










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.18 n.1 Córdoba ene./abr. 2008
TRABAJOS ORIGINALES
Composición y estructura de la vegetación epífita vascular en un bosque primario de Olivillo (Aextoxicon punctatum R. et P.) en el sur de Chile
José San Martín1, Alejandro Espinosa2, *, Silvana Zanetti2, Enrique Hauenstein3, Nelson Ojeda2 & César Arriagada2
1. Instituto de Biología Vegetal y Biotecnología, Universidad de Talca, Talca, Chile.
2. Departamento de Ciencias Forestales, Universidad de La Frontera, Temuco, Chile.
3. Facultad de Recursos Naturales, Universidad Católica de Temuco, Temuco, Chile.
* Departamento de Ciencias Forestales, Universidad de La Frontera, Casilla 54-D, Temuco, Chile. Email: aes@ufro.cl
Recibido: 25 de abril de 2006;
Fin de arbitraje: 10 de abril de 2007;
Revisión recibida: 25 de septiembre de 2007;
Aceptado: 15 de octubre de 2007
RESUMEN. La flora epífita vascular de los bosques templados y su relación con el hábitat no ha sido estudiada en profundidad. En este trabajo analizamos estos componentes en un bosque de la asociación Lapagerio-Aextoxiconetum, en un cordón montañoso de la Depresión Intermedia de la Región de La Araucanía, Chile. Se identificaron ocho especies de epífitas vasculares, distribuidas en cuatro géneros y tres familias: Asplenium dareoides (Aspleniaceae), Hymenoglossum cruentum, Hymenophyllum caudiculatum, H. cuneatum, H. pectinatum, H. plicatum e H. secundum (Hymenophyllaceae) y Sarmienta scandens (Gesneriaceae). Estas especies constituyen el 72.7% de la flora epifítica del bosque, de la cual un 62.5% se encuentra en la categoría vulnerable. Se determinó que la especie más importante es Hymenophyllum cuneatum, que también es la más higrófila. En el otro extremo, Hymenophyllum plicatum fue la más resistente a la sequía. Finalmente, debido a que la presencia de esta diversa comunidad de epífitas se desarrolla sobre diferentes forófitos presentes en el bosque primario de la asociación Lapagerio-Aextoxiconetum (Aextoxicon punctatum, Persea lingue, Nothofagus obliqua, Eucryphia cordifolia y Laureliopsis philippiana), se concluye que la vegetación epifítica vascular es altamente dependiente de la permanencia del bosque.
Palabras clave: Forófito; Hospedero; Hymenophyllaceae; Comensalismo.
ABSTRACT. Composition and structure of vascular epiphytes in an old-growth forest of Olivillo (Aextoxicon punctatum R. et P.) in southern Chile: Vascular epiphytes represent 10% of the total vascular plants of the world. Nevertheless, because it is hard to reach them in the forest upper-canopy where they usually live, there are few studies (especially in Chile) about them. With the objective of identifying the vascular epiphytes growing on tree stems (under 1.5 m height), we sampled three transects in an Olivillo old-growth stand, in the Rucamanque forest, in the central valley of south-central Chile, near the city of Temuco. Several variables were measured for both the vascular epiphytes and their host-trees. We studied the floristic composition of the vascular epiphytes community, and their relationship with their host-trees, as well as their sociability, cover, and frequency. Eight vascular epiphytes species were identified, corresponding to four genus distributed in three families: Hymenophyllum and Hymenoglossum (Hymenophyllaceae), Asplenium (Aspleniaceae) and Sarmienta (Gesneriaceae). We found only a non-statistically significant linear correlation between vascular epiphytes and their host-trees. We determined that Hymenophyllum cuneatum was the most important vascular epiphytes species.
Keywords: Phorophytes; Host; Hymenophyllaceae; Commensalism.
INTRODUCCIÓN
En los ecosistemas forestales es posible identificar cinco estratos: arbóreo, arbustivo, herbáceo, lianas o trepadoras y plantas epífitas (Kimmins 1996), lo que refleja la complejidad de la estructura vertical de los bosques, dependiente de las formas de crecimiento y de las estrategias de las especies para aprovechar la luz y la humedad del sitio. En este contexto, el estrato de menor representatividad florística son los epífitos vasculares, alcanzando sólo un 10% de la flora mundial hasta hoy descrita (Benzing 1989; Kress 1989). El epifitismo es una interacción comensalística entre dos especies (Smith & Smith 2001), en la cual una es el forófito u hospedero y la otra una planta epífita. En el caso del bosque, el hospedero es un árbol vivo, el cual sin exponerse a un daño fisiológico, beneficia con el soporte a otra especie epífita de menor crecimiento. La planta epífita, al fijarse en la corteza del fuste, logra una posición más elevada, con mejor alcance a la luminosidad y a la precipitación fustal (Donoso 1981; Granados & Tapia 1990, Nieder et al. 1997).
