentarían su vigor y tasa de crecimiento y reducirían su producción de semillas (Whelan 1995) o presentarían dormancia en sus semillas, lo cual explicaría en parte su persistencia en estos sistemas perturbados (Mayor et al. 2003). Oesterheld et al. (1999) señalan la falta de datos de incendios para pastizales correspondientes a precipitaciones inferiores a los 400 mm, si bien suponen que los efectos del fuego en estas regiones pueden ser severos. El daño parece estar inversamente relacionado con la frecuencia con que ocurren los incendios, pero al aumentar la severidad del fuego los mayores efectos se producen sobre el estrato arbustivo (Bran et al. 2007). En un estudio realizado en la región austral del Monte, Rostagno et al. (2006) hallaron un cambio drástico en la fisonomía de la vegetación producido por el fuego, pasando de una estepa arbustiva a una estepa graminosa. En tales sistemas, con una larga historia de pastoreo ovino, la sola remoción del ganado no cambió la dominancia de los arbustos, aunque favoreció la recuperación de los pastos que proporcionaron suficiente biomasa para que el fuego se propagara. Si bien los fuegos y las condiciones ambientales variaban entre sitios, tanto la biomasa como la cobertura y densidad de los pastos perennes de las áreas quemadas aumentaron. Los autores, sin embargo, mencionan lo difícil que resulta predecir los cambios en la vegetación cuando se combinan el pastoreo y la variabilidad de las lluvias después del fuego, y existen datos de Giorgetti et al. (1997) que muestran que las respuestas particularmente dependen de las precipitaciones anuales.
Todos los disturbios mencionados tienen lugar en el tipo fisonómico-florístico más extendido (unos 4000 km2) del Monte Oriental, en la provincia de Río Negro (Bran et al. 2005); la ganadería es la principal actividad productiva de la región, los incendios ocurren de manera natural y los desmontes se realizan como una práctica de control del monte. La vegetación es una estepa arbustiva baja dominada por Chuquiraga erinacea D.Don subs. erinacea y Condalia microphylla Cav. (en algunos sectores acompañados por Prosopis alpataco Phil.), y el estrato inferior es un pastizal predominantemente invernal dominado por Stipa tenuis Phil. Un tercer grupo funcional de interés, aunque muchas veces inadvertido, son las costras biológicas que cubren el suelo; en este sistema, sus principales componentes son dos especies de Briófitas: Syntrichia princeps (De Not.) Mitt y Ceratodon purpureus (Hedw.) Brid.
Las costras biológicas tendrían un papel fundamental en la germinación y establecimiento de nuevas plántulas, la infiltración de las precipitaciones, y la reducción de los efectos de los encostramientos superficiales (St. Clair et al. 1984). Numerosos trabajos muestran el efecto negativo del pastoreo (Belnap et al. 2001; Scutari et al. 2004) sobre las costras biológicas (especialmente a través del pisoteo), y algunos describen el efecto del fuego o la maquinaria sobre ellas (Belnap et al. 2001; Hawkes & Fletchner 2002), pero no es fácil encontrar trabajos que reporten lo que ocurre cuando el pastoreo se superpone a los otros disturbios.
Nuestro trabajo sustenta la idea de que los disturbios que actúan sobre este ecosistema a distintas escalas pueden modelar la vegetación a través de la modificación de la cobertura, densidad, biomasa y fenología de las especies, aún cuando la variabilidad climática podría ser mucho más determinante, como sugieren varios de los autores citados. Dado que las interacciones entre disturbios pueden ser un aspecto potencialmente importante y no tan conocido como el de algunos procesos aislados, el objetivo de nuestro trabajo fue evaluar los efectos del fuego, el desmonte mecánico, el pastoreo y la combinación de algunos de esos disturbios sobre la vegetación natural, a través de algunas características estructurales de los estratos arbustivo, herbáceo y costra biológica, y el comportamiento fenológico de algunas especies de los primeros dos estratos en el Monte oriental rionegrino. Nuestras predicciones fueron: (1) si existen interacciones, los efectos de los disturbios no serían simplemente aditivos, y en cada caso la respuesta de la vegetación podría tomar distintas direcciones; (2) si los disturbios producen cambios en la estructura del estrato arbustivo, la fisonomía de la comunidad se vería alterada, por períodos variables según las posibilidades del sistema de recuperarse; (3) si los disturbios producen estrés en las plantas, esto se verá reflejado en cambios en la fenología de las especies, especialmente en la floración, dependiendo de su historia evolutiva y grado de adaptación a cada disturbio.
