










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.18 n.1 Córdoba ene./abr. 2008
TRABAJOS ORIGINALES
Efecto del ganado sobre los niveles de polinización en especies vegetales del monte patagónico
Mariana Tadey *
1. Laboratorio Ecotono, CRUB-Universidad Nacional del Comahue, S. C. Bariloche, Argentina.
* Laboratorio Ecotono, CRUB-Universidad Nacional del Comahue, Pasaje Gutiérrez 1125, S. C. Bariloche, Argentina. Email: mtadey@crub.uncoma.edu.arm
Recibido: 30 de julio de 2007;
Fin de arbitraje: 11 de octubre de 2007;
Revisión recibida: 20 de noviembre de 2007;
Segunda revisión recibida: 29 de noviembre de 2007;
Aceptado: 29 de diciembre de 2007
RESUMEN. El ganado puede afectar indirectamente los niveles de polinización de las plantas a través de modificaciones en el ensamble de plantas y/o sus polinizadores. Estudié el efecto del ganado sobre los niveles de polinización, estimado como número de tubos polínicos/estilo, en cinco de las especies más frecuentes del monte patagónico (Atriplex lampa, Gutierrezia solbrigii, Larrea cuneifolia, L. divaricata, Monttea aphylla). En siete potreros de similares características pero con diferentes cargas ganaderas, conté el número de tubos polínicos por estilo de flores de cada una de las especies seleccionadas, durante cuatro años consecutivos. En general, las variaciones entre años afectaron más los niveles de polinización que los cambios en la carga animal. Sólo dos de las cinco especies estudiadas presentaron cambios en sus niveles de polinización asociados a variaciones en la carga ganadera. Asimismo, el ganado no afectó de forma directa la frecuencia de visita pero afectó la cobertura de dos de las especies estudiadas. Los cambios en la cobertura (generados o no por el ganado) afectaron de forma directa e indirecta (e.g., a través de modificaciones en la frecuencia de visita) los niveles de polinización. Los resultados, tanto de los análisis de vías como de las regresiones, fueron consistentes en cuanto a las especies que fueron más afectadas por el ganado (e.g., Gutierrezia solbrigii y Monttea aphylla). Sería necesario complementar la información de la cantidad de tubos polínicos con información sobre la calidad del polen y los recursos nutritivos de las plantas para comprender mejor los efectos indirectos del ganado sobre los niveles de polinización.
Palabras clave: Carga ganadera; Efectos indirectos; Interacción planta-polinizador; Tubos polínicos.
ABSTRACT. Livestock effect on pollination levels in native plant species of the Patagonian Monte desert: Livestock may indirectly affect plant pollination levels through changes in both plants and pollinators' assemblages. I studied the effect of livestock on the pollination level, estimated as the number of pollen tubes/style, on five frequent species of the Patagonian Monte desert (Atriplex lampa, Gutierrezia solbrigii, Larrea cuneifolia, L. divaricata, Monttea aphylla). In seven paddocks that share similar habitat characteristics but with different stocking rates, I counted pollen tubes per style of each species studied during four consecutive years. In general, pollination levels are affected more by variation between years than by stocking rates. Only two of the five focal species showed changes in their pollination levels (e.g., pollen tubes/style) associated with changes in stocking rates. Likewise, livestock did not affect the frequency of pollinator visits but affected cover of two of the studied species. The changes in cover (caused or not by livestock) affected directly and indirectly (e.g., through modification in pollinator visits frequency) pollination levels. Results from the regressions and path analyses were consistent in relation to which species were more affected by livestock (e.g., Gutierrezia solbrigii y Monttea aphylla). Additional information about pollen quality and nutritional plant resources would be necessary to better understand livestock indirect effects on pollination level.
Keywords: Indirect effects; Plant-pollinators interaction; Pollen tubes; Stocking densities.
INTRODUCCIÓN
La polinización es uno de los procesos críticos para el funcionamiento de los ecosistemas y el mantenimiento de la diversidad, y es muy sensible a disturbios de origen antrópico como el sobrepastoreo por ganado introducido (Kearns & Inouye 1997; Aizen & Feinsinger 2003). Este disturbio es una de las causas más importantes del empobrecimiento de los ecosistemas naturales alrededor del mundo (Watkinson & Ormerod 2001; Cingolani et al. 2005). El ganado produce, por pisoteo o ramoneo, efectos negativos directos sobre la vegetación a través del consumo de tejido fotosintético y la eliminación de partes reproductivas de las plantas (Milchunas et al. 1988; Miller et al. 1994). Aunque también se conoce que el ganado produce efectos indirectos sobre la vegetación y/o su fauna asociada (Miyashita et al. 2004; Tadey & Farji-Brener 2007a, 2007b), sus efectos indirectos sobre la polinización están menos documentados (Vázquez & Simberlof 2003, 2004).
El ganado puede afectar indirectamente los niveles de polinización (e.g., tubos polínicos/estilo) a través de mecanismos tales como cambios en la densidad absoluta y relativa de las especies vegetales o como cambios en su estructura demográfica y/o distribución espacial de individuos sexualmente compatibles, que podrían restringir el flujo de polen (Bond 1994; Murcia 1996). Esta restricción disminuye la probabilidad que se deposite polen proveniente de plantas no emparentadas y aumenta la probabilidad de autopolinización o de polinización con polen de plantas emparentadas porque los polinizadores visitan plantas cercanas entre sí, con un menor costo de forrajeo (Harder 1988; Aizen & Vázquez 2006). Este cambio en la calidad del polen puede incrementar la proporción de tubos polínicos abortados, el aborto de semillas, la mortalidad de plántulas o disminuir el vigor de la planta adulta (Waser 1989; Barrett & Harder 1996).