La riqueza y el tipo de especies epífitas de un bosque se relaciona, fundamentalmente, con factores ambientales, tales como las precipitaciones y la temperatura, que regulan su desarrollo y continuidad temporal (Hietz 1998). Otros factores importantes son la presencia e interacción con otras especies vegetales y las características del sustrato (Hietz 1998; Hernández-Rosas 2001). La mayor representatividad del grupo se observa en los bosques húmedos tropicales y templados húmedos (Webb 1959). Este hecho posiciona a los epífitos como una comunidad (Braun-Blanquet 1950), indicadora de la calidad ecológica y del estado de conservación e integridad física del bosque (Hietz 1998; Barthlott et al. 2001). En la actualidad, la fragmentación de los bosques atenta contra la estabilidad de los ecosistemas forestales. Las consecuencias de este proceso se traducen en pérdida de hábitats, disminución de la biodiversidad, así como también en la modificación de los procesos biológicos de las especies que componen la comunidad (Riveros & Ramírez 1978; Gullison & Nissan 1999).
Los bosques dominados por Aextoxicon punctatum R. et P. se han descrito como los más propicios para el establecimiento y desarrollo de epífitas vasculares de Chile (Riveros & Ramírez 1978; Godoy et al. 1981). Sin embargo, los fragmentos de este bosque ubicados en la depresión intermedia de la Región de La Araucanía en Chile, constituyen un caso poco documentado (Oberdorfer 1960; Ramírez & Figueroa 1985). El objetivo del presente trabajo consistió en caracterizar la composición y estructura de la vegetación epífita vascular asociada al bosque de A. punctatum. Partimos de la hipótesis que la diversidad de la flora epífita está regulada por el tipo de forófito y las condiciones generales del hábitat; es decir, el tipo de corteza y condiciones de humedad a que está expuesta la vegetación epífita.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se realizó en el predio Rucamanque, de 435.1 ha, propiedad de la Universidad de La Frontera, ubicado 12 km al norte de la ciudad de Temuco, Región de La Araucanía, Chile (38° 39' S, 72° 35' W; Salas & García 2006). El lugar se caracteriza por tener un clima templado-húmedo, con una precipitación media anual de 1400 mm distribuidos a lo largo del año, pero con mayor intensidad en los meses de invierno, con uno o dos meses de sequía en verano y una temperatura media anual de 12°C (Di Castri & Hajek 1976). Corresponde a una quebrada con orientación sureste a noroeste, ubicada en la ladera oriental del cordón montañoso Huimpil-Ñielol, cuyas altitudes varían entre 191 msnm y 556 msnm (Ramírez et al. 1989). En los sectores más bajos predominan los suelos rojo arcillosos y en sectores más altos, depósitos de trumao (CIREN-CORFO 1989).
Rucamanque es actualmente considerado Sitio Prioritario de Conservación (CONAMA 2002), debido a su vegetación relicta y rica en endemismos, característica de la depresión intermedia del centro sur de Chile (Arroyo et al. 1996); además, en el interior del bosque es posible encontrar varias especies con problemas de conservación, entre las que se destacan Satureja multiflora (R. et P.) Briq. y Citronella mucronata (R. et P.) D. Don., entre las especies de flora (CONAF 1987); Dromiciops gliroides Thomas, Felis concolor L. y Pudu pudu Mol., entre las especies de fauna (González et al. 2000; Quintana et al. 2000).
En el sitio de estudio se encuentran distintas formaciones de vegetación, siendo el tipo boscoso la de mayor importancia ecológica. En su interior se encuentra representada la asociación Lapagerio Aextoxiconetum Oberdorfer, dominada por Aextoxicon punctatum R. et P., que es la que ocupa la mayor superficie (250.6 ha) (Ramírez et al. 1989a; 1989b). La estructura vertical de esta comunidad forestal es compleja ya que posee múltiples estratos: se pueden identificar todos los estratos descritos por Kimmins (1996). Las especies arbóreas perennifolias más importantes son Aextoxicon punctatum R. et P., Persea lingue (R. et P.) Ness, Weinmannia trichosperma Cav. y Laureliopsis philippiana (Phil) Losser, en tanto, entre las caducifolias se destaca Nothofagus obliqua (Mirb.) Oerst, que es una especie colonizadora.
