










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.18 n.1 Córdoba ene./abr. 2008
TRABAJOS ORIGINALES
Impact of rainbow trout on aquatic invertebrate communities in subtropical mountain streams of northwest Argentina
Carlos Molineri *
INSUE-CONICET, Facultad de Ciencias Naturales e IML, San Miguel de Tucumán, Tucumán, Argentina.
* INSUE-CONICET, Facultad de Ciencias Naturales e IML, M. Lillo 205, San Miguel de Tucumán, 4000, Tucumán, Argentina. Email: cmolineri@csnat.unt.edu.ar
Recibido: 3 de agosto de 2007;
Fin de arbitraje: 17 de noviembre de 2007;
Revisión recibida: 17 de diciembre de 2007;
Aceptado: 20 de diciembre de 2007
ABSTRACT. The present study was conducted to assess the effect of rainbow trout introduction on benthic invertebrate communities, stratified as epibenthos and infauna. Two main questions are explored: 1) do the trout-invaded streams show a different community structure than the streams with autochthonous fish?, and 2) does the presence of trout affect differentially the epibenthos and the infauna? Epibenthic and infaunal samples, drift samples and fish stomach content were sampled four times from three stations to assess the impact of rainbow trout (Oncorhynchus mykiss) on the invertebrate aquatic fauna. Only one species of native fish was recorded in the studied sites, the siluriform Trichomycterus corduvense. Both fish species were found always in allopatry. The trout-invaded site has a different community structure than the other streams with a much lower abundance of large and active epibenthic taxa (e.g., Perlidae, Gripopterygidae, Hydropsychidae, Leptoceridae, Elmidae adults) and an increase in the importance of infaunal organisms (Chironomidae, Oligochaeta). Diversity indices showed alternating and opposite high and low values along time in trout-free and invaded sites.
Keywords: Benthos; Macroinvertebrates; Introduced species;Oncorhynchus mykiss;Trichomycterus corduvense.
RESUMEN. Impacto de la trucha arcoiris sobre comunidades de invertebrados acuáticos en ríos montañosos del noroeste argentino: Este estudio se realizó para evaluar el efecto de la introducción de trucha arcoiris en las comunidades de invertebrados bentónicos, estratificados en epibentos e infauna. Se exploran principalmente dos cuestiones: 1) si la estructura de la comunidad bentónica es diferente en el arroyo con truchas respecto a los arroyos con peces autóctonos; y 2) si la presencia de truchas afecta diferencialmente al epibentos y a la infauna. Se tomaron muestras del epibentos, infauna, deriva y contenido estomacal de peces en cuatro fechas y tres sitios para estudiar el impacto de la trucha arcoiris (Oncorhynchus mykiss) sobre la fauna de invertebrados acuáticos. Se registró solo una especie de pez nativo, el siluriforme Trichomycterus corduvense. Ambas especies de peces fueron encontradas siempre en alopatría. La estructura de la comunidad en el sitio con truchas resultó diferente de la de los otros arroyos con una menor abundancia de invertebrados epibentónicos grandes y activos (e.g., Perlidae, Gripopterygidae, Hydropsychidae, Leptoceridae, adultos de Elmidae) y una mayor importancia de organismos infaunales (Chironomidae, Oligochaeta). Los índices de diversidad presentaron valores altos y bajos alternados en el tiempo, y este patrón oscilante resultó opuesto en el sitio con truchas al de los sitios sin trucha.
Palabras clave: Bentos; Macroinvertebrados; Especie introducida;Oncorhynchus mykiss;Trichomycterus corduvense.
INTRODUCTION
Invasive species are currently seen as a dangerous hazard for receptor ecosystems (e.g., Simon & Townsend 2003), although specific studies on salmonids have shown contrasting results. In places where salmonids were autochthonous, fish depletion did not have a significant effect on benthos (Allan 1982), but some studies showed effects on the behavior of aquatic insect larvae (e.g., McIntosh et al. 2002) and in the structure of the epibenthic communities (Bechara et al. 1992, 1993). Where salmonids constitute invasive species (e.g., New Zealand, South America), the impact on benthos is more evident. Flecker (1992) noted impacts on the periodicity and density of invertebrate drift, and an effect on different ecological levels had been reported (Townsend 2003, Simon & Townsend 2003).
Salmonid introduction in Argentina dates back a century ago, but surveys about its impact on natural ecosystems are wanting (Vázquez & Aragón 2002). In Patagonia, a temperate area, at least seven salmonid species present stable populations (Pascual et al. 2002), but in the northernmost portion of Argentina, the subtropical NW, only one species of salmonid survives (Fernández & Fernández 1998), the rainbow trout Oncorhynchus mykiss (Walbaum). Here, relatively stable trout populations are found only above 1500 m a.s.l., as downstream temperatures are extremely high for salmonid survival. Salmonid impact in Argentina has been little studied but a negative effect is evident as autochthonous fish fauna suffered local extinction of selected species (Pascual et al. 2002) and a drastic reduction of invertebrate body size and abundance of selected functional feeding groups has been found (Buria et al. 2007).