Por otro lado, el ganado puede disminuir la densidad floral, ya sea porque se alimenta de flores o porque las plantas ramoneadas producen una menor cantidad. Esta disminución puede ser de tal magnitud que la comunidad vegetal no alcance a proveer suficiente alimento para mantener una adecuada diversidad y abundancia de polinizadores (Aizen & Vázquez 2006). Además, al disminuir la densidad de plantas, y en consecuencia la densidad de flores, disminuye también la atracción de los polinizadores con el consecuente cambio en sus comportamientos de forrajeo (Kwak 1987; Sih & Baltus 1987; Sowig 1989; Klinkhamer & Jong 1990; Vaughton 1992). Estos cambios en la diversidad, composición y/o abundancia del ensamble de polinizadores pueden afectar los niveles de polinización y eventualmente de producción de semillas (Aizen & Feinsinger 1994a, 1994b).
Paralelamente, el ganado puede disminuir la densidad de algunas especies de polinizadores a través de un aumento en la desecación o compactación del suelo, lo cual reduce la cantidad y calidad de los sitios de nidificación (Lovejoy et al. 1986; Saunders et al. 1991; Murcia 1995). Al mismo tiempo, otras especies de polinizadores, más oportunistas, podrían verse favorecidas por estos cambios y desplazar ciertas especies produciendo un cambio en el ensamble de polinizadores (Aizen & Feinsinger 1994a). Estos cambios en el ensamble de polinizadores también pueden afectar los niveles de polinización de las especies vegetales, ya que no todos los visitantes florales son equivalentes en su eficiencia como polinizadores (Aizen & Vázquez 2006).
En resumen, todos estos posibles efectos indirectos del ganado pueden resultar, finalmente, en una disminución en los niveles de polinización de las especies vegetales involucradas. Si esto es así, e independientemente de la importancia relativa de los diferentes mecanismos descriptos, el resultado final en la vegetación debería ser una disminución en sus niveles de polinización al incrementar la carga ganadera. Mediante este trabajo procuro explorar la existencia de dicho patrón en algunas de las especies vegetales más abundantes del monte patagónico. Si el ganado afecta negativamente los niveles de polinización (e.g., el número de tubos polínicos en el estilo), espero que la cantidad de tubos polínicos presentes en los estilos disminuya con un aumento en la carga animal. Adicionalmente, determinaré mediante modelos de ecuaciones estructurales, si esta relación entre la carga ganadera y el nivel de polinización es resultado principalmente del ensamble de polinizadores o de variaciones en la cobertura de las especies estudiadas.
MATERIALES Y MÉTODOS
Sitios de estudio
Trabajé en siete potreros localizados al NO de la Patagonia, en la provincia de Neuquén, entre Arroyito (39°5' S, 68°35' W) y Villa "El Chocón" (39°17' S, 68°55' W) (Tadey 2006). Es una región templada árida con una precipitación anual media (PMA) entre 80 y 300 mm, y lluvias distribuidas a lo largo del año (Paruelo et al. 1998; Tadey 2006). La vegetación de esta región pertenece a la provincia fitogeográfica del Monte (Cabrera 1953). Se caracteriza por ser una estepa arbustiva dominada por Larrea cuneifolia (Cavanilles) y L. divaricata (Cavanilles) asociada a varias especies arbustivas xerofíticas como Monttea aphylla (Miers) Berthan et Hooker, Bougainvillea spinosa (Cavanilles) Heimerl, Chuquiraga erinacea (Don), Prosopis alpataco (Philippi), y Atriplex lampa (Gillies ex Moquin) D. Dietrich, entre otras (Cabrera 1953). Los pastos son muy escasos (menos del 6% de la cobertura) y generalmente están representados por especies del género Stipa (Tadey 2006).
Seleccioné siete potreros que diferían en su carga ganadera pero compartían similar tipo de ambiente, clima, fisonomía vegetal y características edáficas (Del Valle 1998; León et al. 1998). Los muestreos se llevaron a cabo durante las temporadas primavera-verano del 2001 al 2004. Los potreros están situados casi consecutivamente a lo largo de la ruta nacional 237 y todos poseen la misma orientación (noroeste). La distancia máxima entre ellos fue de aproximadamente 42 km, y el área de los mismos varió entre 4000 y 25000 ha.
La carga animal y la historia de pastoreo de cada potrero (cantidad de años con presencia de ganado) se obtuvieron a partir de entrevistas a los dueños y se corroboraron con información provista por la comisión de sanidad de la provincia de Neuquén (Tabla 1). La carga ganadera se expresa como equivalente vacas por hectárea (caballo=1.25 vacas, chivas=0.17 vacas y las ovejas=0.3 vacas), según la metodología propuesta por Vallentine (2001).
Tabla 1. Carga ganadera para cada potrero expresadas en individuos/ha y en equivalente vacas/ha; especies de ganado presente en cada potrero e historia de pastoreo como cantidad de años que el potrero estuvo pastoreado por ganado hasta el momento del muestreo. Los potreros, en orden creciente de intensidad de pastoreo, son los siguientes: Mangrullo (MA), Montesino sin ganado (MOS), Campo A (A), Agua Pesada (AP), Montesino con ganado (MOC), Grossenbache (GR) y Horacio López (HL).
Table 1. Stocking rates for each paddock expressed as individuals/ha and transformed to cattle equivalent/ha, livestock type in each paddock and grazing history as number of years the paddock was grazed by cattle at the moment of sampling. Paddock names arranged according to increasing grazing intensity, are as follow: Mangrullo (MA), Montesino without livestock (MOS), Campo A (A), Agua Pesada (AP), Montesino with livestock (MOC), Grossenbache (GR) and Horacio López (HL).