Diseño del muestreo
En el interior del bosque de A. punctatum se muestrearon individuos arbóreos con las siguientes características: tener vegetación epifítica vascular sobre sus fustes, pertenecer a las clases de iluminación de copas emergente y plena iluminación superior (Dawkins 1958; citado por Synnott 1991); tener la corteza intacta y no tener oquedades en los primeros 2 m del fuste, correspondiente a la Zona I de Johansson (Gradstein et al. 2003). Para evaluar la vegetación epífita, primero se determinó el tamaño adecuado de las parcelas de muestreo, a través de la confección de la curva especies-área (Braun-Blanquet 1950; Mueller-Dombois & Ellemberg 1974; Steubing et al. 2001), muestreando los fustes de cinco árboles elegidos al azar que cumplieran las características antes mencionadas. Luego se trazaron tres transectos de 250 m cada uno, paralelos entre sí y perpendiculares a las curvas de nivel aplicando el tamaño de parcelas determinado en la etapa anterior. Cada transecto se ubicó al azar, se eligió un individuo arbóreo cada 25 m, muestreándose un total de 10 árboles por transecto, sobre los cuales se ubicaron dos parcelas de 0.20 cm2 (45 cm x 45 cm), en las caras norte y sur de cada fuste, respectivamente. Para cada individuo arbóreo se consignó: especie, diámetro a la altura del pecho (DAP), altura total, clase de iluminación de copa, tipo de corteza (lisa, rugosa, agrietada, escala de Gibson 1996) y la posición topográfica en la ladera (ladera alta, media o baja). A la vegetación epifítica vascular se le determinaron los siguientes atributos de composición y estructura: especies presentes, sociabilidad (formaciones puras, pequeñas colonias, en matas, en grupos o aisladas), densidad y cobertura (proyección sobre el fuste, %) por especie (Braun-Blanquet 1950; Margalef 1998; Hernández-Rosas 2001). Para cada especie se calculó la frecuencia relativa de distintas clases de cobertura y sociabilidad, de densidad y de posición en el fuste. A nivel de comunidad, la vegetación epífita vascular fue analizada considerando el número total de especies, géneros y familias. Se obtuvo la diversidad alfa, que depende de las especies presente en un mismo hábitat (Whittaker 1967), se calculó la frecuencia y densidad relativa de cada especie (Margalef 1998) y el índice de importancia como la suma de ambas características, indicador de la importancia ecológica de cada especie en el ecosistema (Hernández-Rosas 2001). La diversidad beta, medida del grado de partición del ambiente en parches o mosaicos que permite evaluar la contigüidad de hábitats diferentes en el espacio, se calculó mediante el índice de Jaccard (Moreno 2001). Para ello las epífitas vasculares se agruparon según especie de forófito, tipo de corteza, cara del fuste y gradiente de humedad según posición topográfica del sector. A partir de esta información se generaron dendrogramas, mediante un programa estadístico (SPSS: Statistical Package for the Social Sciences ®). Para determinar la preferencia de las especies epífitas vasculares por algún forófito o variable del hábitat en particular, se realizó un análisis de Pearson, considerando una correlación positiva entre un par de variables si el valor de significancia del índice era menor a 0.05 (Ludwing & Reynolds 1988).
RESULTADOS
Se identificaron cinco especies de forófitos: Aextoxicon punctatum (Aextoxicaceae), Nothofagus obliqua (Fagaceae), Persea lingue (Lauraceae), Eucryphia cordifolia (Eucryphiaceae) y Laureliopsis philippiana (Monimiaceae). A. punctatum resultó ser la especie más frecuente (66%) y la que registró los diámetros máximos y mínimos (104 y 26 cm, respectivamente). El diámetro medio de los forófitos fue de 61 cm y su altura media 28 m. En la clase plena iluminación superior de la copa se agruparon cuatro de las cinco especies de forófitos (73.3%), mientras que la clase emergente (26.7%) agrupó casi exclusivamente los individuos de N. obliqua. La corteza más común es la lisa (70.0%), donde el 90.5% corresponde a individuos de A. punctatum; en cambio, los tipos rugosa y agrietada, sólo están representados en un 13.3% y un 16.6%, respectivamente, repartidos en individuos de las cinco especies presentes.
Se identificaron ocho especies de epífitas vasculares, una de las cuales, Sarmienta scandens (J.D.) Bradis, pertenece a la clase Magnoliopsida, familia Gesnerianceae, mientras que las siete restantes pertenecen a la clase Polypodiopsida. Dentro de esta clase, las especies están representadas dos familias: Aspleniaceae con Asplenium dareoides Desv. e Hymenophyllaceae, con seis especies: Hymenoglossum cruentum (Cav.) K. Presl., Hymenophyllum caudiculatum Mart. var. productum (K. Presl.) C. Chr., H. cuneatum Kunze var. cuneatum, H. pectinatum Cav., H. plicatum Kaulf. e H. secundum Hook. Grev. Las distintas especies epifíticas no mostraron preferencia por un forófito particular (Tabla 1), excepto H. cruentum que sólo se registró sobre A. punctatum. Por otro lado, resultó destacable que A. punctatum fue el único hospedero que albergó a todas las especies. Del mismo modo, N. obliqua también ofreció un hábitat adecuado para el establecimiento de las epífitas vasculares, aunque en menor grado que A. punctatum. Las epífitas H. caudiculatum e H. cuneatum fueron las únicas que superaron en muchos casos el 75% de cobertura (Figura 1), en contraste con Sarmienta scandens, que no superó el 5%. Los resultados anteriores complementan lo documentado en la Figura 2, donde nuevamente H. caudiculatum var. productum e H. cuneatum var. cuneatum fueron las únicas que en algunos casos crecieron en formaciones puras. A su vez, A. dareoides y S. scandens sólo crecieron como plantas individuales. La diferencia entre las dos últimas especies se manifestó en su estado de desarrollo, por cuanto A. dareoides se encontró siempre en estado adulto, mientras que S. scandens como plántulas de escasa edad y tamaño.
