










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.18 n.3 Córdoba sept./dic. 2008
SECCIÓN ESPECIAL
Efecto de las terrazas sobre la diversidad de artrópodos caminadores en cultivos de soja
Federico Weyland1,* & María Elena Zaccagnini2
1. INTA EEA Parana, Oro Verde, Entre Rios, Argentina.
2. Instituto de Recursos Biológicos (IRB), INTA-CNIA, Castelar, Buenos Aires, Argentina.
* INTA EEA Parana, Ruta 11 km 12.5, Oro Verde, Entre Rios, Argentina. Dirección actual: IFEVA - Facultad de Agronomía, Universidad de Buenos Aires, Argentina. Email: fweyland@ifeva.edu.ar
Recibido: 23 de mayo de 2008;
Fin de arbitraje: 28 de julio de 2008;
Revisión recibida: 2 de septiembre de 2008;
Aceptado: 6 de octubre de 2008
RESUMEN. La pérdida de la biodiversidad y sus funciones en agroecosistemas es un tema de interés creciente en la investigación ecológica. Las terrazas, utilizadas para el control de la erosión hídrica, podrían actuar como refugios de artrópodos en los paisajes agrícolas. En este trabajo relevamos la riqueza y la abundancia total de especies y de depredadores en lotes de soja cultivados bajo el sistema de siembra directa y con la implementación o no de terrazas. Para el total del ciclo del cultivo, la riqueza total de especies fue significativamente mayor en los márgenes que en el cultivo, pero no así la de depredadores. La abundancia fue mayor en los márgenes que en los cultivos sólo para el total de individuos. Los depredadores fueron más abundantes en los márgenes sólo al comienzo y al final del ciclo del cultivo. Los lotes con terrazas no favorecieron la abundancia de artrópodos pero sí la riqueza tanto de depredadores como del total de especies, lo cual indica que la heterogeneidad ambiental que introducen las terrazas favorece la diversidad de artrópodos en la escala de lote. La riqueza de depredadores estuvo altamente correlacionada a la riqueza total de especies, lo que sugiere que este grupo funcional podría ser un buen indicador de la riqueza de especies estimada por trampas de caída.
Palabras clave: Biodiversidad; Depredadores; Riqueza de especies; Control de la erosión; Agroecosistemas; Siembra directa.
ABSTRACT. Effect of terraces on soil dwelling arthropod diversity in soybean production: The loss of biodiversity in agroecosystems constitutes a main issue in ecological research. Steep backslope terraces, used for soil erosion control, could act as refuges for arthropods in agricultural landscapes. Total and predator arthropod species richness and abundance were surveyed in soybean fields in Entre Ríos under a no-till system with and without terraces using pit-fall traps. For the total crop period, total species richness was significantly higher at the margins than in the crop, but not for predator species. Abundance was higher in margins than in crops only when total individuals were considered. Predators were more abundant in margins only at the beginning and the end of the crop cycle, although, in fields with terraces abundance and richness were never higher than in fields without terraces for both total species and predators. Field with terraces did not increase the abundance but increased the species numbers, both total and predators, showing that habitat heterogeneity introduced by terraces favors arthropod diversity at the field scale.
Keywords: Biodiversity; Predators; Species richness; Erosion control; Agroecosystems; No-tillage.
INTRODUCCIÓN
La conservación de la biodiversidad y de sus funciones ha sido uno de los temas centrales de la Ecología en los últimos años (Altieri 1999; Chapin III et al. 2000). Las especies proveen bienes y servicios a la agricultura y a otras actividades humanas, y la diversidad resulta una manera de mantener el funcionamiento de los ecosistemas y su resiliencia (Pimentel et al. 1997; Swift et al. 2004). Por lo general, ciertos factores asociados con las actividades productivas del hombre amenazan estas funciones (e.g., el cambio en el uso de la tierra). Según las proyecciones de Chapin III et al. (2000), para el año 2100 este factor será el de mayor impacto sobre la biodiversidad. Por esta razón, las investigaciones han sido orientadas al estudio de aquellos elementos y manejos del paisaje que pueden favorecer la biodiversidad.
En paisajes agrícolas, los márgenes de los lotes son considerados reservorios de biodiversidad ya que sustentan poblaciones de artrópodos que, de otra manera, no podrían sobrevivir en campos cultivados (Dennis & Fry 1992; de Snoo 1999). Estudios empíricos han demostrado que los márgenes sustentan poblaciones de artrópodos al ofrecer fuentes alternativas de alimento, protección frente a pesticidas y refugios para la sobreinvernación (Dennis & Fry 1992; Thomas & Marshall 1998; Thomas et al. 2002).
