










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.18 n.2 Córdoba mayo/ago. 2008
TRABAJOS ORIGINALES
Características del néctar de Cologania broussonetii (Balb.) DC. (Fabaceae) y su relación con los visitantes florales
Mariana L Musicante1 & Leonardo Galetto2, *
1. CONICET - Centro de Investigaciones Entomológicas de Córdoba, Instituto Multidisciplinario de Biología Vegetal, Universidad Nacional de Córdoba, Argentina.
2. CONICET - Grupo de investigaciones en ecología reproductiva de plantas, Instituto Multidisciplinario de Biología Vegetal, Universidad Nacional de Córdoba, Argentina.
* CONICET - Centro de Investigaciones Entomológicas de Córdoba - Instituto Multidisciplinario de Biología Vegetal, Univ. Nacional de Córdoba, Casilla de Correo 495, 5000 Córdoba, Argentina. Email: marymusicante@yahoo.com.ar
Recibido: 22 de febrero de 2008;
Fin de arbitraje: 13 de mayo de 2008;
Revisión recibida: 1 de julio de 2008;
Aceptado: 15 de julio de 2008
RESUMEN. La relación entre las plantas con flores y sus polinizadores es influenciada por la cantidad y calidad de la recompensa ofrecida. En este trabajo se estudiaron las características del néctar de Cologania broussonetii (Fabaceae) considerando la composición de azúcares, la dinámica de su producción a lo largo de la vida de la flor y el efecto de las extracciones sucesivas de néctar sobre la secreción y la relación de estos caracteres con sus polinizadores. La composición de azúcares del néctar reveló la presencia de sacarosa, fructosa y glucosa (62, 24 y 14%, respectivamente). La secreción de néctar fue continua durante el primer día de vida de la flor y luego se detuvo; cuando el néctar no fue extraído, la flor lo reabsorbió activamente durante el segundo día. Las extracciones sucesivas (i.e., visitas) no afectaron la cantidad total de néctar ni su ritmo de producción. Las flores de C. broussonetii fueron visitadas sólo por dos especies de abejorros: Bombus opifex y Bombus bellicosus (Hymenoptera; Apidae), ambas en baja frecuencia. Las particularidades encontradas en el modo de secreción del néctar en esta planta pueden relacionarse con polinizadores que visitan las flores en baja frecuencia. Es decir, considerando las características de la producción de néctar en esta especie sería razonable especular que el patrón encontrado tendría valor adaptativo frente a condiciones de visitación variables e impredecibles.
Palabras clave: Faboideae; Bombus sp.; Composición de azúcares del néctar; Patrón de secreción del néctar.
ABSTRACT. Nectar characteristics of Cologania broussonetii (Balb.) DC. (Fabaceae) and its relationship with flower visitors: Relationships between flowering plants and their pollinators are usually affected by the amount and quality of reward, mainly pollen or nectar, offered to pollinators. Here we analyzed nectar sugar composition, nectar secretion pattern, the effects of removal of nectar on the secretion pattern, and the relationships between these variables and the frequency of visits of the pollinators in Cologania broussonetii (Fabaceae). Nectar sugar composition showed sucrose 62%, fructose 24% and glucose 14%. Nectar secretion resulted continuous through the first day after flower opening. When flower nectar was not removed, during the second day of flower lifetime the sugar content declined due to nectar reabsorption. Nectar removal did not affect the total amount of sugar produced by the flowers. Flowers were visited in low frequencies just by two bumblebees: Bombus opifex and Bombus bellicosus (Hymenoptera: Apidae). Some of the nectar secretion traits can be related with a low frequency of pollinator visits. Thus, considering the nectar secretion pattern in these species seem to be in accordance with variables and unpredictable visit rates.
Keywords: Faboideae; Flower visitors; Bombus sp.; Sugar nectar composition; Nectar secretion pattern.