Since the beginning of the 20th century, many rivers in NW Argentina, flowing through the mountain cloud forest (the "Yungas"), have been affected by rainbow trout periodic introductions. In this region, watercourses experience sudden spates during the rainy season (Hunzinger 1997). This natural disturbance (Sousa 1984) is a very important factor structuring invertebrate communities in general (e.g., Resh et al. 1988) and in the studied region in particular (Fernández et al. 2001). This alternation of unstable and stable periods is expected to influence the relative importance of abiotic and biotic factors controlling invertebrate communities (e.g., Flecker & Feifarek 1994; Jacobsen & Encalada 1998). During the dry season, the high stability of the flow regime results in higher richness, density and biomass (Flecker & Feifarek 1994). Depending on the time passed since the last substrate disturbance, the equitability within the community is expected to be low (due to short dry periods and dominance of fast-growing, opportunistic species) or high (due to longer dry periods and communities nearer equilibrium). The presence of trout in this scenario constitutes an additional, anthropogenic-related, disturbance (Resh at al. 1988). The relative importance of abiotic (spates) versus biotic (top predators) factors in structuring these communities is expected to vary between stable and disturbance periods. Predation has proved to be a strong modeling force (e.g., Glasser 1979); in aquatic ecosystems, this is true not only for exotic but also for native predatory fish and invertebrates (Bechara et al. 1992; Kohler 1992).
The aim of the present study was to assess the effect of rainbow trout introduction on stream benthos, stratified as epibenthos and infauna. This was done by comparing these invertebrate communities in three sampling sites: two close sampling sites in the Chaquivil stream (with cascades preventing trout invasion in the upstream site); and one in the adjacent Liquimayo stream, from a different, trout-free basin. The existence of neighbor basins, with and without trout, represents a natural experiment for testing some statements about trout impact on the benthic community. Only one autochthonous fish species (Trichomycterus corduvense Weyenberg, Siluriformes, Trichomycteridae) is present in the studied streams.
The main hypothesis to be tested is that the presence of trout alters the community structure of a stream. Bechara et al. (1993) suggest that high infaunal abundance may shade the effect of trout predation on epibenthos when studying complete surber samples. Spatial refuges, trapped detritus and higher substrate surface area offered by the profound benthos generally increase invertebrates densities (Flecker & Allan 1984). A second hypothesis to be tested here is that the effect will be detected on the epibenthos and not on the infauna.
MATERIAL AND METHODS
Study sites
Two drainage sub-basins were studied (Lules and Vipos) belonging to an endorrheic basin (Sali-Dulce), with a main north-south flow. The Sali-Dulce basin receives almost all the runoff waters of the densely populated Tucumán province. Annual mean precipitation varies between 1000 and 2000 mm, with 80% falling in the warmest months, December to March. The natural vegetation type of the upper part of both basins (>1000 m) is the mountain cloud forest dominated by alder (Alnus acuminata Humboldt, Bonpland & Kunth, Betulaceae) and belonging to the Yungas phytogeographic province of the Amazon domain (Cabrera & Willink 1973). For a complete zone characterization see Fernández & Molineri (2006).
Three sample sites were selected, with similar climate, hydrology, and composition and structure of the substrate and where the only native fish species present was Trichomycterus corduvense. Two on the Chaquivil stream (Vipos basin) and one on the Liquimayo (Lules basin, intensively studied in recent years (Fernández et al. 2002) because of the imminent construction of a dam) (Fig.1). Rainbow trout (Oncorhynchus mykiss) was first observed in the Chaquivil stream in 1989, but it was introduced before downstream of the drainage basin. The presence of cascades prevented upstream invasion until incidental transport of some individuals took place in 1989. Nowadays trout can be found up to an elevation of 2200 m a.s.l. Higher up, the existence of other cascades prevented further upstream trout movements. This fact allowed us to select two sampling sites, one with trout (Chaquivil-down at 2170 m a.s.l.) and the other trout-free (Chaquivil-up at 2260 m a.s.l), below and above these cascades, respectively. The third sampling site selected is on the trout-free Liquimayo stream, located at 2170 m a.s.l., its source being a high-mountain shallow lake (Huaca Huasi, at 4000 m a.s.l.).
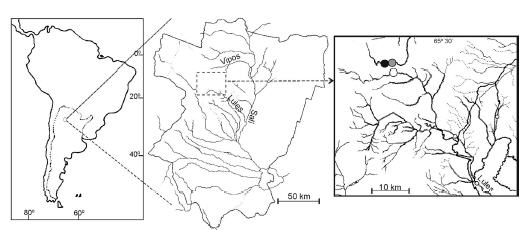
Figure 1. Study area and sampling sites. White circle=Liquimayo; black=Chaquivil-up; grey=Chaquivil-down.
Figura 1. Área de estudio y sitios de muestreo. Círculo blanco=Liquimayo; negro=Chaquivil-arriba; gris=Chaquivil-abajo.