METODOLOGÍA
El estudio se realizó en un campo de Monte ubicado a unos 30 km de la localidad de Viedma, en el departamento Adolfo Alsina de la provincia de Río Negro (40º48'S; 63º14'W). El relieve del área es plano y extendido, y los suelos son arcillosos, moderadamente alcalinos y con salinidad en profundidad; el clima es subtemplado seco de transición, con veranos cálidos e inviernos moderados, sin ningún exceso estacional de agua, y la temperatura media anual es de 15ºC. Las precipitaciones promedian los 362 mm. anuales y presentan una alta variabilidad, tanto intra como interanual; las mayores precipitaciones se producen en otoño y primavera, aunque llueve con mayor frecuencia en invierno, en forma de lluvias pequeñas. El clima es ventoso, especialmente en primavera y verano, y ello contribuye a hacer más marcado el déficit de humedad del suelo.
El trabajo se llevó a cabo durante 1985 en un potrero de 400 ha representativo de la vegetación de la región, en el cual tuvieron lugar todos los disturbios que se querían evaluar: se había desmontado en franjas en 1982 con una rastra pesada, se había quemado accidentalmente (en sentido transversal al desmonte) en el verano de 1984 y, tras haber permanecido en descanso durante ese período, fue pastoreado en forma continua por bovinos (con una carga de 6 ha/U.G., muy superior a la media de la zona) durante el desarrollo del trabajo. Se instalaron cuatro clausuras fijas de aproximadamente 0.25 ha cada una, para excluir el pastoreo de grandes herbívoros, en sitios que habían sido afectados por cada uno de los distintos disturbios, y se realizaron mediciones dentro y fuera de las clausuras, para evaluar también el efecto del pastoreo en cada sitio (n = 8).
Las mediciones se iniciaron al año de instalarse las clausuras, y las características analizadas fueron: densidad y superficie ocupada por arbustos, cobertura basal y biomasa aérea de las especies herbáceas, y fenología de las dos especies arbustivas y las cinco especies de gramíneas dominantes. La densidad (total y específica) de los arbustos se estimó mediante el "método de los cuartilos" (Bonham 1989), tomando 30 estaciones muestrales en cada uno de los sitios, y para cada arbusto, paralelamente, se estimó el área cubierta (como proyección de la copa) a través de las medidas de sus dos diámetros transversales. Con ambas medidas pudo estimarse la proporción de la superficie accesible al pastoreo en cada sitio. La cobertura basal de especies herbáceas, suelo desnudo y costras biológicas se estimó mediante líneas de Canfield (Bonham 1989) en áreas libres de arbustos, en cuatro transecciones de 5 metros de largo para cada sitio y fecha, discriminándose la especie en el caso de las gramíneas. Debido a que las áreas del potrero donde podían hallarse "puras" cada una de las distintas situaciones eran limitadas y no se pudieron realizar varias clausuras por disturbio, en este caso se considera que en realidad se trabajó con pseudoreplicaciones para corregir ese error (Hurlbert 1984), y los datos se analizaron de manera de detectar diferencias entre disturbios y no entre localizaciones. La biomasa aérea del estrato herbáceo se estimó mediante un corte al ras en el momento de mayor acumulación de biomasa (fin de primavera), en parcelas muestrales cuyo tamaño y número se determinaron previamente para lograr una distribución normal de los datos (Rice 1967). Se ubicaron al azar 15 parcelas de 40 cm. de lado entre los arbustos, y el material cosechado se separó manualmente en los compartimentos "broza", "seco en pie" y "vivo", distinguiéndose en el caso de las gramíneas la especie y el estado de desarrollo (vegetativo o reproductivo). Finalmente, se determinó el peso de la materia seca, y los resultados se expresaron en kg/superficie efectivamente accesible al pastoreo por hectárea. Se registró la fenología de cinco especies de gramíneas (Stipa tenuis, S. papposa, S. clarazii Ball., Poa ligularis Nees.ap.Steud, Pappophorum caespitosum R.E.Fr.) y los dos arbustos dominantes en esa comunidad (Condalia microphylla y Chuquiraga erinacea), en forma mensual o bimensual durante un año. Para ello se marcaron aleatoriamente cuatro plantas de cada especie por sitio, y en cada una se registró periódicamente qué proporción de fases "vegetativa", "reproductiva" y "muerta o inactiva" presentaban. En el caso de las gramíneas, al finalizar la observación se extrajeron otros individuos de las mismas especies cuyas proporciones se estimaron en forma análoga para realizar posteriormente un recuento de los macollos que se hallaban en cada fase y analizar mediante un test de Bondad de Ajuste si existían diferencias entre la estimación visual y el recuento. Las diferencias nunca fueron significativas. En el caso de los arbustos, se consideró "inactiva" toda rama que mostrara caída de hojas. Los datos obtenidos fueron transformados (arcsenvp), y se analizó si existían diferencias en los porcentajes de fases vegetativas, reproductivas y muertas entre los distintos sitios y fechas.