La construcción de "terrazas" siguiendo curvas de nivel constituye una técnica de control de la erosión hídrica muy difundida en la provincia de Entre Ríos, Argentina. Sus efectos sobre la biodiversidad podrían resultar similares a los de los márgenes de cultivo debido a sus características estructurales y de manejo. Las terrazas consisten en una loma que retiene el agua de escorrentía y la conduce fuera del lote a través de un canal. Como en estas lomas no se realizan cultivos, se desarrolla una franja de entre cuatro y cinco metros de vegetación espontánea. El conocimiento del efecto de estas terrazas sobre la biodiversidad todavía es escaso. Si bien estudios previos muestran una asociación positiva entre la presencia de terrazas y la diversidad de aves, no sucede lo mismo con los anfibios y los mamíferos (Zaccagnini et al. 2007). Hasta donde sabemos, no existen estudios que exploren la influencia de las terrazas sobre las comunidades de artrópodos, aunque sí se ha estudiado la influencia de los márgenes de los cultivos. Liljesthröm et al. (2002), por ejemplo, encontraron que en la región pampeana la diversidad de arañas era mayor en los bordes naturales que en los cultivos de soja. En el caso de que las terrazas tuvieran una influencia positiva sobre la biodiversidad, es posible que el incentivo para implementarlas fuera mayor. Esto resultaría particularmente importante en aquellas áreas con pendientes considerables. Por lo tanto, creemos que es de fundamental importancia estudiar y caracterizar la diversidad de artrópodos en las terrazas, de acuerdo con sus roles funcionales vinculados con la agricultura.
Existe una cantidad de especies que podrían funcionar como indicadoras en el monitoreo de la diversidad de artrópodos (Oliver & Beattie 1996). La familia Carabidae (Coleoptera) es uno de los indicadores que más se utilizan por la facilidad para identificar las especies que la integran, y porque estas especies desempeñan un papel muy importante en el control de plagas en los cultivos (Lövei & Sunderland 1996; Pearce & Venier 2006). También se han propuesto otros grupos de artrópodos, como mariposas (Mac Nally & Fleishman 2004; Fleishman et al. 2005), ortópteros (Kati et al. 2004), abejas (Oertli et al. 2005) y arañas (Billeter et al. 2008). Los resultados son variables, y hasta ahora no se han encontrado grupos que muestren correlaciones fuertes con la diversidad total. Esto sugiere la necesidad de continuar explorando otros grupos que permitan el monitoreo de la biodiversidad en los agroecosistemas. En este trabajo, propusimos el grupo de los depredadores caminadores dado que cumplen un papel importante como controladores de plagas en los agroecosistemas. Intentamos responder las siguientes preguntas: 1) ¿Tienen las terrazas el mismo efecto que los márgenes de cultivo en la riqueza y abundancia de artrópodos? 2) ¿Pueden las terrazas y los márgenes sustentar mayor abundancia y riqueza de artrópodos que los cultivos de soja? 3) ¿Pueden los cultivos con terrazas sustentar una mayor riqueza y abundancia de artrópodos que los cultivos sin terrazas? 4) ¿Tienen los depredadores el mismo patrón de riqueza y abundancia que el total de artrópodos?
MÉTODOS
El área de estudio estuvo ubicada en la subregión del ñandubay, en la ecoregión del Chaco-Espinal (Burkart et al. 1999), un territorio que pertenece a la provincia de Entre Ríos (Figura 1). La subregión del ñandubay se caracteriza por poseer un clima templado y relativamente húmedo, con una temperatura media anual de 18.5 ºC y una precipitación media anual de 995 mm. La vegetación es de tipo xerofítica, dominada por especies leñosas como el ñandubay (Prosopis affinis), el algarrobo (Prosopis nigra), el molle (Schinus longifolia) y el espinillo (Acacia caven) (Burkart et al. 1999). Esta área se encuentra actualmente bajo un uso agrícola intenso, y los cultivos predominantes son la soja, el trigo y el maíz.

Figura 1. Lotes muestreados. Los polígonos en tonalidad gris oscuro representan lotes con terrazas, y los polígonos en tonalidad gris claro, lotes sin terrazas. El diseño de muestreo es ejemplificado en un lote con terrazas y otro sin terrazas. Los puntos representan sitios de muestreo con dos trampas de caída en cada uno.
Figure 1. Fields surveyed. Dark grey polygons represent fields with terraces and light grey polygones fields without terraces. Sampling design is exemplified with a field with terraces and a field without terraces. Points represent sampling points with two pit-fall traps in each one.