Sampling
Liquimayo and Chaquivil-down were sampled four times (November 2002, June and September 2003, and January 2004); Chaquivil-up, was incorporated later and only sampled on the last two dates). Habitat parameters measured were water temperature, conductivity, mean width and depth, and discharge (by the flotation method, Hynes 1970) (Table 1). All sampling was carried out during low-discharge periods, except the last one.
Table 1. Environmental data (mean ± S.E.).
Tabla 1. Datos ambientales (media ± E.S.).

Benthos was sampled using a surber sampler of 300 µ mesh (0.09 m2 sample area). Three surber samples were taken at each site and date, in the riffle zone. Each sample was discriminated in superficial (epibenthos) and deep (infauna) portions. Epibenthos was obtained by cleaning the upper side of stones without removing the substrate underneath, the content of the net was then extracted and fixed. The infauna was obtained by removing a 10 cm-deep substrate layer in the marked site and conserved separately. Adults of aquatic insects were captured with light traps and emergence traps for taxonomic identifications. The samples were fixed with alcohol 96% but, formalin 4-10% was used when organic material was abundant in the sample.
A maximum of 10 individuals of each fish species were captured on each sampling date using nets and fishing-hooks; the digestive system was extracted and fixed. Stomach content was separated under magnification and identified to the highest possible resolution. Relative abundances of drifting invertebrates were calculated for Chaquivil-down from 40 samples (12 samples at each date, except 4 in January 2004) and for Liquimayo from 24 samples (12 samples at each date of Trichomycterus capture, November 2002 and January 2004). A drift net (300 µ mesh) was used to collect drifting organisms from the water surface and from the water column to a depth of 20 cm. Net content was extracted and fixed every 2 h during a 24 h period.
Samples were sorted out in the laboratory under a stereomicroscope at 10 and 20X, picking up all the specimens. Invertebrates were identified to species or morphospecies in Plecoptera, Ephemeroptera, Trichoptera, and Elmidae, and genus in Chironomidae. A few groups were identified to family level or higher taxonomical units (Table 2).
Table 2. Density (ind/m2) of taxa at three sampling stations (Liquimayo, Chaquivil-down, Chaquivil-up) and four sampling dates (November 2002; June 2003; September 2003; January 2004). Only taxa with >1% density (*) were included in the analyses.
Tabla 2. Densidad (ind/m2) de taxa de tres estaciones (Liquimayo, Chaquivil-abajo, Chaquivil-arriba) y cuatro fechas de muestreo (noviembre 2002; junio 2003; setiembre 2003; enero 2004). Sólo taxa con densidad >1% (*) se incluyeron en el análisis.
Data analysis
Richness and Diversity indices (Shannon-Weiner, Dominance, and PIE) were calculated using EcoSim (Gotelli & Entsminger 2001) for all complete samples (benthos: epibenthos + infauna) and subsamples (epibenthos and infauna, separately). Richness is given as total number of taxa (species or morphospecies). Hurlbert´s (1971) PIE index (Probability of Interspecific Encounter) was used as indicative of equitability. Dominance (D) was estimated as the proportion of the dominant taxon. Statistical significances using T test (P<0.05) were calculated based on the variability between the three replicates of each sample. Further data analyses were carried out by comparing the abundance of each taxon at each sampling site and date; differences (P<0.05) were tested using Mann-Whitney U test with INFOSTAT (2004). Comparisons between all possible pair of sites (Liquimayo/Chaquivil-down, Liquimayo/Chaquivil-up and Chaquivil-down/Chaquivil-up) were done. Fish stomach content is given as percentage occurrence (%F) and percentage abundance (%A) of each prey type (Oscoz et al. 2005):
%Fi=(Ni/N)x100 ,
where Ni is the number of predators consumed by prey i and N is the total number of studied predators
%A=(Pi/P)x100 ,
where Pi is the abundance of prey i in all stomach contents and P is the total abundance of all prey items consumed.
The ADE software (Thioulouse et al. 1995) was used to explore multivariate data structure. Infrequent taxa (less than 1% of total abundance) were ignored and abundances were log-transformed (log x+1) to normalize data structure. Correspondence analysis (CA, Greenacre 1984) was used to analyze: complete surber samples (three for each sampling date and station, n=26), and disaggregated surber samples (separated in epibenthos and infauna, n=58). Six sample typologies were considered: (1) from streams with or without trout; (2) epibenthic or infaunal samples; (3) samples per site; (4) samples per date; (5) samples per river; and (6) samples per altitude. Then, Within and Between-Group analyses (Dolédec & Chessel 1989) were conducted for each typology, to better visualize possible differences between groups of samples (typology 2 was only analyzed with disaggregated surber samples). In the Between-Group analysis, the processed table contains the center of gravity of each group (means of variable and groups). The Within-Group analysis is a linear ordination method for removing the effect of a qualitative variable. The significance of these differences was evaluated by Monte-Carlo tests, with 100000 random permutations.