Los datos fueron analizados mediante ANA-VAs, tests de Tuckey para Comparación de medias en los casos en los que se detectaron diferencias (Sokal & Rohlf 1981) o ANOM (análisis de medias-proporciones binomiales) para los análisis de proporciones.
RESULTADOS
Estrato arbustivo
El fuego (F) y el desmonte (D), separados o en forma conjunta (DF), redujeron significativamente (P < 0.05) la densidad y la superficie ocupada por los arbustos como también la riqueza florística de ese estrato (Tabla 1). No se detectaron diferencias debidas a las clausuras ni a la interacción, y por ello los datos de los dos sitios (clausura y pastoreo, C y P) se reunieron de manera de contar con mayor número de muestras por tratamiento. Los disturbios de mayor magnitud (F, D y DF) redujeron significativamente (P < 0.01) la densidad y la cobertura de Chuquiraga erinacea, el arbusto dominante en todos los sitios (Tabla 1). Ningún disturbio afectó significativamente (P > 0.05) la densidad de Condalia microphylla, (Tabla 1), y si bien su cobertura mostró menores valores en los sitios perturbados, las diferencias no fueron estadísticamente significativas. En base a los valores medidos de densidad y cobertura específicos, se calculó la superficie ocupada por cada especie en la hectárea y se estimó la proporción de la superficie accesible al pastoreo en cada sitio (Tabla 1). Si bien C. erinacea fue la especie más abundante, no se descontó su cobertura en ese cálculo debido a que por su baja altura media (± 1.10 m) y su forma de cono invertido no generaría un obstáculo importante para el pastoreo bajo su copa.
Tabla 1. Densidad y cobertura de los dos arbustos dominantes en la vegetación, y total del estrato (± SD), riqueza florística, y proporción de la superficie cubierta por arbustos en un sitio testigo (T) y tres sitios perturbados por Fuego (F), Desmonte (D) y Desmonte-Fuego (DF). Las letras minúsculas indican diferencias estadísticamente significativas entre tratamientos (P < 0.05).
Table 1. Density and cover of the two dominant shrubs on vegetation, and total of the layer (± SD), floristic richness and proportion of surface covered by shrubs in a control site (T), and three sites disturbed by fire (F), mechanical removal (D) and mechanical removal-fire (DF). Small letters show statistical significant differences between treatments (P < 0.05).

No hubo diferencias fenológicas significativas entre los sitios C y P durante el período del ensayo para ninguna de las especies. En todos los tratamientos tanto C. microphylla como C. erinacea conservaron una alta proporción de ramas en estado vegetativo (Fig. 1) a lo largo del año y mantuvieron sus hojas durante el período de reposo. El área sólo desmontada (D) presentó mayor proporción de material muerto o inactivo que las áreas quemadas (F y DF), y la caída de las hojas viejas se registró en primavera, recién después de producida la extensión de los nuevos tallos a fines del invierno. Los sitios quemados presentaron la menor proporción de material reproductivo, y en el caso de Condalia microphylla no se registró floración en absoluto en el tratamiento FP. En todos los sitios se registró una alta variabilidad en las proporciones de floración entre individuos.

Figura 1. Proporciones de fenofases vegetativas, reproductivas e inactivas de Chuquiraga erinacea y Condalia microphylla en un sitio testigo y tres sitios perturbados por Fuego, Desmonte, y Desmonte-y-Fuego, bajo condiciones de: a) Clausura y b) Pastoreo (separadas por el área en blanco) a lo largo del año.
Figure 1. Proportions of vegetative, reproductive and inactive phenophases of Chuquiraga erinacea and Condalia microphylla in three sites disturbed by fire, mechanical removal and mechanical removal-and-fire, under a) exclosure and b) grazing conditions along the year.
Estrato herbáceo
El Pastoreo redujo significativamente (P < 0.05) la biomasa verde total, y homogeneizó las respuestas respecto a los otros disturbios, sin que se registraran interacciones significativas. Los otros disturbios, en cambio, redujeron la biomasa seca total (Fig. 2), aunque la reducción sólo fue significativa en el caso de D (P < 0.05), y no se registraron efectos debidos a P ni a la interacción. Los sitios quemados arrojaron la mayor (P < 0.05) biomasa vegetativa de Stipa papposa (Fig. 2), aunque no mostraron diferencias significativas en los valores de biomasa reproductiva (probablemente debido a la alta variabilidad de los datos), de manera que la biomasa total reflejó las diferencias halladas en la vegetativa, y no se registraron efectos debidos a la interacción.