Para analizar cómo la variación de carga ganadera afectaba la reproducción vegetal, seleccioné un conjunto de especies con diferentes tipos de polinización y sistemas reproductivos que estuvieran presentes en todos los sitios de estudio. Las especies elegidas fueron: Gutierrezia solbrigii, Larrea divaricata, L. cuneifolia, Monttea aphylla, Atriplex patagonica.
Niveles de polinización
Para medir los niveles de polinización recolecté, al azar, estilos de flores de las cinco especies estudiadas (Atriplex lampa, Gutierrezia solbrigii, Larrea cuneifolia, L. divaricata, Monttea aphylla). Dado que las flores de estas especies son muy pequeñas, a excepción de los dos géneros de Larrea, colecté las flores enteras y luego en el laboratorio separé los estilos. Recolecté estilos de por lo menos cinco flores por planta (10 plantas por especie, n=50) para cada especie en cada año de muestreo (2001-2004). En el 2004 recolecté, cinco estilos por planta de 20 plantas al azar para aumentar el número de réplicas por potrero. La recolección de estilos la realicé durante el pico de floración de cada especie y en flores cuya antesis había finalizado. Conservé los estilos en tubos de Eppendorf con FAA (formol:ácido acético:alcohol, 5:5:90) para su posterior tratamiento y recuento de tubos polínicos en el laboratorio. En el laboratorio realicé la diafanización de los estilos con NaOH (hidróxido de sodio, 10 N), la tinción de los mismos con anilina decolorada, y el recuento de los tubos polínicos bajo microscopio de epifluorescencia (Kearns & Inouye 1993).
Análisis estadístico
Para determinar si variaciones en los niveles de la carga ganadera afectaron la polinización de las especies estudiadas, realicé regresiones lineales entre la cantidad de tubos polínicos promedio de cada potrero (nivel de polinización; variable dependiente) y la carga de ganado (variable independiente), para cada especie y año por separado. También efectué ANOVAs de medidas repetidas para analizar la existencia de variaciones en la cantidad de tubos polínicos entre años, utilizando el promedio de tubos polínicos por potrero y año. Los datos obtenidos durante la temporada 2003 no fueron analizados, ya que en dicho año ocurrió una sequía muy intensa que afectó la floración de la zona, impidiendo que se obtuvieran un número de muestras adecuado.
La co-variación total entre dos variables (i.e., su coeficiente de correlación) puede descomponerse en componentes atribuibles a efectos directos e indirectos. Para identificar estos efectos utilicé modelos de ecuaciones estructurales (Shipley 2000), los cuales descomponen la variación encontrada en componentes causales y no causales de acuerdo a una hipótesis particular (análisis de vías). Consideré un modelo teórico hipotético para explicar cómo el ganado puede afectar indirectamente la cantidad de tubos polínicos en los estilos (niveles de polinización) a través de cambios en la cobertura vegetal y la frecuencia de visitas de los polinizadores (Fig. 1). Este modelo se planteó para cada especie por separado. La cobertura de cada especie focal fue medida sólo una vez y considerada constante durante el período de muestreo (Tadey, 2007). La frecuencia de visita de los polinizadores se midió para dos años (2003, 2004) y se utilizó el promedio entre ambos para el análisis. Asimismo, se utilizó el promedio de tubos polínicos/estilo (de todos los años muestreados) por potrero para el análisis de vías de cada especie. El objetivo de este análisis fue obtener una aproximación a los mecanismos por los cuales el ganado podría afectar indirectamente los niveles de polinización (Fig. 1). Para estos modelos el número máximo de réplicas fue de siete (n=7, i.e., número de potreros), aunque no se obtuvieron datos de todas las especies en todos los potreros, por lo cual para algunas vías el número de réplicas se redujo a cinco. La significancia de los coeficientes se obtuvo a partir de pruebas de t calculadas para estimar los parámetros de las regresiones (Legendre & Legendre 1998). Los coeficientes de cada vía representan la cantidad de cambio en una variable dependiente que está asociada a una unidad de cambio de una determinada variable independiente cuando el resto de las variables independientes permanecen constantes. Estos modelos también estiman la variación no explicada producida por factores que no fueron incluidos en el diagrama de vías (flechas verticales, Fig. 3). La estimación de los parámetros se realizó mediante el procedimiento de máxima similitud (ML, "Maximum Likelihood"), con 100 iteraciones. Se analizó la matriz de correlación directamente para obtener un modelo completamente estandarizado y poder calcular los errores estándar de cada variable. La significancia del modelo se determinó mediante la obtención de un valor crítico con distribución de X2 y su probabilidad; un X2 no significativo indica un buen ajuste del modelo a los datos (Shipley 2000). El nivel de significación de todos los análisis realizados se fijó en P<0.10, nivel comúnmente utilizado cuando el número de réplicas es bajo (Vázquez & Simberloff 2004).

Figura 1. Modelo hipotético planteado para explicar el efecto indirecto del ganado sobre los niveles de polinización de las especies estudiadas mediante un análisis de vías. Las variables estudiadas son: carga ganadera (vacas/ha), cobertura focal (promedio de la cobertura de cada una de las especies estudiadas por potrero), frecuencia de visitas (promedio por potrero de visitas de polinizadores/planta x 10 minutos de observación), tubos polínicos/estilo (promedio por potrero del número de tubos polínicos/estilo de cada especie focal, que representa los niveles de polinización de cada una).
Figure 1. Hypothetical model used to explain the indirect effect of livestock on pollination levels of the studied species using path analysis. Variables studied: stocking rate (cattle/ha), focal cover (mean cover of the focal species of each paddock), visit frequency (mean pollinators visits/plant x 10 min of observation.), and pollen tubes/style (mean pollen tubes/style per plant species and paddock).