INTRODUCCIÓN
Las plantas con flores han desarrollado diversas estrategias para atraer a los agentes polinizadores y asegurarse repetidas visitas que conducirán a la polinización (Simpson & Neff 1983). El uso de recompensas, como el néctar y el polen, es la estrategia más utilizada y difundida (Faegri & Van der Pijl 1971; Dafni 1992). Los azúcares presentes en el néctar lo convierten en la principal fuente de energía para varios grupos de polinizadores. Además, aporta pequeñas cantidades de fenoles, aminoácidos, ácidos reductores, lípidos, proteínas y alcaloides (Baker & Baker 1983). El comportamiento de los polinizadores (frecuencia de visitas y movimiento entre las flores) es influenciado por la calidad y la cantidad de recompensa que encuentren en las flores (i.e., concentración de azúcares y volumen del néctar) (Waser 1983; Real & Rathcke 1988), por lo cual se espera que las plantas ajusten la provisión de la recompensa (i.e., secreción del néctar) en relación a los recursos maternos disponibles y a las características del polinizador más frecuente (e.g., Torres & Galetto 1998). Características del néctar tales como el volumen total producido y la concentración de azúcares se relacionan con determinados grupos o gremios de polinizadores. Por ejemplo, Opler (1983), en una comunidad de Costa Rica, encontró una relación positiva entre la producción de néctar y el tamaño corporal de los polinizadores. Por su parte, Baker & Baker (1983) encontraron una relación entre la composición de azúcares del néctar floral y el gremio al que pertenecen los polinizadores (i.e., murciélagos del nuevo mundo, escarabajos, abejas de trompa larga, picaflores, etc.). Sin embargo, también existe evidencia que indica que las particularidades del néctar en ciertas familias y géneros de plantas podrían estar más relacionadas con restricciones filogenéticas que con sus polinizadores (Galetto & Bernardello 2003 y bibliografía allí citada).
La naturaleza exhibe distintos patrones de secreción de néctar. Las plantas pueden secretar néctar en forma continua desde la antesis hasta la senescencia de la flor, reponerlo luego de cada visita de un polinizador, o bien interrumpir su secreción luego de una extracción (e.g., Galetto & Bernardello 1992, 1993, 1995, 2004; Bernardello et al. 1994; Galetto et al. 1994, 2000; Torres & Galetto 1998). Las variaciones en la composición de azúcares y en el patrón de secreción de néctar durante la antesis floral afectan, en parte, el comportamiento de los polinizadores ante las flores (Galetto et al. 1994, 2000). Esto, a su vez, tiene consecuencias sobre el flujo de polen dentro y entre individuos y sobre la producción de frutos y semillas de las plantas (Herrera 1987). Si bien las Fabaceae están representadas por cientos de especies en la Argentina y sólo se dispone de datos sobre las características del néctar para unas pocas de ellas, resulta interesante analizar las tendencias generales que pueden vislumbrarse. Así, estudios previos sobre el néctar de Fabáceas nativas de Argentina muestran que la composición de azúcares está dominada principalmente por monosacáridos, mientras que la sacarosa está ausente o es muy escasa (Galetto & Bernardello 2003), la producción es continua durante toda la vida de la flor y el néctar no es reabsorbido (Cocucci et al. 1992; Galetto et al. 2000). La mayoría de estas especies son visitadas por distintos grupos de Apoidea, aunque también algunas son visitadas por mariposas nocturnas y aves (Galetto & Bernardello 2003).
Cologania (Fabaceae, Faboideae) es un género americano que se extiende desde Estados Unidos hasta Argentina central. C. broussonetii (Balb.) DC. es la única especie que crece en la Argentina. Se distribuye desde la Provincia de San Luis, sierras de Córdoba, Jujuy y Tucumán hasta los Andes orientales en Bolivia (Burkart 1952). Es una hierba perenne, cuyos tallos volubles alcanzan unos 150 centímetros de longitud. Sus flores se disponen en grupos de dos a tres por nudo y ocupan los últimos cinco a seis nudos de las ramas, miden de 17 a 22 mm de largo, son papilionadas, zigomórficas, fucsias, y vistosas. C. boroussonetii produce néctar como recompensa para sus agentes polinizadores (Musicante & Galetto 2008), el nectario se encuentra sobre la base del receptáculo floral y, como ocurre en muchas flores papilonadas, el néctar se acumula en la base de la corola. La recompensa queda accesible a los polinizadores cuando estos presionan la quilla e introducen su probóscide; al mismo tiempo, la columna estaminal y el estigma emergen depositando el polen en el cuerpo de los polinizadores (Arroyo 1981). En la población estudiada C. boroussonetii fue atractiva solo para dos especies de abejorros del género Bombus que visitan sus flores con baja frecuencia (Musicante & Galetto 2008). C. boroussonetii produce frutos por autopolinización espontánea, lo que indica que tiene la capacidad de producir semillas sin la intervención de los animales polinizadores (Musicante & Galetto 2008). Sin embargo, el papel de los polinizadores es importante ya que provocan un incremento significativo en el porcentaje de fructificación y el valor de éxito reproductivo (Musicante & Galetto 2008).