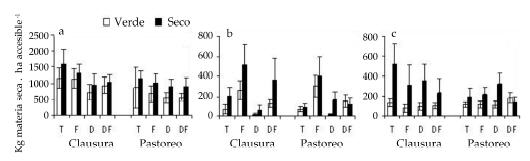
Figura 2. Biomasa aérea verde y seca (+ EE) total (a), Stipa papposa (b) y S. tenuis (c) en un sitio testigo (T) y tres sitios disturbados por Fuego (F), Desmonte (D) y Desmonte-Fuego (DF) bajo condiciones de clausura (C) y pastoreo (P). (Observar que los gráficos a) y c) no tienen la misma escala que el a)).
Figure 2. Green and dry aerial biomass (+ SE) for total (a), Stipa papposa (b) and S. tenuis (c), in a control site (T) and three sites disturbed by fire (F), mechanical removal (D) and mechanical removal-fire (DF), under exclosure (C) and grazing (P) conditions. (Charts b) and c) are at different scales than a)).
Los sitios quemados (F y DF) también mostraron la mayor (P < 0.05) cobertura basal de esta especie (Tabla 2). Los disturbios no afectaron significativamente (P > 0.05) la biomasa total de Stipa tenuis, y esta alcanzó su mayor valor en TC (Fig. 2) debido a la mayor cantidad de material reproductivo en ese tratamiento, aunque sólo difirió en forma significativa (P < 0.05) del tratamiento más disturbado (DFP). La interacción fue significativa (P < 0.05) en el caso de la biomasa vegetativa, y en el tratamiento más disturbado (DF), el sitio P superó al C, a la inversa de lo ocurrido en el tratamiento T; en entre sitios (Fig. 3). Los disturbios tampoco afectaron significativamente (P > 0.05) la cobertura basal de esta especie (Tabla 2).
Tabla 2. Cobertura basal (%) de diversos componentes del estrato herbáceo, suelo desnudo y costra biológica en el sitio testigo (T) y los 3 sitios disturbados (F, D y DF). Las letras minúsculas indican diferencias significativas entre sitios (P < 0.05).
Table 2. Basal cover (%) of several components of the herbaceous layer, bare soil and biological crust in the control site (T), and three disturbed sites (F, D and DF). Small letters show statistical significant differences between sites (P < 0.05).


Figura 3. Biomasa aérea vegetativa de Stipa tenuis por superficie accesible en el sitio T y los sitios F, D y DF bajo condiciones de Clausura (C) y Pastoreo (P). Las letras minúsculas indican diferencias significativas entre sitios para cada tratamiento (P < 0.05).
Figure 3. Aerial vegetative biomass of Stipa tenuis in site T and sites F, D and DF under exclosure (C) and grazing (P) conditions. Small letters show statistical significant differences between sites for each treatment (P < 0.05).
El comportamiento fenológico en respuesta a los disturbios fue variable para las distintas gramíneas estudiadas, si bien el ciclo general de las cuatro gramíneas invernales fue similar, comenzando la floración a fines del invierno y reiniciando el crecimiento vegetativo al promediar el verano. El Pastoreo fue el disturbio que mayor efecto tuvo sobre Stipa tenuis, pues redujo marcadamente la floración de las plantas y aumentó porcentualmente el compartimiento vegetativo. En todos los sitios, los macollos reproductivos murieron en diciembre-enero y contribuyeron a aumentar la proporción del compartimiento "muerto", que incluyó cañas secas en pie del año anterior (Fig. 4). El Pastoreo prácticamente no modificó las proporciones de los distintos tipos de males descriptas precedentemente. El Pastoreo en el sitio T redujo la floración de Pappophorum , la única especie con síndrome fotosintético C, pero no hubo otros efectos observables. En todos los sitios esta especie collos de Stipa papposa (Fig. 4), y en general los resultados muestran que esta especie no fue afectada negativamente por ningún disturbio. Al clasificar las fenofases, se consideró que a partir de diciembre los macollos reproductivos volvían a ser "vegetativos" debido a la característica de esta especie de conservar verdes las cañas al perder las espigas. Una observación que cabe destacar es que encontramos plantas que presentaban cortos rizomas, si bien esta característica no aparecía descripta en la bibliografía. El Pastoreo fue el disturbio que más modificó la fenología de Poa ligularis al reducir su compartimiento reproductivo, aunque en este caso no de manera tan notoria como en las Stipas, pues esta especie siempre presenta una baja proporción de macollos florales. En el caso de S. clarazii, la falta de disturbios (sitio T) redujo la floración, aunque en todos los tratamientos de manera similar; la aparición de los macollos florales comenzó un par de semanas más tarde que en las tres gramíneas invernales descriptas precedentemente. El Pastoreo en el sitio T redujo la floración de Pappophorum caespitosum, la única especie con síndrome fotosintético C4, pero no hubo otros efectos observables. En todos los sitios esta especie detuvo su actividad con la primera helada y no la reanudó hasta la primavera, manteniéndose hasta marzo. Los primeros macollos vegetativos registrados en primavera fueron macollos anillados que parecían helados (por su color morado) y reverdecieron desde la base.