RESULTADOS
En general, los niveles de polinización de las especies estudiadas variaron más entre años que entre potreros; casi ninguna especie mostró una dependencia de la cantidad de tubos polínicos con la carga ganadera. Sólo dos de las especies (Monttea aphylla y Gutierrezia solbrigii) mostraron una asociación con la carga animal en al menos uno de los años muestreados (Tabla 2). En el año 2002, M. aphylla aumentó sus niveles de polinización con el ganado, mientras que G. solbrigii lo disminuyó.
Tabla 2. Resumen de las regresiones lineales realizadas entre la carga ganadera (variable independiente) y el número de tubos polínicos en el estilo (variable dependiente) de cada una de las especies estudiadas para todos los años de muestreo. P significativo cuando P< 0.10.
Table 2. Linear regressions performed to determine the effect of stocking rates (independent variable) on the number of pollen tubes/style (dependent variable) for each plant species and year of sampling. P values significant at P< 0.10.
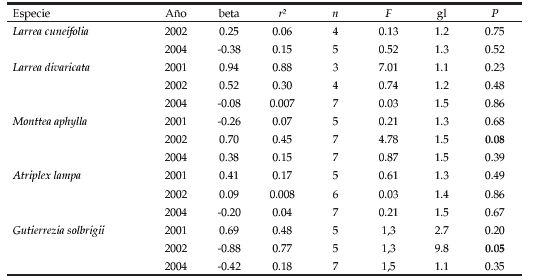
En cuanto a la variación interanual de los niveles de polinización se observó que tanto Larrea divaricata (F2,6=7.7; P=0.02), como Atriplex lampa (F2,8=44.8; P<0.001) presentaron distintos niveles de polinización entre años, siendo 2001 el año con mayores niveles de polinización (Fig. 2). Monttea aphylla también tendió a variar su nivel de polinización interanual (F2,8=3.1; P=0.10; Fig. 2). Finalmente, Larrea cuneifolia y G. solbrigii mantuvieron sus niveles de polinización aproximadamente constante a lo largo de los años muestreados (F2,2=1.25, P=0.44; F2,6= 0.3, P=0.73, respectivamente; Fig. 2).
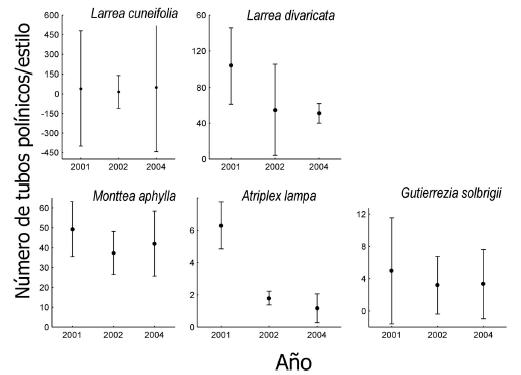
Figura 2. Promedio del número de tubos polínicos/estilo por año de muestreo para cada especie estudiada. Las barras denotan el error estándar de cada promedio.
Figure 2. Mean number of pollen tubes/style for each year and plant species. Bars represent the standard error of each mean.
El modelo puesto a prueba mediante el análisis de vías ajustó bien para todas especies estudiadas, excepto para Larrea divaricata (Fig. 3a-e). En general, la carga afectó los niveles de polinización más a través de modificaciones en la cobertura que por cambios en la frecuencia de visitas de los polinizadores. La carga tuvo un efecto negativo sobre la cobertura de las especies estudiadas, aunque este efecto fue estadísticamente significativo sólo para Gutierrezia solbrigii y Monttea aphylla (Fig. 3c y e). Sin embargo, el signo del efecto de la cobertura sobre los niveles de polinización varió según la especie; consecuentemente, el efecto neto de la carga sobre la polinización a través de la cobertura fue positivo para Monttea aphylla pero negativo para Gutierrezia solbrigii. Por otra parte, la frecuencia de visitas, aunque no fue afectada por la carga, tuvo un efecto positivo sobre los niveles de polinización en Larrea cuneifolia (Fig. 3a). Por último, en Atriplex lampa, una especie polinizada por viento, incrementos en su cobertura afectaron positivamente sus niveles de polinización (Fig. 3d).
Figura 3. Resultados de los análisis de vías representando los efectos indirectos de la carga ganadera sobre los niveles de polinización de cada especie focal a través de cambios en su cobertura y en la frecuencia de visitas de polinizadores; a) Larrea cuneifolia, b) Larrea divaricata, c) Monttea aphylla, d) Atriplex lampa, e) Gutierrezia solbrigii. Se muestra el X2 de cada análisis, grados de libertad (gl) y su probabilidad (P), resultados en negrita son estadísticamente significativos. Cada flecha posee su coeficiente de vía y su probabilidad (*P<0.10, **P<0.05, ***P<0.01) representado por el grosor de la flecha. Flechas continuas indican relaciones positivas entre variables, las discontinuas, negativas. Las variables estudiadas: carga ganadera (vacas/ha), cobertura focal (promedio de cobertura de cada una de las especies estudiadas por potrero), frecuencia de visitas (promedio por potrero de visitas de polinizadores/planta x 10 minutos de observación), y tubos polínicos/estilo (promedio por potrero del número de tubos polínicos/estilo de cada especie focal, que representa los niveles de polinización de cada una).