The deficiency in spatial replication in this study (two trout-less reaches and one trout-invaded reach) is mitigated by the similarity in physical and chemical characteristics of the sampled streams, and most importantly by the fact that Chaquivil-down (with trout) and Chaquivil-up (trout-free) are contiguous segments of the same stream (i.e., the presence of trout being the only difference). Moreover, the inference about trout effect is drawn using other premises: 1) difference in infauna and epifauna, 2) the increasing differences during low water (i.e., stable) periods, and 3) the fish stomach contents.
RESULTS
Macroinvertebrates densities
Austrelmis spp. (larvae, Elmidae) was the first (occasionally, the second) most abundant taxon in all sites and dates. At Chaquivil-down (with trout), in all sampling dates, a Chironomidae (most of the times Cricotopus spp., Orthocladiinae) or Pristina sp. (Oligochaeta, Naididae) was the second most abundant taxon. At Liquimayo and Chaquivil-up (both trout-free) most abundant taxa were, in order of importance: Glossosomatidae, Andesiops peruvianus and Camelobaetidius penai (Baetidae), Nectopsyche sp. (Leptoceridae), and Smicridea spp. (Hydropsychidae); Cricotopus spp. was of some importance in Chaquivil-up (Table 2). Some taxa also showed significant differences in abundance between trout-free sites (Fig. 2), Leptohyphes eximius, Smicridea spp., Claudioperla tigrina, Pseudochironomus sp., adults of Austrelmis spp., and Anacroneuria spp.
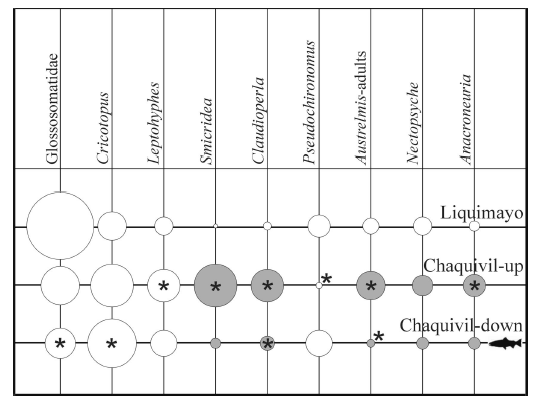
Figure 2. Differences in densities (circle size) of selected taxa between sampling-sites. Significant difference (Mann-Whitney U test, p <0.05) between Liquimayo and the other two marked with "*", significant differencebetween Chaquivil-down and Chaquivil-up marked with gray circles.
Figura 2. Diferencias en densidades (tamaño de los círculos) de algunos taxones entre los sitios de muestreo. Diferencias significativas (Mann-Whitney test U, p < 0.05) entre Liquimayo y los otros dos marcada con "*", diferencias entre Chaquivil-arriba y Chaquivil-abajo marcada con círculos grises.
When comparing the "trout" reach (Chaquivil-down) with the immediately upstream trout-free reach (Chaquivil-up), significant (P<0.05 U test) lower densities of Smicridea spp., Claudioperla tigrina, adults of Austrelmis spp., Nectopsyche sp., and Anacroneuria spp. were found (Fig. 2). Glossosomatidae and Leptohyphes eximius also decreased in the presence of trout, but not significantly.
Seasonality was evident in some taxa, especially in Camelobaetidius penai (frequent in spring-summer, but absent in winter samples), and with an opposite pattern (only collected on June and September 2003), Paraheptagyia sp. (Chironomidae, Diamesinae) or Orthocladiinae 3 (only collected at all sites on September 2003). By contrast, other taxa were common all year round (e.g., Leptohyphes eximius, Andesiops peruvianus, and Nectopsyche sp.).
Macroinvertebrate richness and diversity
Taxonomic richness did not vary between trout and trout-free sites (Fig. 3a). Overall, richness was higher in the dry period for all sites (June and September 2003; Fig. 3a). The pattern was similar when analyzing benthic strata separately, although infauna showed higher richness and a less pronounced increment through time than epibenthos.

Figure 3. Diversity indices (three replicates, whole surbers). a: Richness; b: Shannon; c: PIE; d: Dominance.
Figura 3. Índices de diversidad (tres replicas, surbers completos). a: Riqueza; b: Shannon; c: PIE; d: Dominancia.
Dominance (D), Shannon diversity (H´) and Hurlbert's PIE indices, showed oscillating patterns (Figs. 3b-d). The three indices, at each site, were found to oscillate from date to date but patterns were different for trout-free and trout-invaded reaches. The trout-free sites Chaquivil-up and Liquimayo (Figs. 3b-d) showed similar tendencies, while Chaquivil-down showed the opposite pattern. This temporal fluctuation is also observable when benthic strata are analyzed separately.
Macroinvertebrate communities
The CA of complete surber samples yielded the factorial maps shown in Figs. 4a-b. Overall, this description was the same as the one obtained when analysing separate infauna and epibenthos samples. Figure 4a depicts sample distribution in the first factorial plane, which explains 48.8% of data variability. Samples are discriminated by date of sampling, each one of the four sampling dates occupying one quadrant on the first factorial plane. The first axis highest contributions (Fig. 4b) are those of Orthocladiinae 3, Paraheptagyia sp., Claudioperla tigrina, Diptera-pupae and Tanytarsus sp. (positive side); and Camelobaetidius penai, Atopsyche spp., and Ceratopogonidae (negative side). Important contributions to the second axis (Fig. 4b) are Hydrachnidia and Pseudochironomus sp. in the positive side; and Anacroneuria spp., Smicridea spp., adults of Austrelmis spp., and Tipulidae in the negative side. There is no evidence of differences between with-trout and trout-free sites (Fig. 4a).