Tabla 1. Presencia y número de especies epífitas (diversidad alfa) según hospederos.
Table 1. Presence and number of epiphyte (alfa diversity) species according to host.
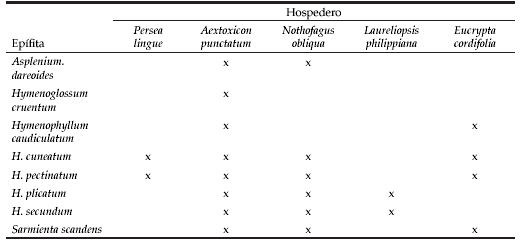
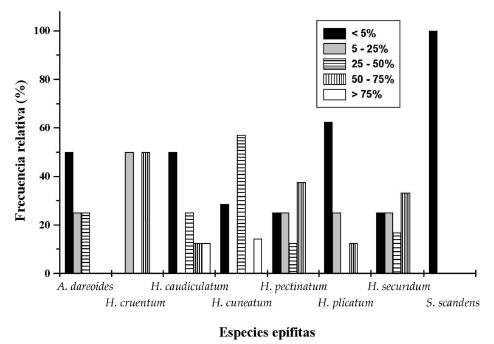
Figura 1. Distribución de ocho especies de epífitos según clases de cobertura.
Figure 1. Distribution of eight epiphytes according to cover classes.

Figura 2. Distribución de ocho especies de epífitos según su sociabilidad.
Figure 2. Sociability distribution for eight epiphytes (solitarily, in groups, cushions, in small colonies, pure population stands).
La epífita de mayor frecuencia relativa fue H. secundum (17.3%), en tanto que H. cruentum mostró el valor más bajo (3.8%) (Figura 3). Este mismo atributo varió según la cara del fuste: en la cara norte S. scandens, H. plicatum e H. pectinatum alcanzaron los valores más altos, con un 17.4% cada una, en cambio en la cara sur los registraron H. secundum e H. caudiculatum var. productum (20.7% y 17.2%, respectivamente). La densidad relativa fue máxima para H. cuneatum var. cuneatum (50%), mientras que S. scandens sólo alcanza un 0.1% (Figura 4).
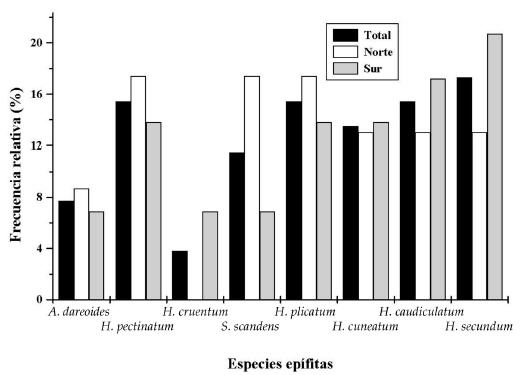
Figura 3. Distribución de los valores de frecuencia relativa por especie epífita en relación con su posición sobre el fuste según hemisferio.
Figure 3. Relative frequency for each vascular epiphyte in relation to its stem position (north or south facing).

Figura 4. Valores de la densidad relativa (%) para cada especie epifita vascular.
Figure 4. Relative density (%) for each vascular epiphyte species.
Desde el punto de vista ecológico (Tabla 2), H. cuneatum var. cuneatum resultó la epífita vascular más relevante del rodal, con un índice de importancia de 65.9 y alcanzó, además, los valores más altos en la cara sur de los fustes y en la posición topográfica de ladera media. En tanto, H. plicatum resultó más importante en la cara norte de los fustes y la posición ladera alta (39.6 y 65.8, respectivamente). Por último, en la ladera baja la especie más importante fue H. caudiculatum var. productum (90.4).
Tabla 2. Índice de importancia de las especies epífitas por hemisferio y posición topográfica.
Table 2. Importance index of epiphyte species by hemisphere (north or south facing) and topographic position (high, medium and low slope).

El mayor índice de similitud de la composición de especies entre los distintos forófitos se registró en el par formado por A. punctatum y N. obliqua, que alcanzó un índice de 0.75, mientras que L. philippiana mostró la menor similitud con el resto de los forófitos (0.15) (Figura 5). En cuanto al grado de afinidad entre diferentes posiciones topográficas, el índice de Jaccard alcanzó su máximo valor (1.0) al comparar las laderas media y baja, que presentaron una composición de especies idéntica; en tanto, la parte alta resultó levemente distinta a las anteriores, con un índice de 0.88.