Figura 4. Proporciones de fenofases vegetativas, reproductivas y muertas de Stipa tenuis y S. papposa en un sitio testigo y tres sitios disturbados por Fuego, Desmonte, y Desmonte-y-Fuego, bajo condiciones de a) clausura y b) pastoreo, a lo largo del año.
Figure 4. Proportions of vegetative, reproductive and death phenophases of Stipa tenuis and S. papposa in three sites disturbed by fire, mechanical removal and mechanical removal-and-fire, under a) exclosure and b) grazing conditions.
Ninguno de los disturbios afectó significativamente (P > 0.05) la cobertura basal de las Dicotiledóneas herbáceas (Tabla 2). Los tratamientos F, D y DF incrementaron el suelo desnudo y redujeron la superficie cubierta por costra biológica (P < 0.05) (Tabla 2), sin que se hayan registrado diferencias debidas a P ni a la interacción, por lo cual los valores de los sitios C y P aparecen promediados.
DISCUSIÓN
Los disturbios de mayor magnitud y menor frecuencia (fuego y desmonte) impactaron sobre el estrato arbustivo y la costra biológica, mientras que el pastoreo afectó a las gramíneas forrajeras que forman parte del estrato herbáceo, homogeneizando sus respuestas frente a los otros disturbios. La ausencia general de interacciones entre los disturbios permitiría predecir la dirección particular de los cambios en el sistema frente a la ocurrencia de cada uno de ellos.
El mayor daño directo producido por el fuego a los arbustos respecto a los pastos, podría explicarse tanto por la diferente ubicación de los puntos de crecimiento de cada grupo funcional como por sus diferencias en el desarrollo fenológico. Los incendios de la región tienen lugar básicamente en verano, momento en que los arbustos se encuentran en activo crecimiento y los pastos en reposo, con la mayor parte de su biomasa seca. Tales incendios dañarían severamente a los arbustos debido a la localización más expuesta de sus meristemas respecto a los de los pastos, la pérdida del material fotosintetizante necesario para aportar energía a los órganos reservantes en el otoño (Richburg et al. 2002), y la nueva inversión que deben realizar en tejidos de sostén (Heisler et al. 2004). También la reducción (o aún la ausencia) de floración encontrada en los arbustos que rebrotaron después del fuego y del desmonte podría entenderse como parte de una estrategia de persistencia en el sistema, que prioriza la asignación de los recursos producidos durante los primeros años al crecimiento vegetativo en desmedro del reproductivo (Schwilk & Ackerly 2005). En cambio, el fuego estival, beneficiaría a los pastos directamente al remover el material senescente acumulado en la estación de crecimiento previa sin afectar los meristemas, e indirectamente, como sugieren Heisler et al. (2004), a través de la mejora de su ambiente energético (disponibilidad de luz y temperatura del suelo).
Por otro lado, el más rápido aprovechamiento inicial por parte de los pastos de los recursos liberados después de un incendio podría conferirles ventaja frente a los arbustos para rebrotar cuando las condiciones climáticas son favorables, pues el fuego, además de reducir la competencia de los arbustos, libera espacios previamente ocupados por ellos (Cook et al. 1994). En parcelas donde se había realizado remoción de arbustos, Inouye (2006) encontró un aumento en el contenido de humedad en el suelo respecto a sitios intactos, mostrando un balance de agua más favorable para los pastos. Berlow et al. (2003) mostraron un aulos pastos que el daño directo que les produjo mento en la productividad y la cobertura de las especies herbáceas al remover los arbustos en una estepa arbustiva de Artemisia rothrockii, y el incremento fue mayor cuando las parcelas se regaron, sugiriendo que la remoción de arbustos en sí misma es capaz de incrementar la biomasa de herbáceas en sitios áridos durante los años húmedos. Coincidiendo con esos resultados, Peláez et al. (2001) sólo detectaron mortalidad de pastos en el Caldenal cuando en el período posterior a la quema se produjo una sequía, en tanto el fuego estival no mató las plantas ni cambió su diámetro basal, por lo cual concluyeron que la disponibilidad hídrica fue más importante para el rebrote de el disturbio.
En nuestro estudio, el fuego mostró efectos positivos o neutros sobre el estrato herbáceo, pero no todas las formas de remoción de arbustos actuarían en la misma dirección, como lo sugieren las diferencias fenológicas registradas entre los sitios perturbados. En el sitio desmontado todas las especies mostraron mayor proporción de floración, y si la tasa reproductiva de las especies aumenta en condiciones de stress hídrico, como proponen Walker & Noy Meir (1982), nuestros datos estarían señalando algún grado de aridez relativamente mayor en ese sitio que en los otros. Ese déficit hídrico sería consecuencia de la reducción de la porosidad superficial del suelo y de la generación de microdepresiones de infiltración reducida por compactación del suelo que produjo el tránsito de la maquinaria utilizada en este caso (Valentin 1981; Fuls 1992).