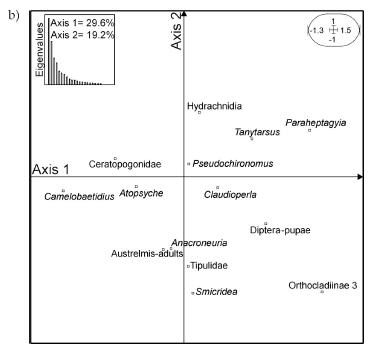
Figure 4. Faunal description (Correspondence Analysis): a: samples grouped by sampling site and season; b: invertebrates. Arrows show the temporal sequence between sampling dates. Circles correspond to mean of three replicates (dots) for each of the three sampling sites.
Figura 4. Descripción faunística (Análisis de Correspondencia): a: muestras agrupadas por sitio y fecha de muestreo; b: invertebrados. Las flechas indican la secuencia temporal entre fechas de muestreo. Los círculos representan la media de las tres réplicas (puntos) para cada uno de los tres sitios.
Differences between groups of samples for every typology (1 to 6) were highly significant (P<0.01), except for samples from trout-invaded /trout-free stations when surbers were treated as a unit (P=0.06) or when infaunal samples were analyzed alone (P=0.08). Analysis including epibenthic samples only, successfully discriminated between trout-invaded and trout-free sites (P=0.02).
The first factorial plane of the Between-Group analysis explained 84% of the total variation. The best-discriminated category was "samples per date" (Fig. 5a). This temporal effect was removed through the Within-Group analysis, as shown in Fig. 5b, where the groups of sampling dates are centered (all centers of gravity are positioned at the origin). As a result, a typology of the samples, released from the temporal effect, is obtained (Figs. 5c-d). "Trout-free" samples (from Liquimayo and Chaquivil-up) are significantly different from those of the trout invaded site (Chaquivil-down), as shown by the centers of gravity of both groups of samples (Fig. 5c). The other typologies showed also significant differences, "samples per site" is shown as an example (Fig. 5d).

Figure 5. Between and Within-group Correspondence Analysis: a: first factorial plane of the between-group analysis, depicting sample distribution; b-d: idem from the within-group analysis; b: all centers of gravity of date category are at the origin, showing that temporal effect was removed (see text for explanation), c: groups of samples from trout-free sites and the trout site, d: samples from each sampling station.
Figura 5. Análisis de correspondencia Entre y Dentro de grupos: a: primer plano factorial del análisis Entre-grupos, mostrando la distribución de las muestras de cada sitio; b-d: ídem, a partir del análisis Dentro de grupos: b: todos los centros de gravedad de la categoría "fecha" están en el origen, mostrando la eliminación del efecto temporal (ver texto), c: segregación de grupos de muestras provenientes de sitios con y sin truchas, d: segregación de los grupos de muestras provenientes de cada uno de los tres sitios estudiados.
The contradiction that trout-samples are not different from trout-free samples in the CA, but they do in the Between-Within-Group analyses, is only apparent because in the CA the temporal effect is so strong as to shade all other possible comparisons (Dolédec & Chessel 1989). When the temporal effect is removed (with the Within-group analysis), an effect attributable to differences in the reaches (absence/presence of trout) is evidenced (Figs. 5c-d).
The distinction of both portions of benthic samples (epibenthos and infauna) was not significant with log-transformed data. This seems logical since the main difference between them (abundance) is reduced by logarithmic transformation. As expected, analyses with non-transformed data significantly discriminated between epibenthos and infauna.
Fish stomach content
Only two species of fish were present in the studied sites, the autochthonous siluriform Trichomycterus corduvense and the introduced rainbow trout (Oncorhynchus mykiss), never found in sympatry. Twelve individuals of Trichomycterus corduvense were captured in Liquimayo (7 in November 2002, 5 in January 2004). This species was not captured in Chaquivil-down nor in Chaquivil-up, but was observed in this last site. This native fish is only active during the night, using mechanical cues to detect preys. Benthic organisms such as Baetidae, some Orthocladiinae and Simuliidae were the most frequently consumed items (Table 3).
Table 3. Relative abundance of invertebrates in infauna (%Ai), epibenthos (%Ae), drift (%Ad) and fish stomach content (%A). Relative frequency (%F) of invertebrates in fish stomach content is also given. Trout=Oncorhynchus mykiss (n=26); native=Trichomycterus corduvense (n=12).
Tabla 3. Abundancia relativa de invertebrados en infauna (%Ai), epibentos (%Ae), deriva (%Ad), y contenido estomacal de peces (%A), y frecuencia relativa (%F) de invertebrados en contenido estomacal de peces. Trout=Oncorhynchus mykiss (n=26); native=Trichomycterus corduvense (n=12).