Figura 5. Dendrograma de afinidad de los epífitos para los distintos forófitos, obtenido por el Índice de Jaccard. (Ap: Aextoxicon punctatum, No: Nothofagus obliqua, Pl: Persea lingue, Ec: Eucrypta cordifolia, Lp: Laureliopsis philippiana).
Figure 5. Similarity dendrogram between epiphytes and phorophytes, using Jaccard's index.
Los resultados obtenidos para los distintos tipos de corteza indicaron que existe una marcada preferencia de los epífitos por forófitos de corteza agrietada, propia de N. obliqua de gran tamaño; y por los de corteza lisa, característica de A. punctatum, reflejado en un valor del índice de Jaccard de 0.75. En cambio, el tipo de corteza rugosa sólo alcanzó un índice de similitud de 0.53, lo que indica grandes diferencias en la composición florística de la vegetación epifítica vascular que se desarrolla en los hospederos que ofrecen este tipo de sustrato.
Los valores del índice de Pearson obtenidos para las distintas situaciones indican que no existe ninguna relación entre las distintas epífitas vasculares y las condiciones de hábitat, ya sea tipo de corteza (índice= 0.06; p=0.65), posición topográfica (índice= -0.07; p= 0.62) y exposición (índice=-0.08; p= 0.59).
DISCUSIÓN
Las distintas especies de forófitos pertenecen a los estratos emergente, dominante y codominante descritos por Hueck (1978) y por Donoso (1981) para este tipo de bosque. Se trata de especies que representan la dinámica natural de estos bosques, que inicialmente están compuestos por especies intolerantes a la sombra, como Nothofagus obliqua, para luego ser reemplazadas por especies semitolerantes como Eucryphia cordifolia, que establecen las condiciones adecuadas para la colonización posterior de las tolerantes a la sombra. Estas últimas se convierten en las más abundantes, por ser las únicas capaces de regenerar en las condiciones de cobertura que se generan en el interior del bosque, como es el caso de Aextoxicon punctatum, Laureliopsis philippiana y Persea lingue (Donoso 1993).
En relación con la composición florística de las epífitas vasculares, se detectó el 72.7% del total de las especies descritas para Rucamanque por Ramírez et al. (1989a; 1989b). Dado que sólo se capturó información en los primeros dos metros del fuste, la marcada presencia de la familia Hymenophyllaceae se debería a que encuentra allí las condiciones de humedad y temperatura adecuadas para su desarrollo (Benzing 1989; Rodríguez & Baeza 1991; Marticorena & Rodríguez 1995; Clement et al. 2001). Este antecedente explica por qué especies epifíticas características de los bosques del sur de Chile, como Polypodium feuillei Bert. y Fascicularia bicolor (R. et P.) Mez. (Muñoz et al. 2003), no se registraron en el muestreo, aunque fueron vistas a mayores alturas (Zanetti, obs. pers.). Del mismo modo, los hábitats más húmedos, como las partes media y baja de la ladera y la cara sur de los fustes, resultan más adecuados para el desarrollo de las plantas de esta familia, aún cuando algunas especies muestran una alta resistencia al desecamiento (Alberdi et al. 1978; Benzing 1989), como es el caso de Hymenoglossum cuneatum var. cuneatum. El tipo de bosque es otro aspecto que favorece la presencia de helechos epífitos, ya que la asociación Lapagerio-Aextoxiconetum, por sus condiciones de luminosidad y humedad, permite el desarrollo de pteridófitos epífitos, mientras que limita la presencia de helechos hemicriptófitos (Godoy et al. 1981). La angiosperma epífita Sarmienta scandens pertenece a un género monotípico (Marticorena & Quezada 1985; Kress 1989) y es frecuente en Rucamanque, donde crece como especie trepadora (Muñoz et al. 2003). Sin embargo, en este estudio, como en otros realizados anteriormente, se ha encontrado arraigando sobre árboles, aunque a mayores alturas, debido a sus grandes requerimientos de luz (Riveros & Ramírez 1978; Hauenstein et al. 1988).
Otro resultado importante de este estudio, es que se determinó la presencia de especies con problemas de conservación, entre ellas: Asplenium dareoides, Hymenophyllum cruentum, H. caudiculatum var. productum, H. cuneatum var. cuneatum e H. secundum. Estas especies presentan un estado de conservación vulnerable (Baeza et al. 1998), hecho que aumenta considerablemente el valor ecológico del predio Rucamanque.
Los resultados de cobertura y sociabilidad son consistentes entre sí, ya que a medida que aumenta el agrupamiento también aumenta la cobertura de las distintas especies sobre los fustes (Braun-Blanquet 1950). El estado de desarrollo de las distintas especies fue un factor determinante en la cobertura de S. scandens, ya que al encontrarse sólo en estado de plántula cubría poco el fuste (Zanetti, obs. pers.). Hymenophyllum secundum presentó los mayores valores de frecuencia relativa, lo que implica que está ampliamente distribuida dentro del bosque debido, posiblemente, a su capacidad de adaptarse tanto a los ambientes húmedos como a los secos (Ramírez et al. 1976), lo cual le permite establecerse en sectores no aptos para otras especies. En el lugar de estudio es común encontrar especies de Hymenophyllaceae en los fustes de diversos árboles, así como también sobre árboles caídos y en el suelo (Ramírez et al. 1976), siendo esta última situación la más común para A. dareoides (Zanetti, obs. pers).