Figure 3. Results of the path analyses representing the indirect effect of livestock on pollination levels of the studied species through changes in their cover and frequency of pollinator visits; a) Larrea cuneifolia, b) Larrea divaricata, c) Monttea aphylla, d) Atriplex lampa, e) Gutierrezia solbrigii. Chi-square (X2), degrees of freedom and their probabilities (P) are shown for each analysis, statistically significant results are in bold letters. Each arrow has its path coefficient and probability (*P<0.10, **P<0.05, ***P<0.01), represented by arrow line-thickness. Continuous lines represent positive effects; dashed lines, negative effects. Variables studied: stocking rate (cattle/ha), focal cover (mean cover of focal species of each studied paddock), visit frequency (mean pollinators visits/plant x 10 min observation), and pollen tubes/style (mean pollen tubes/style per species and paddock).
DISCUSIÓN
El ganado, al modificar la abundancia de la vegetación y el ensamble de polinizadores, posee la capacidad de afectar indirectamente los niveles de polinización de la comunidad vegetal (Tadey 2007). Sin embargo, yo encontré un efecto de la carga animal sobre la polinización en sólo dos de las cinco especies estudiadas. En términos generales, las variaciones climáticas entre años parecen afectar más los niveles de polinización que la variación en la carga ganadera. Los niveles de polinización variaron entre años para tres de las cinco especies estudiadas (Larrea divaricata, Monttea aphylla y Atriplex lampa). En particular, el año 2001 presentó mayores niveles de polinización que el resto, posiblemente debido a mejores condiciones climáticas (Tabla 3) y/o a una mayor abundancia de polinizadores (M. Tadey, datos sin publicar), ambos factores asociadas a un incremento de la reproducción sexual en plantas (Herrera 1989; Harder & Barrett 1996; Kandori 2002; Klein et al. 2003).
Tabla 3. Datos del clima de la zona de "Villa El Chocón" proporcionados por la AIC (Autoridad Interjurisdiccional de Cuencas). Los datos representan el promedio anual de cada variable. T° min.=temperatura mínima media anual (°C); T° max.=temperatura máxima media anual (°C), Velocidad promedio del viento (km hr-1).
Table 3. Climatic data from "Villa El Chocón" obtained from the AIC ("Autoridad Interjurisdiccional de Cuencas"). The data represent the annual mean for each variable. T° min.=mean annual minimum temperature (°C); T° max.=mean annual maximum temperature (°C); Precipitation (mm); and mean wind speed (km.h-1).

La variación en los niveles de polinización fue generalmente bien explicada por el modelo hipotético propuesto, independientemente del efecto de la carga ganadera sobre las variables estudiadas. El ganado no afectó de forma directa la frecuencia de visita pero afectó la cobertura de dos de las especies estudiadas. No obstante, los cambios en la cobertura (generados o no por el ganado) afectaron de forma directa e indirecta (i.e., a través de modificaciones en la frecuencia de visita), los niveles de polinización. Los resultados tanto de los análisis de vías como de las regresiones fueron consistentes en cuanto a las especies que fueron más afectadas por el ganado. El ganado afectó negativamente la polinización de Gutierrezia solbrigii y positivamente la polinización de Monttea aphylla. Los modelos sugieren que este efecto contrastante no se debe al efecto del ganado per se (que en ambos casos afecta negativamente la cobertura de estas especies) sino al efecto de los cambios en la cobertura sobre los niveles de polinización de estas especies. Por un lado, un incremento en la cobertura afectó negativamente la polinización de M. aphylla y junto al efecto negativo del ganado sobre la cobertura resultaron en un efecto neto positivo sobre la polinización (Fig. 3c). Por el otro lado, la cobertura tuvo un efecto positivo sobre la polinización de G. solbrigii pero como el ganado afectó negativamente su cobertura, el efecto neto sobre la polinización fue negativo (Fig. 3 e). Estos resultados sugieren que el efecto del ganado sobre la polinización es generalmente a través de cambios en la cobertura, y en consecuencia, el signo de su efecto neto dependerá de la relación entre la cobertura y los niveles de polinización, el cual puede variar entre especies. Por ejemplo, si una especie posee una cantidad limitada de polinizadores, o estos son muy territoriales (e.g., como ocurre con Monttea aphylla; Tadey 2007), un incremento en la cobertura que aumente la cantidad de flores puede disminuir los niveles de polinización por un efecto de dilución. Por otra parte, si los polinizadores de una especie son abundantes y no presentan territorialidad, un incremento de cobertura que implique una mayor abundancia de flores incrementaría la atracción de polinizadores y por ende los niveles de polinización.
El efecto de un empobrecimiento en la abundancia de plantas y polinizadores sobre los niveles de polinización de una determinada especie puede depender de cómo ésta es polinizada (Aizen & Feinsinger 2003; Aizen & Vázquez 2006). En aquellas especies polinizadas por viento y dioicas, como Atriplex lampa, se espera una disminución en los niveles de polinización siempre y cuando disminuya la abundancia o cobertura de plantas, ya que éstas no necesitan de polinizadores pero sí de la presencia de individuos de su misma especie. Los niveles de polinización de esta especie no disminuyeron con la carga ganadera posiblemente porque su abundancia y cobertura no fueron afectadas considerablemente por la misma (Tadey & Farji-Brener 2007b). Sin embargo, la cobertura sí tuvo un fuerte efecto sobre los niveles de polinización.