El fuego determinó un importante cambio en la composición florística, ya que la dominante del estrato herbáceo pasó a ser Stipa papposa en lugar de S. tenuis. Su ventaja para colonizar sitios quemados probablemente derive del hecho de que también tendría yemas de renuevo subterráneas que asegurarían la supervivencia de la especie y le conferirían mayor habilidad inicial para ocupar espacios abiertos después del incendio. Si bien ambas especies pueden clasificarse como "deseables" (Giorgetti et al. 1997), Stipa papposa es una especie altamente preferida por los animales y posee la característica de mantener verdes sus cañas florales en el verano después de caídas las panojas, lo cual la convierte en una de las pocas forrajeras disponibles para el herbívoro en el período de menor oferta de biomasa verde del pastizal. Probablemente la presencia de rizomas detectada también contribuya a su persistencia en el sistema frente a los disturbios.
Las otras dos gramíneas invernales, Poa ligularis y Stipa clarazii, también conservaron una importante proporción de material vegetativo durante el verano y esto las haría tan deseables en el pastizal como S. papposa, pero tanto ellas como la estival Pappophorum caespitosum son especies de baja constancia en este pastizal, baja cobertura basal y escaso aporte de biomasa, y su abundancia no fue incrementada por efecto de los disturbios.
Stipa tenuis no modificó su cobertura por efecto de los disturbios e incluso aumentó su biomasa y crecimiento vegetativo, coincidiendo con los resultados hallados para esta especie después de fuegos (Boó et al. 1996; Peláez et al. 2003) o defoliaciones (Becker et al. 1997 a, b y c) en el Caldenal y que los autores atribuyen a su tolerancia al fuego y a su crecimiento compensatorio frente al pastoreo por producción de nuevos macollos o aún de raíces. Aún en estos sistemas tan dependientes del clima, el crecimiento compensatorio explicaría parte de los efectos positivos del pastoreo sobre la productividad (Oesterheld et al. 1999). La menor biomasa vegetativa de esta especie encontrada en el sitio pastoreado del testigo respecto a la clausura podría deberse a una mayor presión de pastoreo sobre esta especie en ese sitio respecto a los otros, debido a que el estrato herbáceo resulta menos accesible por la presencia de los arbustos.
El aumento en la proporción de macollos vegetativos de las gramíneas forrajeras producido por el pastoreo implicaría un menor costo energético para las plantas y, desde el punto de vista de la utilización del pastizal, representaría una mejora para su aprovechamiento.
La ausencia de daño sobre la costra biológica por acción del pastoreo contradice la información corriente en la bibliografía que muestra como el pisoteo de los animales afecta negativamente tales costras en diversos ecosistemas áridos y semiáridos (Memmott et al. 1998), incluyendo el Monte del NE de Chubut (Scutari et al. 2004), donde evaluaron la abundancia de un grupo morfológico de líquenes (caracterizado por su alta sensibilidad a la degradación) frente al pastoreo. La falta de respuesta al pastoreo observada en nuestro caso podría estar vinculada, por un lado, al gran daño que le produjeron el fuego y el desmonte previos, pero también al hecho de que nuestra medición se realizó a fines de primavera antes del comienzo de la sequía estival. En nuestra región, el verano sería el momento de mayor susceptibilidad a la degradación, pues al secarse las costras pueden desprenderse fácilmente del suelo por efecto del pisoteo, resultar fragmentadas y desaparecer (Belnap et al. 2001), lo cual en estos sistemas conlleva riesgos para la sustentabilidad.
AGRADECIMIENTOS
Agradecemos a la Profesora C. Matteri del Museo Argentino de Ciencias Naturales "Bernardino Rivadavia" por determinarnos las especies de musgos. Los comentarios del Dr. M. Oyarzábal, un revisor anónimo, el Dr. R. Distel, y la editora de la revista, Dra. M. Semmartin, contribuyeron a mejorar este manuscrito con valiosas sugerencias y críticas. Este trabajo fue realizado con un subsidio de CONICET.