Dadas las características del sistema reproductivo de esta especie y la baja frecuencia de visitas que recibe, resulta muy interesante conocer las características de la secreción del néctar y la composición de azúcares para relacionarlas con la actividad de los polinizadores. Es decir, falta aún mucho por conocer en las interacciones planta-polinizador en general, y dentro de las Fabaceae en particular, para poder establecer algunos patrones entre la fisiología de la secreción del néctar, el sistema reproductivo de las plantas y el comportamiento de los polinizadores que permitan vislumbrar más claramente los posibles mecanismos que regulan las estrategias relacionadas a la optimización de los recursos tanto en plantas como en los animales polinizadores. El objetivo general de este trabajo fue estudiar las características del néctar de C. broussonetii y su relación con la frecuencia de visita de los polinizadores. Específicamente nos propusimos: (a) determinar la composición de azúcares del néctar, (b) establecer la tasa de producción de néctar desde la antesis hasta la senescencia floral y la respuesta de la flor ante extracciones sucesivas de néctar, (c) comparar la actividad de los visitantes florales según la cantidad de flores disponibles y según el momento del día, considerando las características de la fisiología de secreción del néctar a lo largo de la vida de la flor, y (d) estimar el número de granos de polen que depositan los polinizadores en el estigma.
MÉTODOS
Área de estudio
El trabajo de campo se realizó en una comunidad de bosque Chaqueño Serrano, localizada al pie de las Sierras Chicas (faldeo Oriental) en la Reserva Hídrica y Natural la Quebrada, (31°09' S, 64°20' W), provincia de Córdoba, Argentina. La región pertenece al piso de bosque Serrano, distrito Chaqueño Serrano, provincia fitogeográfica Chaqueña (Cabrera 1976). Se caracteriza por comunidades boscosas con un estrato arbóreo generalmente abierto (Luti et al. 1979). La vegetación natural es en general xerofítica.
En el área de estudio, C. broussonetii florece desde comienzos de febrero hasta fines de marzo. En general, los individuos de una misma población florecen de manera sincrónica. Los muestreos se realizaron sobre una misma población, entre el 21 de febrero y el 31 de marzo de 2001. Los ejemplares de referencia se depositaron en el Museo Botánico de Córdoba (CORD, Galetto 889).
Composición de azúcares
Se extrajo el néctar de cinco flores, mediante capilares de vidrio, con especial cuidado de no dañar el nectario o contaminar el néctar con polen. Se conservaron entre 2 y 3 µl de néctar a -18 °C, que se transportaron al laboratorio donde se realizó la identificación y cuantificación de carbohidratos por cromatografía gaseosa. La extracción de las muestras se realizó por medio de trimetilclorosilano + hexametildisilasano en piridina (Sweeley et al. 1963). Los extractos fueron inyectados en un cromatógrafo gaseoso Konik 3000-HRGS equipado con un integrador Spectra - Physics SP4290, un detector de ionización de llama y una columna OV 101 (2 m de longitud), 3% sobre Chromosorb G/AW - DMCS (malla 100-120). Se utilizó nitrógeno como gas portador (30 ml/ minuto) y el siguiente programa de temperatura: temperatura inicial 208 °C/ 2 minutos; luego, 1 °C/minuto hasta los 215 °C, temperatura que se mantuvo durante 1 minuto; por último, 8 °C/minuto hasta los 280 °C, en donde se estabilizó durante 5 minutos. Los azúcares testigo (Sigma Chemical Co.) fueron procesados del mismo modo.
La relación de azúcares se estimó como: r= sacarosa/(fructosa+glucosa) (Baker & Baker 1983). Antes de evaluar la tasa y producción total de néctar fue necesario determinar la longevidad floral; para ello se marcaron botones florales y se observaron desde la antesis hasta la senescencia.
Producción de néctar y tasa de producción
La producción de néctar durante la vida de la flor se evaluó mediante el siguiente procedimiento: en siete plantas se marcaron cuatro botones florales de la misma edad, las ramas floríferas se cubrieron con bolsas de tul para evitar la visita de polinizadores y permitir así la acumulación de néctar. Los botones florales se dividieron en cuatro grupos (un botón floral de cada una de las siete plantas conforman un grupo; es decir, se contó con cuatro grupos de siete botones cada uno). Cada grupo representa una edad en la vida de la flor, por lo que las extracciones de néctar se repartieron durante el tiempo que vive una flor (a las 14, 24, 38 y 48 h luego de producida la apertura floral). Se midió una vez a todas las flores de cada grupo: el volumen del néctar (µl), utilizando micropipetas graduadas y la concentración de azúcares (% sacarosa p/p), mediante un refractómetro de bolsillo Atago (rango: 0-32%). La cantidad de azúcar promedio producido por flor se expresó en miligramos (Galetto & Bernadello 2005).
La tasa de producción de néctar por hora (TPNh) se calculó como mg de azúcar producidos entre dos mediciones/número de horas transcurridas entre ellas (mg/h), mientras que la tasa de reabsorción de néctar por hora (TRNh) se estimó a través de la siguiente relación: mg de azúcar reabsorbidos entre dos mediciones/número de horas transcurridas (mg/h) (Galetto & Bernardello 2005).