Asimismo, aquellas especies dependientes de polinizadores y con sistemas de autoincompatibilidad, como Monttea aphylla (Tadey 2007), deberían ser afectadas seriamente por una disminución en la abundancia de plantas y/o polinizadores (Aizen & Feinsinger 2003; Aguilar 2005; Aizen & Vázquez 2006). Sin embargo, a pesar de que la abundancia de M. aphylla disminuyó con el ganado (Tadey & Farji-Brener 2007b), esta disminución incrementó sus niveles de polinización. La explicación a esta aparente contradicción, puede estar dada por el hecho de que M. aphylla depende casi exclusivamente de tres especies de polinizadores pertenecientes a la familia Anthophoridae (Tadey 2007). Dos de estas especies son especialistas en M. aphylla, mientras que la tercera es generalista (Tadey 2007). Las tres especies de polinizadores presentan un comportamiento muy territorial, visitando sólo un grupo reducido de plantas y alejando a otros polinizadores (Tadey 2007). Dado este comportamiento territorial de los polinizadores, la frecuencia de visitas podría mantenerse constante o incluso aumentar, si por disminución en la densidad de plantas, los polinizadores se ven forzados a incrementar sus visitas en el grupo de plantas remanentes para mantener los niveles de alimento necesarios. Así, la cantidad de polen transportado por los polinizadores sería potencialmente la misma aunque aumentaría la probabilidad de polinización con polen autógamo (Aizen & Feinsinger 2003).
Finalmente, para las especies parcialmente autocompatibles y dependiente de polinizadores como Larrea cuneifolia, L. divaricata y Gutierrezia solbrigii (Tadey 2007), esperaba que una disminución en la abundancia de plantas y/o polinizadores causada por el ganado disminuyera sus niveles de polinización. Sin embargo, dado que estas especies toleran cierto grado de autopolinización, este efecto podría compensarse mediante la autofecundación. Aunque una disminución en la abundancia de polinizadores podría comprometer la polinización a pesar de la autopolinización, esto es poco probable ya que estas especies son polinizadas por varias especies de polinizadores generalistas (Tadey 2007) lo que les confiere una gran tolerancia a disturbios (Jordano et al. 2003; Memmott et al. 2004; Vázquez & Aizen 2004). Dos de estas especies estudiadas (Larrea cuneifolia y Gutierrezia solbrigii) disminuyeron su abundancia y cobertura con el ganado mientras que no se observó disminución de L. divaricata (Tadey & Farji-Brener 2007b). Para Larrea cuneifolia se observó, mediante el análisis de vías, que podría existir una competencia por polinizadores ya que la cobertura afectó negativamente la frecuencia de visitas. También se observó que tanto la frecuencia de visitas como la cobertura son importantes para incrementar los niveles de polinización. En cambio, en Gutierrezia solbrigii fue más importante el aumento de la cobertura que la frecuencia de visitas para determinar la cantidad de tubos polínicos en los estilos. De cualquier manera, es probable que la calidad del polen recibido en estas especies sea menor en los potreros más pastoreados, ya que tanto la cobertura y como la abundancia de las mismas es menor (Tadey 2007). Por último, es probable que la falta de asociación observada entre la carga ganadera y la cantidad de tubos polínicos/estilo en Larrea divaricata, se deba a que no hubo cambios en su abundancia y a que es polinizada por varias especies de polinizadores generalistas (Tadey 2007).
Sin embargo, los efectos no significativos encontrados en los análisis de vías realizados pueden deberse también a otras razones, por ejemplo al bajo número de réplicas. Al utilizar réplicas reales (i.e., potreros) se evadió el peligro de la pseudo- replicación (Hurlbert 1984) pero se incrementó la probabilidad de no detectar patrones verdaderos (error de tipo II). Incrementar el número de potreros muestreados sería una solución, aunque esto no siempre es factible debido a problemas económicos y de logística que involucran la medición de varias especies simultáneamente en un área relativamente amplia. Segundo, la cantidad de tubos polínicos es normalmente muy variable, con lo cual se necesitan números de flores muy grandes para poder detectar patrones generales, y en muchos casos, es logísticamente difícil de realizar (Dafni 1992). Tercero, el efecto del ganado sobre la polinización de las distintas especies vegetales depende de la combinación entre el tipo de polinización, sus sistemas reproductivos y las especies de polinizadores que las visitan (Strauss et al. 1996; Strauss & Armbruster 1997; Morris 2003; Memmott et al. 2004). Esto hace que cada especie responda de forma muy diferente al ganado impidiendo observar un patrón general en los niveles de polinización. Cuarto, las relaciones entre variables muchas veces no son lineales y las transformaciones no son una buena solución. Quinto, los efectos negativos del ganado pueden compensarse con eventuales efectos positivos (e.g., una disminución de la competencia intra-específica por polinizadores), por lo cual pese a bajar la abundancia de una especie vegetal, los individuos remanentes no verían afectados sus niveles de polinización. Finalmente, las variaciones meteorológicas entre años pueden ayudar a compensar el efecto indirecto que el ganado pueda poseer sobre los niveles de polinización (Boyer 1988; Diggle 1992). Por ejemplo, un exceso de precipitación puede mejorar el nivel de polinización mediante una sobre-compensación por herbivoría (i.e., mayor producción de flores, mayor atracción de polinizadores) (McNaughton 1983; Maschinski & Whitham 1989; Paige 1992).
A pesar de que la cantidad de tubos polínicos está asociada a una determinada calidad de polen (ya que son los granos de polen que lograron germinar a pesar de las distintas barreras que pueden ocurrir a nivel de estigma y estilo) y que es una medición importante para los estudios de polinización, es imprescindible estudiar la calidad del polen que reciben las plantas para determinar su éxito reproductivo. Asimismo, es importante determinar cuál es el tipo de sistema de autoincompatibilidad de cada especie (si es pre- o postcigótico) y a que nivel ocurre (estigma o estilo) para comprender mejor el proceso de polinización. Todos estos datos deberían complementarse con información sobre los recursos nutritivos de las plantas y otros factores que influyan sobre la reproducción (Herrera 1987, 1989, 2000). La incorporación de estas mediciones en futuros estudios mejorará la compresión de los efectos indirectos del ganado sobre la polinización de las especies vegetales.