BIBLIOGRAFÍA
BECKER, GF; CA BUSSO & T MONTANI. 1997a. Effects of defoliating Stipa tenuis and Piptochaetium napostaense at different phonological stages: axillary bud viability and growth. J. Arid Environ., 35:269-283. [ Links ]
BECKER, GF; CA BUSSO; T MONTANI; MA BURGOS; AC FLEMMER & MB TORIBIO. 1997b. Effects of defoliating Stipa tenuis and Piptochaetium napostaense at different phonological stages: root growth. J.Arid Environ., 36:639-653. [ Links ]
BECKER, GF; CA BUSSO; T MONTANI; AL ORCHANSKY; RE BREVEDAN; MA BURGOS ET AL. 1997c. Effects of defoliating Stipa tenuis and Piptochaetium napostaense at different phonological stages: tiller demography and growth. J. Arid Environ., 35:251-268. [ Links ]
BELNAP, J; R ROSENTRETER; S LEONARD; JH KALTNECKER; J WILLIAMS & D ELDRIDGE. 2001. Biological soil crusts: ecology and management. Tech. Ref. 1730-2, US Department of Interior Bureau of Land Management. Denver, Co. [ Links ]
BERLOW, EL; CM D'ANTONIO & H SWARTZ. 2003. Response of herbs to shrub removal across natural and experimental variation in soil moisture. Ecol. Appl., 13:1375–1387. [ Links ]
BISIGATO, AJ & MB BERTILLER.1997. Grazing effects on patchy dryland vegetation in northern Patagonia. J. Arid Environ., 36:639-653. [ Links ]
BONHAM, CD. 1989. Measurements for terrestrial vegetation. John Wiley & Sons (eds). New York, USA. 338 pp. [ Links ]
BOÓ, RM; DV PELÁEZ; SC BUNTING; OR ELÍA & MD MAYOR. 1996. Effect of fire on grasses in central semi-arid Argentina. J. Arid Environ., 32:259-269. [ Links ]
BOÓ, RM; DV PELÁEZ; SC BUNTING; OR ELÍA & MD MAYOR. 1997. Effect of fire on woody species in central semi-arid Argentina. J. Arid Environ., 35:87-94. [ Links ]
BRAN, D; J AYESA; G CECCHI; A MARCOLIN; C LÓPEZ & H MORAGA. 1988. Principales tipos de campos y forrajeras naturales del Noreste de la provincia de Río Negro. Pp. 176-180 en: Estación Experimental Agropecuaria Valle Inferior Convenio provincia de Río Negro. Manual de especies silvestres del Monte rionegrino. INTA. [ Links ]
BRAN, DE; GA CECCHI; JJ GAITÁN; JA AYESA & CR LOPEZ. 2007. Efecto de la severidad de quemado sobre la regeneración de la vegetación en el Monte Austral. Ecol. Austral, 17:123-131. [ Links ]
CECCHI, GA; AI KRÖPFL; NM VILLASUSO & RA DISTEL. 2006. Stemflow and soil water redistribution in intact and disturbed plants of Larrea divaricata in Southern Argentina. Arid Land Res. Manag., 20:209-217. [ Links ]
COOK, JG; TJ HERSHEY & LL IRWIN. 1994. Vegetative response to burning on Wyoming mountain-shrub big game ranges. J.Range Manage., 47:296-302. [ Links ]
FULS, ER. 1992. Ecosystem modification created by patch-overgrazing in semi-arid grassland. J. Arid Environ., 23:59-69. [ Links ]
GIORGETTI, HD; OA MONTENEGRO; GD RODRÍGUEZ; CA BUSSO; T MONTANI; MA BURGOS ET AL. 1997. The comparative influence of past management and rainfall on range herbaceous standing crop in east-central Argentina: 14 years of observations. J. Arid Environ., 36: 623-637. [ Links ]
HAWKES, CV & VR FLETCHER. 2002. Biological soil crusts in a xeric Florida shrubland: composition, abundance and spatial heterogeneity of crusts with different disturbance histories. Microb. Ecol., 43:1-12. [ Links ]
HEISLER, JL; JM BRIGGS; AK KNAPP; JM BLAIR & A SEERY. 2004. Direct and indirect effects of fire on shrub density and aboveground productivity in a messic grassland. Ecology, 85:2245-2257. [ Links ]
HURLBERT, SH. 1984. Pseudoreplication and the design of ecological experiments. Ecol. Monog., 54:187-211. [ Links ]
INOUYE, RS. 2006. Effects of shrub removal and nitrogen addition on soil moisture in sagebrush steppe. J.Arid Environ., 65:604-618. [ Links ]
KNOOP, WT & BH WALKER. 1985. Interactions of woody and herbaceous vegetation in a southern African savanna. J. Ecol.,73:235-253. [ Links ]
KRÖPFL, AI; GA CECCHI; NM VILLASUSO & RA DISTEL. 2002. The influence of Larrea divaricata on soil moisture and on water status and growth of Stipa tenuis in southern Argentina. J. Arid Environ., 52:29-35. [ Links ]