Twenty-six trout (Oncorhynchus mykiss) sto-mach contents from the only site were it is pre-sent (Chaquivil-down) were analyzed (11 from November 2002, 1 from June 2003, 8 from September 2003, 6 from January 2004). Trouts are active during the day, since prey capture relies mainly on vision (e.g., Flecker & Allan 1984). The most frequent items in their stomach content were from drift and benthos: Simuliidae, Gripopterygidae, Chironomidae, Trichoptera, Elmidae, and aerial arthropods (Table 3). Fish or other vertebrate remains were absent. Some plant material like seeds and moss fragments were occasionally found.
DISCUSSION
Effect of fish on the benthic community
The results strongly suggest a trout-effect on the benthic community. Overall, this effect is reflected in the lower density (Fig. 2) of some large epibenthic taxa (e.g., Smicridea spp., Claudioperla tigrina, adults of Austrelmis spp., Nectopsyche sp., Anacroneuria spp., Glossosomatidae). The higher abundance of some infaunal taxa (Cricotopus spp., Pseudochironomus sp., Pristina sp.) may be an indirect effect of trout presence caused by elimination of invertebrate competitors and both, invertebrate and vertebrate (native fish) predators (Townsend 2003). The most frequent taxa in the trout stomach content were from the epibenthos (Claudioperla tigrina, Chironomidae in part, Nectopsyche sp., Smicridea spp., Glossosomatidae, Simuliidae and Austrelmis spp.) and drift (Chironomidae in part, first instars of Hydroptilidae and aerial arthropods). The dietary profile was found to be similar to that of other regions (e.g., Palma et al. 2002). In general, species preyed by trout (large and/or active animals) are less abundant in Chaquivil-down. This pattern is commonly found in studies on the impact of invasive trout (i.e., Townsend 2003; Buria et al. 2007).
Taxa that showed differences in abundance between both trout-free sites are probably reflecting undetected differences between both basins that would be explained by additional sampling in higher sites of the Liquimayo stream.
The native fish (Trichomycterus corduvense) consumed a high percentage of epibenthic taxa such as Baetidae (Camelobaetidius penai, Baetodes spp., Andesiops peruvianus), Orthocladiinae (Cricotopus spp. and Orthocladius sp.) and Simuliidae, while drifting organisms were less frequent. Its diet has received some attention (e.g., Ferriz 1998) but quantitative studies of its impact on benthos are lacking. Low density and biomass of the studied populations probably prevent a high impact on aquatic invertebrates. Dissimilarities between invertebrate communities from the contiguous stream reaches Chaquivil-down (with trout) and Chaquivil-up (with native fish) suggest that the native night-active (Ferriz 1998) predator produces a minor impact on invertebrate fauna. The preys most consumed by Trichomycterus do not show lower abundance in Liquimayo or in Chaquivil-up.
Different effects produced by introduced visual vs. native non-visual predators have been previously reported, for example for brown trout and galaxids in New Zealand (Townsend 2003). Galaxids can cause only a weak trophic cascade while trout have a greater effect. Results of the present study also suggest a greater impact of rainbow trout on invertebrates when compared to the influence of Trichomycterus.
Abiotic vs. biotic impact on benthos
The effect of trout was shaded in the rainy season (January 2004) by major spates. Invertebrate communities were strongly depleted by scouring in all sites (with and without trout), and thus exotic predator effects were undetectable. This temporal pattern caused by hydraulic conditions, and reflected in the CA, is well known in river ecology (Resh et al. 1988). The results obtained in relation to the taxonomic composition of the invertebrate community, coincide with other published surveys from Andean streams at comparable altitudes (Flecker & Feifarek 1994; Jacobsen & Encalada 1998; Rocabado & Wasson 1999; Fernández et al. 2001, 2002), showing the importance of the immature stages of various insect orders (Coleoptera, Trichoptera, Ephemeroptera, Plecoptera, and Diptera). Besides spates, the presence of taxa with highly seasonal life cycles (Camelobaetidius penai, Paraheptagyia sp. and Orthocladiinae 3) also contributes to the temporal pattern observed. Restricted life cycles are largely unknown for Neotropical aquatic invertebrates and it seems that multivoltine species with adults emerging all year round is the most common reported pattern.
Diversity measures
Taxonomic richness was not reduced by the presence of trout, due to the compensation produced by the positively affected groups (e.g., infaunal Diptera), a similar situation was found in Patagonian streams by Buria et al. (2007). Values of D, H´, and PIE indices showed different values in the trout-reach, when compared to the other sites. Temporal oscillation showed by D, H´, and PIE indices at each sampling station denotes the dynamic nature of the benthic community. Such an alternating pattern of consecutively high and low equitability values was also found by Jacobsen & Encalada (1998) in high-Andean Ecuadorian streams. This situation may be predicted from theoretical ecology of disturbances [dynamic equilibrium model and intermediate disturbance hypothesis, as applied to stream ecology (Resh et al. 1988)]: disturbances (e.g., scouring in the rainy season) favor r-selected species in after-disturbance communities, but depending on the time elapsed, a more equitable community may develop through biotic interactions (e.g., competition, predation).