Se observó preferencia de las epífitas vasculares por árboles de N. obliqua y A. punctatum. N. obliqua por presentar corteza agrietada y follaje caducifolio, en relación con A. punctatum, que posee corteza lisa y follaje perennifolio. Está ampliamente documentada la preferencia de epífitas por forófitos con cortezas agrietadas, debido a que acumulan mayor cantidad de nutrientes y materia orgánica (Barthlott et al. 2001; Hernández-Rosas 2001; Nadkarni et al. 2001; Hernández-Rosas 2004). En el caso de A. punctatum, forófito con corteza lisa, la preferencia estaría más ligada con el tipo de follaje, el que al ser poco translúcido preserva mejor la humedad ambiental y regula la entrada de luz directa que puede provocar grandes daños a plantas de frondas delgadas como las de la familia Hymenophyllaceae (Benzing 1989).
Los resultados obtenidos no permiten establecer preferencias específicas de las epífitas por un determinado tipo de corteza, posición topográfica o exposición de los forófitos, resultados también evidenciados por otros autores (Hietz 1998; Barthlott et al. 2001; Nadkarni et al. 2001). Finalmente, debido a que la presencia de esta diversa comunidad de epífitas se desarrolla sobre cinco diferentes forófitos (N. obliqua, A. punctatum, P. lingue, E. cordifolia y L. philippiana) presentes en el bosque primario de Aextoxicon punctatum, se concluye que la vegetación epifítica vascular es altamente dependiente de la permanencia del bosque. Es importante continuar con esta línea de investigación en la comunidad de epifitas de los bosques templados, a fin de determinar con exactitud qué factores ambientales son los que tienen una mayor influencia en la propagación, establecimiento y desarrollo de este tipo de comunidad tan particular.
AGRADECIMIENTOS
Se agradece a la Dirección de Investigación y Desarrollo de la Universidad de La Frontera, por el financiamiento del proyecto DIUFRO N° 9802. De igual forma, se agradece a R. Carrillo, por su apoyo en la identificación de las especies vegetales y a C. Salas, por la traducción del resumen al inglés.
BIBLIOGRAFÍA
ALBERDI, M; C RAMÍREZ & L STEUBING. 1978. La Familia Hymenophyllaceae (Pteridophyta) en el Fundo San Martín, Valdivia, Chile. Vol. II Resistencia al desecamiento y sobrevivencia en comunidades antropogénicas. Medioambiente 3(2):3-13. [ Links ]
ARROYO, TK; L CAVIARES; A PEÑALOZA; M RIVEROS & AM FAGGI. 1996. Relaciones fitogeográficas y patrones regionales de riqueza de especies en la flora del bosque lluvioso templado de Sudamérica. Pp. 71-99, en: Armesto JJ, C Villagrán & MTK Arroyo (Eds.). Ecología de los bosques nativos de Chile. Editorial Universitaria. Santiago, Chile. [ Links ]
BAEZA, M; E BARRERA; J FLORES; C RAMÍREZ & R RODRÍGUEZ. 1998. Categorías de Conservación de las Pteridophytas Nativas de Chile. Boletín del Museo Nacional de Historia Natural de Chile. Dirección de Bibliotecas, Archivos y Museos (DIBAM). Santiago, Chile. 146 pp. [ Links ]
BARTHLOTT, W; V SCHMITH-NEUERBURG; J NIEDER & S ENGWALD. 2001. Diversity and abundance of vascular epiphytes: a comparision of a secondary vegetation and primary montane rainforest in the Venezuelan Andes. Netherlands. Plant Ecology 152:145-156. [ Links ]
BENZING, DH. 1989. The Evolution of Epiphytism. Pp. 15-41, en: Lüttge, U (Ed.). Vascular Plants as Epiphytes:Evolution and Ecophysiology. Ecological Studies, vol. 76. Springer-Verlag. Berlin, Heidelberg, Alemania. [ Links ]
BRAUN-BLANQUET, J. 1950. Sociología Vegetal: Estudio de las Comunidades Vegetales. Acme Agency, S. R.L. Buenos Aires, Argentina. 444 pp. [ Links ]
CIREN-CORFO. 1989. Descripciones de suelos y materiales y símbolos. Estudio agroecológico de la Provincia de Cautín, IX Región. Publicación Centro de Información de Recursos Naturales, Corporación de Fomento (CIREN-CORFO). Santiago, Chile. 210 pp. [ Links ]