MAYOR, MD; RM BOÓ; DV PELÁEZ & IR ELÍA. 2003. Seasonal variation of the soil seed bank of grasses in central Argentina as related to grazing and shrub cover. J. Arid Environ., 54:467-477. [ Links ]
MEMMOTT, KL; VJ ANDERSON & SB MONSEN. 1998. Seasonal grazing impact on Cryptogamic crusts in a cold desert ecosystem. J. Range Manage., 51:547-550. [ Links ]
MILCHUNAS, DG & WK LAUENROTH. 1993. Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol. Monogr., 63:327-366. [ Links ]
MORTON, HL & A MELGOZA. 1991. Vegetation changes following brush control in creosotebush communities. J. Range Manage., 44:133-139. [ Links ]
OESTERHELD, M; J LORETI; M SEMMARTIN & JM PARUELO. 1999. Grazing, fire and climate effects on primary productivity of grasslands and savannas. Pp. 287-306 in: L Walker (ed). Ecosystems of disturbed ground. Elsevier. Amsterdam, Netherlands. [ Links ]
PELÁEZ, DV; RM BOÓ; OR ELIA & MD MAYOR. 2003. Effect of fire on growth of three perennial grasses from central semi-arid Argentina. J. Arid Environ., 55:657-673. [ Links ]
PELÁEZ, DV; RM BOÓ; MD MAYOR & OR ELÍA. 2001. Effect of fire on perennial grasses in central semiarid Argentina. J. Range Manage., 54:617-621. [ Links ]
RICE, EL. 1967. A statistical method for determining quadrat size and adequacy of sampling. Ecology, 48:1047-1049. [ Links ]
RICHBURG, JA; AC DIBBLE & WA PATTERSON III. 2002. Woody invasive species and their role in altering fire regimes of the Northeast and Mid-Atlantic states. Pp. 104-111 in: KEM Galley & TP Wilson (eds.). Proceedings of the Invasive Species Workshop: the Role of Fire in the Control and Spread of Invasive Species. Fire Conference 2000: the First National Congress on Fire Ecology, Prevention and Management. Misc. Publ. No. 11. Tall Timbers Research Station. Tallahassee, FL, USA. [ Links ]
ROSTAGNO, CM; GE DEFOSSE & HF del VALLE. 2006. Postfire vegetation dynamics in three rangelands of Northeastern Patagonia, Argentina. Rangeland Ecol. Manage., 59:163-170. [ Links ]
SALA, OE; RA GOLLUSCIO; WK LAUENROTH & A SORIANO. 1989. Resource partitioning between shrubs and grasses in the Patagonian steppe. Oecologia, 81:501-505. [ Links ]
SANKARAN, M; NP HANAN; RJ SCHOLES; J RATNAM; DJ AUGUSTINE; BS CADE; ET AL. 2005. Determinants of woody cover in African savannas. Nature, 438:846-849. [ Links ]
SCHWILK, DW & DD ACKERLY. 2005. Is there a cost to resprouting? Seedling growth rate and drought tolerance in sprouting and nonsprouting Ceanothus (Rhamnaceae). Am. J. Bot., 92: 404-410. [ Links ]
SCUTARI, NC; MB BERTILLER & AL CARRERA. 2004. Soil-associated lichens in rangelands of north-eastern Patagonia. Lichen goups and species with potential as bioindicators of grazing disturbance. The Lichenologist, 36:404-412. [ Links ]
SOKAL, RR & FJ ROHLF. 1981. Biometry. The principles and the practice of statistics in biological research. 3a edn. WH Freeman & Co (eds). New York, USA. 858 pp. [ Links ]
SORIANO, A & O SALA. 1983. Ecological strategies in a Patagonian arid steppe. Vegetatio, 56:9-15. [ Links ]
ST CLAIR, LL; BL WEBB; JR JOHANSEN & GT NEBEKER. 1984. Cryptogamic soil crusts: enhancement of seedling establishment in disturbed and undisturbed areas. Recl. and Reveg. Res., 3:129-136. [ Links ]
VALENTIN, C. 1981. Organisations pellicullaires superficielles de quelques sols de regions subdesertique (Agadez-Republique du Niger). Dynamique de formation et consequences sur l'economie en eau. These. Université de Paris VII, U.E.R. de Sciences physiques de la terre, Laboratorie de Pedologie. [ Links ]
VAN AUKEN, OW. 2000. Shrub invasions of north american semiarid grasslands. Annu. Rev. Ecol. Syst., 31:197-215. [ Links ]
WALKER, BH & I NOY MEIR. 1982. Aspects of the stability and resilience of savanna ecosystems. Pp. 577-590 in: BH Huntley & BH Walker (eds). Ecology of tropical savannas. Springer-Verlag. New York, U.S.A. [ Links ]
WHELAN, RJ. 1995. The ecology of fire. Cambridge Univ. Press. Cambridge, UK. [ Links ]