A remarkable result of the present study is the opposite nature of evenness oscillation through time, when comparing trout-related with trout-free sites (Figs. 3b-d). Low abundances of Glossosomatidae in the trout-site in June 2003 increased equitability (decreased D, and increased PIE and H´ indices). However, the extremely high numbers of Glossosomatidae in Liquimayo (trout-free) on the same date reduced evenness values (high D, and low PIE and H´). On the other hand, in September 2003, evenness was low at Chaquivil-down (trout invaded) due to the dominant infaunal Chironomidae Cricotopus spp., while epibenthic organisms were much less important (unlike in trout-free sites), as expected from a longer-term interaction with the invasive predator (Bechara et al. 1993) and the cascading effect from the reduction of herbivores (Kohler 1992).
The significant variations of H´, D, and PIE indices between dates may be caused by different factors (i.e., spates, predation, competition). During stable periods (i.e., low discharge), trout predation seems to be an important agent of community structure. Trout stomach contents confirm this pattern, showing high frequency of epibenthic and aquatic drift fauna during low flow periods, but higher importance of terrestrial fauna in the rainy season. During unstable periods, benthic community structure depends more on the importance of abiotic factors such as frequency and intensity of spates, as indicated by the reduction in richness and abundance during the rainy season. Cascading effects (e.g., on epilithic algae biomass), invertebrate competition, and predation may also become important factors during longer periods of physical stability (e.g., at the end of the dry season).
Results of the present work sustain the hypothesis of Bechara et al. (1993) that salmonids should impact more strongly on epibenthos than on infauna since significant differences attributable to trout presence were found by CA only when epibenthic portions were treated separately (i.e., epibenthos alone, or together with infauna but as disaggregated samples). No trout effect was evident from the analysis of infauna alone, or when both portions (infauna and epibenthos) were clumped together as a whole "benthic" sample. Moreover, the most affected taxa were the large and epibenthic Perlidae, Gripopterygidae, Hydropsychidae, Leptoceridae and Elmidae adults.
ACKNOWLEDGEMENTS
I am indebted to the Wilde family for allowing me to work at Chaquivil stream. I am greatly thankful to H. Fernández, O. Fossati, E. Domínguez, R. Albariño and two anonymous reviewers for constructive discussion and revision of the manuscript. W. Villafañe and I. Ferro provided invaluable ideas and help during field trips. I am grateful to I. Ferro for suggesting this study and to M. Orce for expertly processing the bulk of the samples. I thank V. Buzza, G. Cuezzo and M. Molineri for their generous assistance in various aspects of the present work. The author belongs to the National Council of Scientific Research of Argentina (CONICET), support of which is greatly acknowledged. This work was partially financed by PICT 01-12529 and CIUNT 26-G211.
REFERENCES
ALLAN, JD. 1982. The effects on reduction in trout density on the invertebrate community of a mountain stream. Ecology 63:1444-1455. [ Links ]
BECHARA, JA; G MOREAU & L HARE. 1993. The impact of brook trout (Salvelinus fontinalis) on an experimental stream benthic community: the role of spatial and size refugia. J. Anim. Ecol. 62: 451-464. [ Links ]
BECHARA, JA; G MOREAU & D PLANAS. 1992. Top-Down Effects of Brook Trout (Salvelinus fontinalis) in a Boreal Forest Stream. Can. J. Fish. & Aquat. Sc. 49:2093-2103. [ Links ]
BURIA, L; R ALBARIÑO; V DÍAZ VILLANUEVA; B MODENUTTI & E BALSEIRO. 2007. Impact of exotic rainbow trout on the benthic macroinvertebrate community from Andean-Patagonian headwater streams. Arch. Hydrobiol. 168:145-154. [ Links ]
CABRERA, AL & A WILLINK. 1973. Biogeografía de América Latina. Programa Regional de Desarrollo Científico y Tecnológico, OEA, Serie monográfica N° 13. [ Links ]
DOLÉDEC, S & D CHESSEL. 1989. Rythmes saisonniers et composantes stationnelles en milieu aquatique II. Prise en compte et élimination d`effets dans un tableau faunistique. Acta Oecologica, Oecologica Generalis 10:207-232. [ Links ]
FERNÁNDEZ, HR & LA FERNÁNDEZ. 1998. Introduction of Rainbow Trout in Tucumán Province, Argentina: Problems and Solutions. Ambio 27: 584-585. [ Links ]