CLEMENT, J; M MOFFETT; D SHAW; A LARA; D ALARCÓN & O LARRAÍN. 2001. Crown structure and biodiversity in Fitzroya cupressoides, the giant conifers of Alerce Andino National Park, Chile. Selbyana 22(1):76-88. [ Links ]
CONAMA. 2002. Sitios Prioritarios para la Conservación de la Biodiversidad en la Región de La Araucanía. Comisión Nacional del Medio Ambiente IX Región, Chile. [ Links ]
CONAF. 1987. Libro Rojo de los Vertebrados Terrestres de Chile. Glade, A (Ed.). Corporación Nacional Forestal. Santiago, Chile. 65 pp. [ Links ]
DI CASTRI, F & E HAJEK. 1976. Bioclimatología de Chile. Universidad Católica de Chile, Vice- Rectoría Académica, Santiago. 128 pp. [ Links ]
DONOSO, C. 1981. Ecología Forestal, el Bosque y su Medioambiente. Tercera Edición. Editorial Universitaria. Santiago, Chile. 369 pp. [ Links ]
DONOSO, C. 1993. Bosques templados de Chile y Argentina. Primera Edición. Editorial Universitaria. Santiago, Chile. 484 pp. [ Links ]
GIBSON, AC. 1996. Neotropical biodiversity and conservation. Occasional publication of the Mildred E. Mathias Botanical Garden. University of California, Los Angeles, USA. 202 pp. [ Links ]
GODOY, R; C RAMÍREZ; H FIGUEROA & E HAUENSTEIN. 1981. Estudios ecosociológicos en Pteridofitos de comunidades valdivianas, Chile. Bosque 4(1):12-24. [ Links ]
GONZÁLEZ, G; J TORRES-MURA & A MUÑOZ-PEDREROS. 2000. Orden Artiodactyla. En: Muñoz-Pedreros, A & J Yáñez (Eds.). Mamíferos de Chile. CEA Ediciones. Valdivia, Chile. 463 pp. [ Links ]
GRADSTEIN, R; N NADKARNI; T KRÖMER; I HOLZ & N NÖSKE. 2003. A protocol for rapid and representative sampling of vascular and non-vascular epiphyte diversity of tropical rain forest. Selbyana 24(1):105-111. [ Links ]
GRANADOS, D & R TAPIA. 1990. Comunidades Vegetales. En: Castillejos, S & J Rodríguez (Eds.). Colección Cuadernos Universitarios, serie Agronomía N° 19. Universidad Autónoma de Chapingo. México. 235 pp. [ Links ]
GULLISON, RE & S NISSAN. 1999. Evaluación de la factibilidad del modelado de los impactos del manejo forestal sobre la biodiversidad en la concesión de Tarumá. Documento Técnico 77. Proyecto Forestal Sostenible Bolivia BOLFOR. Editorial Cuarto Anillo. Santa Cruz, Bolivia. 108 pp. [ Links ]
HAUENSTEIN, E; C RAMÍREZ & M LATSAGUE. 1988. Evaluación florística y sinecológica del Monumento Natural Cerro Ñielol. Temuco. Chile. Bol. Mus. Reg. Araucanía 3:7-32. [ Links ]
HERNÁNDEZ-ROSAS, J. 2001. Ocupación de los portadores por epífitas vasculares en un bosque húmedo tropical del Alto Orinoco, Edo. Amazonas, Venezuela. Acta Científica Venezolana 52:292-303. [ Links ]
HERNÁNDEZ-ROSAS, J. 2004. Características del substrato de plantas del dosel de un bosque húmedo tropical de tierras bajas (Alto Orinoco, Venezuela). Acta Científica Venezolana 55:35-43. [ Links ]
HIETZ, P. 1998. Diversity and conservation of epiphytes in a changing environment. Invited lecture presented at the International Conference on Biodiversity and Bioresources: Conservation and Utilization, 23-27 November 1997, Phuket, Thailand. Pure Appl. Chem., 70(11). 2114 pp. [ Links ]
HUECK, K. 1978. Los bosques de Sudamérica: ecología, composición e importancia económica. Sociedad alemana de cooperación técnica Ltda. (GTZ). Alemania. 476 pp. [ Links ]
KIMMINS, JP. 1996. Forest Ecology: A Foundation for Sustainable Management. Segunda Edición. Prentice Hall Inc. New Jersey. EE.UU. 596 pp. [ Links ]
KRESS, WJ. 1989. The Sistematic Distribution of Vascular Epiphytes. Pp. 234-261, en: Lüttge, U (Ed.). Vascular Plants As Epiphytes: Evolution And Ecophysiology. Ecological Studies, Vol. 76. Springer-Verlag. Berlin Heidelberg, Alemania. [ Links ]
LUDWING, J & J REYNOLDS. 1988. Statistical Ecology. John Wiley & Son, N.York, EE.UU. 337 pp. [ Links ]