Efecto de las remociones
Para medir el efecto de las remociones de néctar sobre la producción total se realizaron extracciones sucesivas a los cuatro grupos de flores mencionados anteriormente. Al grupo 1 se le extrajo néctar en cuatro oportunidades, a las 8:30 y 18:30 h de cada uno de los dos días que duró el tratamiento (tiempo que coincide con la longevidad floral); a las flores del grupo 2 se les extrajo néctar por primera vez a las 18:30 h del primer día, y a las 8:30 y 18: 30 h del segundo día (un total de tres remociones); el néctar de las flores del grupo 3 se dejó acumular durante el primer día y se extrajo a las 8: 30 y a las 18:30 h del segundo día (dos remociones). El grupo 4 fue utilizado como control, por lo que se dejó acumular el néctar durante el tiempo de vida de la flor y se realizó una única extracción a las 18:30 h del segundo día. En cada ocasión se midió el volumen (µl) y la concentración del néctar (% sacarosa p/p).
Visitantes florales
Las observaciones se llevaron a cabo sobre ocho parcelas de 80x80 cm de plantas de C. broussonetii, por lapsos de 10 minutos, durante distintos momentos del día (de 9 a 19 h) y a lo largo de la temporada de floración. Cada parcela se encontraba separada de las demás por al menos diez metros. Las parcelas fueron observadas una vez por día y cada día en un momento diferente (es decir, no se observó una misma parcela dos veces en el mismo horario). El tiempo total de observación fue de 510 minutos, distribuidos en 13 días.
Durante las observaciones se registró: el número de flores abiertas, el número de flores visitadas, los animales visitantes y la duración de cada visita. Se estimó: el tiempo medio de visita por flor=número de flores visitadas/tiempo que duró la vista; la tasa de visitas=(número de flores visitadas/número de flores observadas)/tiempo total de observación; y el porcentaje de flores visitado por unidad de observación (parcela)=(número de flores visitadas/número de flores observadas) x 100. Para evaluar si la actividad de los insectos era afectada por la disponibilidad de flores abiertas se comparó la actividad de los polinizadores entre parcelas con más de 20 flores abiertas (grupos grandes) y parcelas con menos de 20 flores abiertas (grupos pequeños). A fin de comparar la actividad de los insectos a lo largo del día, las observaciones se distribuyeron en tres franjas horarias; de 8:00 a 12: 00; de 12:00 a 15:00 y de 15:00 a 19:00 h.
Se estimó el número de granos de polen depositados en el estigma por los visitantes florales comparando el número de granos de polen presentes en estilos de flores senescentes que permanecieron expuestas a los polinizadores durante toda la vida de la flor, con el número de granos encontrados en estilos de flores que permanecieron excluidas de los animales visitantes durante toda su vida. En el campo, los pistilos fueron identificados y sujetados con cinta adhesiva a portaobjetos. Posteriormente, en el laboratorio, se tiñeron con azul de algodón y se llevó a cabo el recuento total de los granos de polen bajo el microscopio. Se calculó el número de granos de polen depositados por los visitantes florales como la diferencia entre el número de granos de polen encontrados sobre el estigma de flores expuestas a los polinizadores - el número de granos de polen depositados en el estigma de flores que permanecieron embolsadas.
Análisis de datos
Para el análisis de datos se utilizó el paquete estadístico SPSS (versión 10.0, 1999) siguiendo los métodos descriptos por Sokal & Rohlf (1995). La normalidad se evaluó mediante la prueba de Kolmogorov-Smirnov, mientras que la homogeneidad de varianza se analizó por intermedio de la prueba de Levene. Las variables que cumplieron los supuestos (transformadas o no) fueron analizadas utilizando estadística paramétrica (prueba t, MANOVA y ANOVA), mientras que las que no cumplieron los supuestos fueron analizadas con estadística no paramétrica (Mann-Whitney). Para comparar el efecto de las remociones de néctar sobre la producción total se utilizó la prueba t para la igualdad de medias. Se realizaron análisis multivariados (MANOVAs) para comparar las variables relacionadas con la actividad de los insectos (número de flores visitadas, tiempo medio de visita por flor y porcentaje de flores visitadas por parcela) en relación con el número de flores disponibles y con el momento del día. El porcentaje de flores visitadas por parcela fue transformado a arcoseno de la raíz cuadrada. Las diferencias significativas entre los tratamientos se evaluaron por la prueba 'a posteriori' de Bonferroni. Se utilizó análisis de varianza de un factor (ANOVA) para comparar el comportamiento entre las especies de Bombus; se tuvieron en cuenta distintas variables relacionadas con la polinización (número de flores visitadas, tiempo medio de visita por flor y porcentaje de flores visitadas por planta). Para comparar el número de granos de polen depositados en el estigma entre flores expuestas a los agentes polinizadores y flores que permanecieron aisladas se utilizó la prueba de Mann-Whitney. En el texto y en las tablas se presentan siempre los valores de las distintas variables en la unidad de medida en la que fueron tomadas, a fin de facilitar la lectura y comprensión de los resultados.