FERNÁNDEZ, HR & C MOLINERI. 2006. Toward a sustainable experience in an intermountain valley in northwestern Argentina. Ambio 35:262-266. [ Links ]
FERNÁNDEZ, HR; F ROMERO; M PERALTA & L GROSSO. 2001. La diversidad del zoobentos en ríos de montaña del noroeste de Argentina: comparación entre seis ríos. Ecología Austral 11:9-16. [ Links ]
FERNÁNDEZ, HR; F ROMERO; MB VECE; V MANZO; C NIETO & M ORCE. 2002. Evaluación de tres índices bióticos en un río subtropical de montaña (Tucumán – Argentina). Limnetica 21:1-13. [ Links ]
FERRIZ, AR. 1998. Alimentación de Trichomycterus corduvense Weyenbergh,1879 (Teleostei: Trichomycteridae) en dos ríos Serranos de San Luis, Argentina. Hidrobiología 8:43-49. [ Links ]
FLECKER, AS. 1992. Fish predation and the evolution of invertebrate drift periodicity: evidence from Neotropical streams. Ecology 73:438-448. [ Links ]
FLECKER, AS & JD ALLAN. 1984. The importance of predation, substrate and spatial refugia in determining lotic insect distributions. Oecologia 64:306-313. [ Links ]
FLECKER, AS & B FEIFAREK. 1994. Disturbance and the temporal variability of invertebrate assemblages in two Andean streams. Freshwat. Biol. 31:131-142. [ Links ]
GLASSER, JW. 1979. The role of predation in shaping and maintaining the structure of communities. The American Naturalist 113:631-641. [ Links ]
GOTELLI, NJ & GL ENTSMINGER. 2001. EcoSim: Null models software for ecology. Version 7.0. Acquired Intelligence Inc. & Kesey-Bear. [ Links ]
GREENACRE, M. 1984. Theory and applications of correspondence analysis. Academic Press, London. [ Links ]
HURLBERT, SH. 1971. The nonconcept of species diversity: a critique and alternative parameters. Ecology 52:577-586. [ Links ]
HUNZINGER, H. 1997. Hydrology of the montane forest in the Sierra de San Javier, Tucumán, Argentina. Mountain Research and Development 17:299-308. [ Links ]
HYNES, HBN. 1970. The Ecology of Running Waters. Liverpool University Press, Liverpool. [ Links ]
INFOSTAT. 2004. InfoStat (versión 2004). Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. [ Links ]
JACOBSEN, D & A ENCALADA. 1998. The macroinvertebrate fauna of Ecuadorian high-land streams in the wet and dry season. Archiv Hydrobiol. 142:53-70. [ Links ]
KOHLER, SL. 1992. Competition and the structure of a benthic stream community. Ecological Monographs 62:165-188. [ Links ]
MCINTOSH, AR; BL PECKARSKI & BW TAYLOR. 2002. The influence of predatory fish on mayfly drift: extrapolating from experiments to nature. Freshwater Biol. 47:1497-1513. [ Links ]
OSCOZ, J; PM LEUNDA; F CAMPOS; MC ESCALA; C GARCÍA-FRESCA & R MIRANDA. 2005. Spring diet composition of Rainbow Trout, Oncorhynchus mykiss (Walbaum, 1792), in the Urederra River (Spain). Annales de Limnologie - International Journal of Limnology 41:27-34. [ Links ]
PALMA, A; R FIGUEROA; VH RUIZ; E ARAYA & P BERRIOS. 2002. Composición de la dieta de Oncorhynchus mykiss (Walbaum 1792) (Pisces: Salmonidae) en un sistema fluvial de baja intervención antrópica: estero Nonguen, VIII Region, Chile. Gayana 66:129-139. [ Links ]
PASCUAL, M; P MACCHI; J URBANSKI; F MARCOS; C RIVA ROSSI; M NOVARA & A DELL'ARCIPRETE. 2002. Evaluating potential effects of exotic freshwater fish from incomplete species presence-absence data. Biological Invasions 4:101-113. [ Links ]
RESH, VH; AV BROWN; AP COVICH; ME GURTZ; HW LI; GW MINSHAL, ET AL. 1988. The role of disturbance in stream ecology. J. N. Am. Benthol. Soc. 7:433-455. [ Links ]
ROCABADO, G & JG WASSON. 1999. Regionalización de la fauna bentónica en la cuenca andina del río Beni (Bolivia). Rev. Bol. Ecol. 6:121-132. [ Links ]
SIMON, KS & CR TOWNSEND. 2003. Impacts of freshwater invaders at different levels of ecological organisation, with emphasis on salmonids and ecosystem consequences. Freshwater Biol. 48: 982-994. [ Links ]
SOUSA, WP. 1984. The role of disturbances in natural communities. Ann. Rev. Ecol. Syst. 15:353-391. [ Links ]
TOWNSEND, CR. 2003. Individual, population, community, and ecosystem consequences of a fish invader in New Zealand streams. Conservation Biology 17:38-47. [ Links ]
THIOULOUSE, J; D CHESSEL; S DOLÉDEC; JM OLIVIER; F GOREAUD & R PELISSIER. 1995-2000. ADE4: Ecological Data Analysis: Exploratory and Euclidean methods in Environmental sciences. Version 2001, CNRS, Lyon, France (available at http://pbil.univ-lyon1.fr/ADE-4). [ Links ]
VÁZQUEZ, DP & A ARAGÓN. 2002. Introduction. Biological Invasions 4:1-5. [ Links ]