AGRADECIMIENTOS
Agradezco a A. G. Farji-Brener (LLM), a M. A. Aizen, M. Semmartin y dos revisores anónimos por sus comentarios en versiones anteriores que ayudaron a mejorar este trabajo. Este trabajo fue posible gracias a una beca de doctorado otorgada por CONICET.
BIBLIOGRAFÍA
AGUILAR, R. 2005. Efectos de la fragmentación de hábitat sobre el éxito reproductivo de especies nativas del bosque chaqueño serrano de Córdoba. Tesis doctorado. Universidad nacional de Córdoba, Córdoba. [ Links ]
AIZEN, MA & P FEINSINGER. 1994a. Habitat fragmentation, native insect pollinators, and feral honey bees in Argentine Chaco Serrano. Ecological Applications 4:378-392. [ Links ]
AIZEN, MA & P FEINSINGER. 1994b. Forest fragmentation, pollination, and plant reproduction in a Chaco dry forest, Argentine. Ecology 75: 330-351. [ Links ]
AIZEN, MA & P FEINSINGER. 2003. Bees not to be? Responses of insect pollinator faunas and flower pollination to habitat fragmentation. Pages 111-129 en G Bradshaw, P Marquet, y H Moonet, editores. How Landscapes Change: Human Disturbance and Ecosystems Disruptions in the Americas. Springer-Verlag Berlin Heidelberg. [ Links ]
AIZEN, MA & DP VÁZQUEZ. 2006. Flower performance in human-altered habitats.en LD Harder, & SCH Barrett, editores. Ecology and Evolution of flowers. Oxford University Press. [ Links ]
BARRETT, SCH & LD HARDER. 1996. Ecology of sexual systems of plant mating. Trends in Ecology and Evolution 11:73-79. [ Links ]
BOND, WJ. 1994. Do mutualisms matter? Assessing the impact of pollinator and disperser disruption on plant extintion. Philosophical Transactions of the Royal Society of London: Biological Sciences 344: 83-90. [ Links ]
BOYER, JS. 1988. Cell enlargement and growth-induced water potentials. Physiologia Plantarum 73:311-316. [ Links ]
CABRERA, A. 1953. Esquema fitogeográfico de la República Argentina. Revista del Museo de La Plata, Botánica 8:87-168. [ Links ]
CINGOLANI, AM; I NOY-MEIR & S DÍAZ. 2005. Grazing effects on rangeland diversity: a synthesis of contemporary models. Ecological Applications 15: 757-773. [ Links ]
DAFNI, A. 1992. Pollination Ecology. Oxford University Press Inc., New York. [ Links ]
DEL VALLE, HF. 1998. Patagonian soils: a regional synthesis. Ecologia Austral 8:103-123. [ Links ]
DIGGLE, PK. 1992. Development and evolution of plant reproductive characters. Pag. 397 en R Wyatt, editor. Ecology and evolution of plant reproduction. Chapman & Hall, New York. [ Links ]
HARDER, LD. 1988. Choice of individual flowers by bumble bees: interaction of morphology, time and energy. Behaviour 104:60-77. [ Links ]
HARDER, LD & SCH BARRETT. 1996. Pollen dispersal and mating patterns in animal-pollinated plants. Pag. 409 en DG Lloyd, & SCH Barrett, editores. Floral Biology. Studies on floral evolution in animal-pollinated plants. Chapman & Hall, New York. [ Links ]
HERRERA, CM. 1987. Composes of pollinator "quality": comparative analysis of a diverse insect assemblage. Oikos 50:79-90. [ Links ]
HERRERA, CM. 1989. Pollinator abundance, morphology, and flower visitation rate: analysis of the "quantity" component in a plant-pollinator system. Oecologia 80:241-248. [ Links ]
HERRERA, CM. 2000. Flower-to-seedling consequences of different pollination regimes in an insect-pollinated shrub. Ecology 81:15-29. [ Links ]
HURLBERT, SH. 1984. Pseudoreplication and the design of ecological field experiments. Ecological Monographs 54:187-211. [ Links ]
JORDANO, P; J BASCOMPTE & JM OLESEN. 2003. Invariant properties in coevolutionary networks of plant-animal interactions. Ecology Letters 6:69-81. [ Links ]
KANDORI, I. 2002. Diverse visitors with various pollinator importance and temporal change in the important pollinators of Geranium thunbergii (Geraniaceae). Ecological Research 17:283-294. [ Links ]
KEARNS, CA & DW INOUYE. 1993. Techniques for pollination biologists. University Press of Colorado, Niwot, Colorado. [ Links ]
KEARNS, CA & DW INOUYE. 1997. Pollinators, flowering plants, and conservation biology. Bioscience 47:297-306. [ Links ]
KLEIN, AM; I STEFFAN-DEWENTER & T TSCHARNTKE. 2003. Fruit set of highland coffee increases with the diversity of pollinating bees. Proceedings of the Royal Society 270:955-961. [ Links ]
KLINKHAMER, P & TJ JONG. 1990. Effects of plant density and sex differential reward on pollination visitation in the protandrous Echium vulgare (Boraginaceae). Oikos 57:399-405. [ Links ]
KWAK, M. 1987. Pollination and pollen flow disturbed by honeybees in bumblebee-pollinated Rhinanthus population? Pages 273-283 en J Van Andel, editor. Disturbance in grasslands. Dr. W. Junk, Dordrecht, Netherlands. [ Links ]
LEGENDRE, P & L LEGENDRE. 1998. Numerical Ecology. Elsevier. [ Links ]
LEÓN, RJC; D BRAN; M COLLANTES; JM PARUELO & A SORIANO. 1998. Grandes unidades de vegetación de la Patagonia extra andina. Ecologia Austral 8: 125-144. [ Links ]