RESULTADOS
Composición de azúcares
La población estudiada de Cologania broussonetii produjo néctar dominante en sacarosa (r=1.63; según categorización propuesta por Baker & Baker 1990). Los porcentajes relativos promedio de cada uno de los azúcares del néctar fueron los siguientes: 62, 24 y 14% para sacarosa, fructosa y glucosa, respectivamente (n=5 individuos).
Producción de néctar y tasa de producción
Las flores de C. broussonetii se abrieron al atardecer y llegaron a la senescencia 48 h más tarde. El volumen total promedio producido por flor fue de 3.25 ± 2.25 µl. La secreción de néctar ocurrió durante el primer día de vida de la flor (aunque en forma lenta, ver TNPh más adelante), luego se detuvo y, antes de la senescencia, la flor reabsorbió el néctar que no fue extraído (Tabla 1, datos sobre la diagonal). La concentración del néctar aumentó durante el primer día, para luego comenzar a disminuir (Tabla 1, datos sobre la diagonal). La tasa de secreción fue relativamente constante (TNPh=0.021 mg/h, valor obtenido considerando las primeras mediciones de los grupos 1 y 2). La reabsorción activa del néctar se produjo durante el segundo día de vida de la flor a una tasa similar a la de la secreción activa [TNRh=-0.015 mg/h, valor obtenido tomando en cuenta las primeras mediciones de los grupos 2 y 4, (ver sección 'Métodos')].
Tabla 1. Volumen (µl), concentración (% p/p) y miligramos (mg) de azúcar del néctar de Cologania broussonetii, medidos en grupos de flores sometidos a distintos números de remociones. Los datos sobre la diagonal, primera medición realizada a cada grupo, (subrayados) corresponden a la dinámica de producción de néctar. Nota: valores expresados en promedio ± desvío estándar. Número de flores: 49.
Table 1. Nectar volume (µl), concentration (% wt/wt) and mass (mg) of sugar of Cologania broussonetii, measured in flower sets subjected to different removal schedules. Data on the diagonal (first measurement of each set underlined) correspond to the nectar production dynamics. Note: values expressed in mean ± standard deviation. Number of flowers: 49.

Efecto de las remociones de néctar
Los miligramos de azúcar producidos por flores sometidas a distinto número de remociones de néctar fueron, para el tratamiento uno (dos remociones) 0.69 ± 0.32 mg de azúcar y para el tratamiento dos (ahora control) 0.74 ± 0.32 mg de azúcar. Los tratamientos tres y cuatro no fueron considerados ya que para el momento en que se realizaron estas extracciones el néctar se estaba reabsorbiendo. No se encontraron diferencias significativas entre los dos grupos (prueba t; P=0.45), lo que indica que la extracción de néctar durante el periodo de secreción activa no afectó el ritmo natural de producción (Tabla 1).
Visitantes Florales
Al comparar, en forma conjunta, las variables consideradas en la actividad de los insectos (número de flores visitadas, tiempo medio de visita por flor y porcentaje de flores visitadas por parcela) en relación con el número de flores disponibles (grupos grandes: >20 flores abiertas; grupos pequeños: <20 flores abiertas), se encontraron diferencias marginalmente significativas (MANOVA; P=0.06). Luego de realizar un análisis 'a posteriori' se encontró que el porcentaje de flores visitadas por planta fue significativamente mayor cuando las flores se presentaron en grupos pequeños (Prueba de Bonferroni; P=0.008; Tabla 2). El número de flores visitadas y el tiempo medio de visita por flor no presentaron diferencias significativas entre los grupos grandes (>20 flores abiertas) y los grupos pequeños (<20 flores abiertas) (prueba de Bonferroni; P=0.76 y P=0.30, respectivamente).
Tabla 2. Actividad de los visitantes florales según el número de flores abiertas por sitio. Abreviaturas: n=número de parcelas observadas; n*=número de parcelas que recibieron visitas; (s)=segundos; DE=desvío estándar; Mín=mínimo; Máx=máximo.
Table 2. Activity of the flower visitors according to the number of open flowers per site. Abbreviations: n=number of sites observed; n*=number of sites where bees have visited flowers; (s)=seconds; DE=standard deviation; Mín=minimum; Máx=maximum.