MARGALEF, R. 1998. Ecología. Novena Edición. Ediciones Omega. Barcelona, España. 951 pp. [ Links ]
MARTICORENA, C & M QUEZADA. 1985. Catálogo de la Flora Vascular de Chile. Universidad de Concepción. Concepción, Chile. Gayana Botánica 42(1-2):5-157. [ Links ]
MARTICORENA, C & R RODRÍGUEZ. 1995. Flora de Chile. Volumen I: Pteridophyta-Gymnospermae. Universidad de Concepción. Concepción, Chile. 351 pp. [ Links ]
MORENO, C. 2001. Métodos para medir la Biodiversidad. Editado por el Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo, la Oficina Regional de Ciencia y Tecnología para América Latina y El Caribe y la Sociedad Entomológica Aragonesa. Zaragoza, España. 84 pp. [ Links ]
MUELLER-DOMBOIS, D & H ELLEMBERG. 1974. Aims and Methods of Vegetation Ecology. John Wiley and Sons, Inc. New York, EE.UU. 547 pp. [ Links ]
MUÑOZ, A; P CHACÓN; F PÉREZ; E BARNERT & JJ ARMESTO. 2003. Diversity and host tree preferences of vascular epiphytes and vines in a temperate rainforest in southern Chile. Australian Journal of Botany 51:381-391. [ Links ]
NADKARNI, N; M MARK & N JURGEN. 2001. Forest Canopies, Plant diversity. Pp. 27-40, en: Levin, S (Ed.). Encyclopedia of diversity. Academic Press, San Diego, California, EE.UU. [ Links ]
NIEDER, J; PL IBISCH & W BARTHLOT. 1997. Biodiversidad de Epífitas: una cuestión de escala. La Habana, Cuba. Revista del Jardín Botánico Nacional 18:9-62. [ Links ]
OBERDORFER, E. 1960. Pflanzensociologische Studien in Chile - Ein Vergleich mit Europa. Flora et Vegetation Mundi 2:1-208. [ Links ]
QUINTANA, V; J YÁNEZ & M VALDEBENITO. 2000. Orden Carnívora. En: Muñoz-Pedreros, A & J Yáñez (Eds.). Mamíferos de Chile. CEA Ediciones. Valdivia, Chile. 463 pp. [ Links ]
RAMÍREZ, C; L STEUBING & M ALBERDI. 1976. La Familia Hymenophyllaceae (Pteridophyta) en el Fundo San Martín, Valdivia, Chile. I.- Taxonomía y Ecología. Medioambiente 2(1):21-28. [ Links ]
RAMÍREZ, C & H FIGUEROA. 1985. Delimitación ecosociológica del bosque valdiviano (Chile) mediante análisis estadísticos multivariados. Stvdia Oecologica VI:105-124. [ Links ]
RAMÍREZ, C; E HAUENSTEIN; J SAN MARTÍN & D CONTRERAS. 1989a. Study of the flora of Rucamanque, Cautín Province, Chile. Ann. Missouri Bot. Gard. 76:444-453. [ Links ]
RAMÍREZ, C; J SAN MARTÍN; E HAUENSTEIN & D CONTRERAS. 1989b. Estudio Fitosociológico de la Vegetación de Rucamanque (Cautín, Chile). Estvdia Botánica 8:91-115. [ Links ]
RIVEROS, M & C RAMÍREZ. 1978. Fitocenosis Epífitas de la Asociación Lapagerio-Aextoxiconetum en el Fundo San Martín (Valdivia-Chile). Acta Científica Venezolana 29:163-169. [ Links ]
RODRÍGUEZ, R & M BAEZA. 1991. Pteridófitos de las Áreas Silvestres Protegidas de Nahuelbuta y Contulmo, Chile. Concepción, Chile. Bol. Soc. Biol. Concepción 62:147-177. [ Links ]
SALAS, C & O GARCÍA. 2006. Modelling height development of mature Nothofagus obliqua. Forest ecology and management 229:1-6. [ Links ]
SALAS, C; A STAGE; V LEMAY; P NÚÑEZ; P PACHECO & A ESPINOSA. 2006. Spatial patterns in an old-growth Nothofagus obliqua forest in South-Central of Chile. Forest Ecology and Management 231:38-46. [ Links ]
SMITH, RL & TM SMITH. 2001. Ecología. Cuarta Edición. Pearson Education S.A. Madrid. 664 pp. [ Links ]
STEUBING, L; R GODOY & M ALBERDI. 2001. Métodos de Ecología Vegetal. Primera Edición. Editorial Universitaria. Chile. 345 pp. [ Links ]
SYNNOTT, TJ. 1991. Manual de Procedimientos de Parcelas Permanentes para el Bosque Húmedo Tropical. Valerio J (Trad.). Instituto Tecnológico de Costa Rica (ITCR), Departamento de Ingeniería Forestal. Serie de Apoyo Académico N° 12. Cartago, Costa Rica. 103 pp. [ Links ]
WEBB, J. 1959. A physiognomic classification of Australian rain forests. J. Ecol 47:551-570. [ Links ]
WHITTAKER, RH. 1967. Gradient analysis of vegetation. Biol. Rev. 42:207-264. [ Links ]