LOVEJOY, T; ROJ BIERREGAARD; AB RYLANDS; CE QUINTELA; LH HARPER; KSJ BROWN; AH POWELL & GVN POWELL. 1986. Edge and other effects of isolation on Amazon forest fragments. Pag. 257-285 en ME Soulé, editor. Conservation biology: the science of scarcity and diversity. Sinauer, Sunderland, Massachusetts, USA. [ Links ]
MASCHINSKI, J & TG WHITHAM. 1989. The continuum of plant responses to herbivory: The influence of plant association, nutrient availability, and timing. American Naturalist 134:1-19. [ Links ]
MCNAUGHTON, SJ. 1983. Compensatory plant growth as a response to herbivory. Oikos 40:329-336. [ Links ]
MEMMOTT, J; NM WASER & MV PRICE. 2004. Tolerance of pollination networks to species extinctions. Proceedings of the Royal Society of London, Series B 271:2605-2611. [ Links ]
MILCHUNAS, D; O SALA & WK LAUENROTH. 1988. A generalized model of the effects of grazing by large herbivores on grassland community structure. American Naturalist 132:87-106. [ Links ]
MILLER, R; T SVEJCAR & N WEST. 1994. Implications of livestock herbivory in the intermountain sagebrush region: plant composition. Pag. 101-146 en M Vavra, WA Laycock, y R Pieper, editores. Ecological implications of livestock herbivory in the west. Society for Range Managment, Denver. [ Links ]
MIYASHITA, T; M TAKADA & A SHIMAZAKI. 2004. Indirect effects of herbivory by deer reduce abundance and species richness of web spiders. Écoscience 11:74-79. [ Links ]
MORRIS, WF. 2003. Which mutualists are most essential? Buffering of plant reproduction against the extinction of pollinators. Princeton University Press, Princeton. [ Links ]
MURCIA, C. 1995. Edge effects in fragmented forests: Implications for conservation. Trends in Ecology and Evolution 10:58-62. [ Links ]
MURCIA, C. 1996. Forest fragmentation and the pollination of neotropical plants. Pages 19-36 Forest Patches in tropical landscapes. Island Press, Washington, D.C., Covelo, California. [ Links ]
PAIGE, KN. 1992. Overcompensation in response to mammalian herbivory: from mutualistic to antagonistic interactions. Ecology 73:2076-2085. [ Links ]
PARUELO, JM; A BELTRÁN; E JOBBÁGY; OE SALA & RA GOLLUSCIO. 1998. The climate of Patagonia: general patterns and controls on biotic processes. Ecologia Austral 8:85-101. [ Links ]
SAUNDERS, D; R HOBBS & C MARGULES. 1991. Biological consequences of ecosystem fragmentation: a review. Conservation Biology 5:18-32. [ Links ]
SHIPLEY, B. 2000. Cause and Correlation in Biology: a user guide to path analysis, structural equations and causal inference. Cambridge University Press, Cambridge. [ Links ]
SIH, A & MS BALTUS. 1987. Patch size, pollinator behavior, and pollinator limitation in catnip. Ecology 68:1679-1690. [ Links ]
SOWIG, P. 1989. Effects of flowering plant´s patch size on species composition of pollinator communities, foraging strategies, and resource partitioning in bumblebees (Hymenoptera: Apidae). Oecologia 78:550-558. [ Links ]
STRAUSS, SY & WS ARMBRUSTER. 1997. Linking herbivory and pollination: new perspectives on plant and animal ecology and evolution. Ecology 78:1617-1618. [ Links ]
STRAUSS, SY; JK CONNER & SL RUSH. 1996. Foliar herbivory affects floral characters and plant attractiveness to pollinators: implications for male and female plant fitness. American Naturalist 147: 1098-1107. [ Links ]
TADEY, M. 2006. Grazing without grasses: Effects of introduced livestock on plant community composition in an arid environment in northern Patagonia. Applied Vegetation Science 9:109-116. [ Links ]
TADEY, M. 2007. Efectos del pastoreo sobre la polinización y reproducción de plantas del Monte Patagónico. Tesis de Doctorado. Universidad Nacional del Comahue, S. C. de Bariloche. [ Links ]
TADEY, M & AG FARJI-BRENER. 2007a. Indirect effects of exotic grazers: livestock decreases the nutrient content of refuse dumps of leaf-cutting ants through vegetation impoverishment. Journal of Applied Ecology 44:1209-1218. [ Links ]
TADEY, M & AG FARJI-BRENER. 2007b. Identifying direct and indirect effects of exotic grazers on native plant cover in the Monte desert of Argentina. Journal of Arid Environment 69:526–536. [ Links ]
VALLENTINE, JF. 2001. Grazing management. Academic Press, San Diego, CA, US. [ Links ]
VAUGHTON, G. 1992. Effectiveness of nectarivorous birds and honeybees as pollinators of Branksia spinulosa (Proteacea). Australian Journal of Ecology 17:43-50. [ Links ]
VÁZQUEZ, DP & D SIMBERLOF. 2003. Changes in interaction biodiversity induced by an introduced ungulate. Ecology Letters 6:1077-1083. [ Links ]
VÁZQUEZ, DP & D SIMBERLOFF. 2004. Indirect effects of an introduced ungulate on pollination and plant reproduction. Ecological Monographs 74:281-308. [ Links ]
VÁZQUEZ, DP & MA AIZEN. 2004. Asymmetric specialization: a pervasive feature of plant-pollinator interactions. Ecology 85:1251-1257. [ Links ]
WASER, NM. 1989. Optimal outcrossing in Ipomopsis aggregata: Seed set and offspring fitness. Evolution 43:1097-1109. [ Links ]
WATKINSON, AR & SJ ORMEROD. 2001. Grasslands, grazing and biodiversity: editors' introduction. Journal of Applied Ecology 38:233-237. [ Links ]