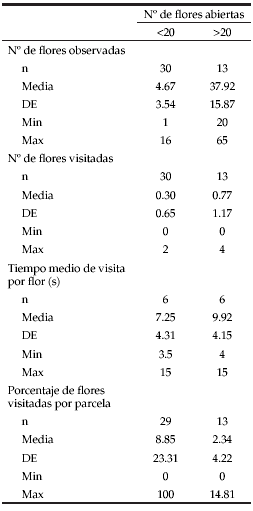
A lo largo del día, el número y porcentaje de flores visitadas disminuyó mientras que el tiempo medio de visita por flor aumentó (Tabla 3). Sin embargo, esas diferencias no resultaron estadísticamente significativas para ninguna de las variables analizadas (MANOVA; P=0.76). Por otro lado, cabe destacar que los datos disponibles sobre el comportamiento de los polinizadores fueron poco numerosos, a pesar de las horas de observación realizadas, que solo permitieron registrar 58 individuos de B. opifex y 18 B. bellicosus que no interactuaron con las flores de C. broussonetii y que atravesaron las parcelas de observación o interactuaron con otras especies en flor. Durante los días nublados o muy ventosos no se observaron visitas de los insectos a las flores.
Tabla 3. Actividad de los visitantes florales de C. broussonetii durante distintos momentos del día. Abreviaturas: (s)=segundos; n=número de parcelas observadas; n*=número de insectos observados por franja horaria; DE=desvío estándar.
Table 3. Activity of the flower visitors of C. broussonetii during different moments of the day. Abbreviations: (s)=seconds; n=number of sites observed; n*=number of insects; DE=standard deviation.

El número de granos de polen depositados en el estigma de las flores expuestas a los agentes polinizadores y en los de las flores que permanecieron embolsadas durante toda la antesis resultó similar (20 ± 18; n=33 flores vs. 22 ± 33; n=55 flores, respectivamente; prueba de Mann-Whitney; P=0.14).
DISCUSIÓN
La alta proporción de sacarosa observada en el néctar floral de C. broussonetii es habitual entre las Fabáceas (Percival 1961) y coincide con la más frecuente que encuentran Baker & Baker (1983) para flores habitualmente visitadas por abejorros de trompa larga (gremio al que pertenecen los abejorros B. bellicosus y B. opifex). Sin embargo, muchas de las Fabáceas nativas de Argentina, estudiadas hasta el momento, poseen un néctar dominante en monosacáridos (Cocucci et al. 1992; Forcone et al. 1997; Galetto & Bernadello 2003). La concentración de azúcares presentes en el néctar se ha relacionado con los requerimientos energéticos que poseen los visitantes florales. Baker & Baker (1983) resumieron los resultados obtenidos por diferentes autores en distintos lugares del mundo, correspondiendo para las abejas de trompa larga valores de la concentración del néctar (% p/p) que oscilan entre 24 y 46%. En la población estudiada se obtuvieron concentraciones promedio que variaron entre 18 y 32%. Asimismo, en esta población se observó un volumen de néctar menor al documentado por Opler (1983) como preferido por los abejorros (media=9.75 µl). Resulta evidente la necesidad de profundizar los estudios sobre el néctar en especies de Fabáceas polinizadas por abejorros a fin de clarificar los patrones y determinar si éstos se relacionan más con un componente filogenético o bien con las interacciones planta-polinizador.
Las flores de C. broussonetii secretaron néctar solo durante el primer día de vida de la flor para luego comenzar a reabsorberlo activamente durante el segundo día. Este patrón de secreción contrasta con lo observado para otras especies argentinas de Fabáceas donde la producción de néctar es continua durante toda la vida de la flor y el néctar no es reabsorbido (Cocucci et al. 1992; Galetto et al. 2000). El patrón de C. broussonetii contrasta con el de otras Fabaceae de Argentina, y las evidencias disponibles para otras regiones del mundo no presentan una consistencia tal que permitan una comparación más amplia.
Las extracciones sucesivas de néctar a las flores pueden tener distintos efectos según las especies. Por ejemplo, se ha observado que en algunas especies se puede modificar el patrón de secreción de néctar ya sea a través del incremento o de la disminución de la producción total (e.g., Galetto & Bernardello 1992, 1993, 1995, 2004), ó bien la cantidad de néctar secretada por flor puede no ser afectada (e.g., Galetto & Bernardello 1992, 1993, 2004). Los resultados obtenidos para esta especie indican que la producción de néctar no es afectada por su remoción (i.e., visita de los polinizadores), ya que no se modifica la tasa de producción ni la cantidad total de azúcares producida. Se ha comprobado que la facultad de reabsorber el néctar puede relacionarse con la reasignación de recursos para madurar semillas (Pyke 1991). Si se tiene en cuenta el bajo número de visitas que recibiría en promedio cada flor de C. broussonetii, que el patrón de secreción luego de una visita no se modifica y que las flores reabsorben el néctar que no fue consumido por los polinizadores, existe la posibilidad que estas particularidades de la fisiología de secreción del néctar le permitan a esta especie ahorrar recursos energéticos y destinarlos a otras funciones de la planta, como la maduración de frutos y semillas. Sin embargo, esta posibilidad debería ser probada experimentalmente.
La población estudiada de C. broussonetii fue visitada legítimamente solo por abejorros del género Bombus, entonces, se podría considerar a esta planta como especializada en su relación con los visitantes florales. Sin embargo, esta interacción de polinización resulta asimétrica por cuanto estos visitantes florales son generalistas, ya que necesitan un amplio rango de plantas del que alimentarse durante la temporada (Arroyo 1981; observaciones personales). Futuras investigaciones deberían orientarse a ampliar la evidencia y, eventualmente, confirmar el grado de especialización de esta interacción. La especialización en las flores papilonadas de Fabaceae se manifiesta cuando se considera una clara tendencia a maximizar la economía del polen y néctar, su frecuente interacción con polinizadores especializados y por la presencia de estructuras que permiten el ocultamiento de las recompensas (Arroyo 1981).
El patrón de visitas obtenido no fue coherente con la hipótesis de atracción que postula un número creciente de visitas de polinizadores al incrementarse la señal de atracción (Thompson 1981). Es decir, las parcelas con grupos grandes de flores (>20 flores abiertas) no resultaron más atractivas para los polinizadores que las parcelas que contenían menos flores disponibles. Por otra parte, la proporción de flores visitadas por polinizador fue mayor en las parcelas con pocas flores, lo que provocaría una mayor geitonogamia en estos grupos pequeños de flores. En el caso particular de C. broussonetii, el polen presente en los estigmas puede ser producto de las visitas de los polinizadores o bien de la autodeposición espontánea dentro de la misma flor (Musicante & Galetto 2008). Al ser una especie autocompatible, la producción de frutos y semillas es asegurada de una u otra manera. Sin embargo y pese a la baja frecuencia de visitas y a la inexistente diferencia entre el número de granos de polen depositados en el estigma de flores expuestas y aisladas de los polinizadores, se observó una mayor cantidad y calidad de los frutos y semillas (mayor éxito reproductivo y mayor masa y germinabilidad de las semillas) producidos por polinización libre (Musicante & Galetto 2008), lo cual estaría indicando que la relación planta-polinizador aporta un beneficio reproductivo a C. broussonetii.
AGRADECIMIENTOS
A L. Ashworth y a dos revisores anónimos que con sus comentarios y sugerencias mejoraron versiones anteriores de este trabajo. Este trabajo fue posible gracias al financiamiento que aportaron SECyT (UNC), FONCyT y CONICET. MM es becaria de CONICET y LG miembro de la Carrera del Investigador de la misma institución. MM es alumna de la Carrera del Doctorado en Ciencias Biológicas de la FCEFyN-UNC.
BIBLIOGRAFÍA
ARROYO, MTK. 1981. Breeding systems and pollination biology in Leguminoseae. Pp. 723-769 en: Polhill, RM & PH Raven (Eds.). Advances in Legume systematics. Part. 2. Royal Botanical Gardens. Kew. [ Links ]
BAKER, HG & I BAKER. 1983. A brief historical review of the chemistry of floral nectar. Pp. 126- 152. en: Bentley, B & TS Elias (Eds.). The Biology of nectarines. Columbia University Press. New York. [ Links ]
BAKER, HG & I BAKER 1990. The predictive value of nectar chemistry to the recognition of pollinators types. J. of Bot. (Israel) 39(1-2):157-166. [ Links ]
BERNARDELLO, LM; L GALETTO & IG RODRÍGUEZ. 1994. Reproductive biology, variability of nectar features and pollination of Combretum fruticosum (Combretaceae) in Argentina. Bot. J. Linn. Soc. 114:293-308. [ Links ]
BURKART, A. 1952. Las leguminosas argentinas silvestres y cultivadas. Pp. 385-386. 2da ed. Editorial ACME. Buenos. Aires. 590 pp. [ Links ]
CABRERA, AL. 1976. Regiones Fitogeográficas argentinas. Pp. 1-85 en: Kugler, WF (Ed.). Enciclopedia argentina de agricultura y jardinería. Tomo 2. Edit. ACME. Buenos Aires. [ Links ]
COCUCCI, AA; L GALETTO & A SERSIC. 1992. El sindrome floral de Caesalpinia gilliesii (Fabaceae- Caesalpinioideae). Darwiniana 31(1-4):111-135. [ Links ]
DAFNI, A. 1992. Pollination ecology. A practical approach. Pp. 165-201 en: Rickwood, D & BD Hames (Eds.) The practical approach series. Oxford University Press. London. 272 pp. [ Links ]
FAEGRI, K & L VAN DER PIJL. 1971. The principles of pollination ecology. 2da ed. Pergamon Press, Oxford. 291 pp. [ Links ]
FORCONE, AL; GALETTO & L BERNARDELLO.1997. Floral nectar chemical composition of some species from Patagonia. Biochem. Syst. and Ecol. 25(5): 395-402. [ Links ]
GALETTO, L & LM BERNARDELLO. 1992. Nectar secretion pattern and removal effects in six Argentinean Pitcairnioideae (Bromeliaceae). Bot. Acta. 105(4):292-299. [ Links ]
GALETTO, L & LM BERNARDELLO. 1993. Nectar secretion pattern and removal effects in three species of Solanaceae. Can. J. Bot. 71(10):1394- 1398. [ Links ]
GALETTO, L & LM BERNARDELLO. 1995. Characteristics of nectar secretion by Lycium cestroides, L. ciliatum (Solanaceae) and their hybrid. Plant Species Biol. 11(2-3):157-163. [ Links ]
GALETTO, L & G BERNARDELLO. 2003. Nectar sugar composition in angiosperms from Chaco and Patagonia (Argentina): an animal visitor's matter? Plant. Syst. and Evolut. 238(1-4):69-86. [ Links ]
GALETTO, L & G BERNARDELLO. 2004. Floral nectarines, nectar production dynamics and chemical composition in six Ipomoea species (Convolvulaceae) in relation to pollinators. Ann. of Bot. 94:269-280. [ Links ]
GALETTO, L & G BERNARDELLO. 2005. Nectar. Pp. 261- 313 en: Dafni, A; PG Kevan & BC Husband (Eds.). Rewards in flowers. Practical Pollination Biology. Cambridge, Ontario. [ Links ]
GALETTO, L; LM BERNARDELLO & HR JULIANI. 1994. Estructura del nectario, composición química del néctar y mecanismo de polinización en tres especies de Siphocampylus (Campanulaceae). Kurtziana 22:81-96. [ Links ]
GALETTO, L; G BERNARDELLO; I ISELE; JL VESPRINI; G SPERONI; ET AL. 2000. Reproductive biology of Erythrina cristagalli (Fabaceae). Ann. of the Missouri Bot. Garden 87:127-145. [ Links ]
HERRERA, CM. 1987. Components of pollinator "quality": comparative analysis of a diverse insect assemblage. Oikos 50(1):79-90. [ Links ]
LUTI, RF; M GALERA & M BERTRAÁN DE SOLIS. 1979. Vegetación. Pp. 125-367 en: Vázquez, JB (Ed.). Geografía física de la provincia de Córdoba. Editorial Boldt. Banco Provincia de Córdoba. Córdoba. [ Links ]
MUSICANTE, ML & L GALETTO. En prensa. Biología reproductiva de Cologania broussonetii (Fabaceae). Darwiniana. [ Links ]
OPLER, PA. 1983. Nectar production in a tropical ecosystem. Pp. 30-79 en: Bentley, B & TS Elias (Eds.) The biology of nectaries. Columbia University Press. New York. [ Links ]
PERCIVAL, M. 1961. Types of nectars in angiosperms. New Phytol 60(3):235-281. [ Links ]
PYKE, GH. 1991. What does it cost a plant to produce floral nectar? Nature 350:58-59. [ Links ]
REAL, LA & BJ RATHCKE. 1988. Patterns of individual variability in floral resources. Ecol. 69(3): 728-735. [ Links ]
SIMPSON, BB & JL NEFF. 1983. Evolution and diversity of floral rewards. Pp. 142-159 en: Jones, CE & RJ Little (Eds.). Handbook of experimental pollination biology. New York. [ Links ]
SWEELEY, SC; R BENTLEY; M MAKITA & WW WELLS. 1963. Gas liquid chromatography of trimethylsilyl derivatives of sugar and related substances. J. of Amer. Chem. Soc. 85:2497-2507. [ Links ]
SOKAL, RR & FJ ROHLF. 1995. Biometry: the principles and practice of statistics in biological research. 3a ed. WH Freeman & Co., New York. 887 pp. [ Links ]
THOMPSON, JD. 1981. Spatial and temporal components of resource assessment by flower-feeding insects. J. Anim. Ecol. 50(1): 49- 59. [ Links ]
TORRES, C & L GALETTO. 1998. Patterns and implications of floral nectar secretion, chemical composition, removal effects, and standing crop in Mandevilla pentlandiana (Apocynaceae). Bot. J. of the Linnean Soc. 127(3):207-223. [ Links ]
WASER, NM. 1983. Competition for pollination and floral character differences among sympatric plant species. a review of evidence. Pp. 277-293 en: Jones, CE & RJ Little (Eds.). Handbook of experimental pollination biology. Van Nostrand Reinhold. New York. [ Links ]