Tomamos muestras en seis lotes de soja durante la campaña agrícola 2003-2004. Los lotes se encontraban bajo un manejo de siembra directa, técnica que prescinde del uso del arado y que realiza la siembra sobre los restos del cultivo anterior (rastrojos). Los lotes estaban localizados en El Palenque, Entre Ríos, Argentina (31º40´ S; 60º11´ W), y la mitad de ellos tenía terrazas para control de la erosión. El tamaño promedio de los lotes fue de 11.5 ha (mín.: 4.7 ha; máx.: 35 ha). Los lotes estaban ubicados relativamente cerca uno del otro (<500 m). Las terrazas tenían una vegetación leñosa bien desarrollada y una gran cobertura del estrato herbáceo, compuesto por especies del género Carduus, Conyza y Cynodon. La vegetación arbórea de los márgenes de los lotes sin terrazas estaba menos desarrollada y la cobertura era principalmente de herbáceas (Stipa sp., Solanum sisybrifolium, Cynodon dactylon, Carduus sp.).
En los lotes con terrazas se seleccionaron al azar dos terrazas, y en cada una de ellas ubicamos tres transecciones también al azar. La primera transección fue establecida a una distancia al azar desde el borde del lote, mientras que las otras dos fueron ubicadas a 30 m de distancia una de la otra (Figura 1). Se establecieron dos puntos de muestreo a lo largo de la transección, uno dentro de la terraza y el otro a una distancia de 20 m dentro del cultivo. En cada punto se ubicaron dos trampas de caída (pit-fall), separadas 2 m entre sí. En los lotes sin terrazas, ubicamos las transecciones desde los márgenes hacia el cultivo bajo el mismo esquema (Figura 1). En total fueron colocadas 144 trampas. Se tomaron muestras en cinco ocasiones desde enero de 2004 (soja en estadio de desarrollo V2-V3) hasta abril 2004 (cosecha del cultivo), una vez cada 3 ó 4 semanas. Las trampas estuvieron activas por cinco días consecutivos durante cada período de muestreo. Entre los muestreos 3 y 4 (fines de febrero) se realizaron aplicaciones de insecticidas en los lotes según el manejo habitual en la zona (Mancuso, comunicación personal). Las trampas consistieron en un vaso de plástico de 8 cm de diámetro, que contenía una solución conservante de formol al 2%, ácido acético al 2%, y una gota de detergente para disminuir la tensión superficial. Se colocó un techo de metal a 10 cm del suelo para proteger a la trampa de la lluvia y de la luz del sol (Lövei & Sunderland 1996; Duelli et al. 1999; Marc et al. 1999).
Los individuos recolectados fueron conservados en etanol al 70% hasta su identificación. En los casos en que no se pudo determinar la identidad taxonómica de los individuos, se utilizó un criterio de morfoespecies para calcular los valores de riqueza. Este criterio ha sido propuesto para esta clase de estudios debido a la dificultad de relevar la diversidad de artrópodos (Oliver & Beattie 1996; Duelli & Obrist 2003). Para determinar la función ecológica de cada especie se utilizaron los hábitos alimentarios definidos según la bibliografía: fitófagos, depredadores, parasitoides, detritívoros y micetófagos. A pesar de que tradicionalmente los isópodos son considerados detritívoros (Sousa et al. 1999) o coprófagos (Kautz et al. 2002), las especies en nuestra área de estudio atacan la soja en los estadios de desarrollo más tempranos del cultivo (Saluso 2004). Por esta razón, los isópodos fueron incluidos dentro de los fitófagos, aunque su rol puede variar según la disponibilidad de materia en descomposición. No contabilizamos los Acari, Collembola, Formicidae y Lepidoptera porque la metodología empleada es inadecuada para estimar correctamente su abundancia o riqueza específica.
Analizamos la riqueza específica y la abundancia de individuos, tanto para el total de artrópodos como para los depredadores solamente. Las variables fueron analizadas para cada fecha de muestreo con un análisis de varianza de dos vías con dos factores: prácticas de control de erosión (con y sin terrazas) y tipo de hábitat (margen y cultivo). Se realizaron contrastes de Tukey para comparaciones entre terrazas y márgenes (Sokal & Rohlf 1995). La riqueza fue estimada mediante curvas de acumulación de especies con el índice ACE-1 (Chao & Lee 1992) para tener en cuenta las especies no detectadas. Utilizamos el programa SPADE (Chao & Shen 2003). Los datos de todas las fechas de muestreo fueron combinados a fin de estimar el efecto de las terrazas sobre la diversidad de artrópodos durante todo el ciclo del cultivo. En los casos de ruptura de alguna de las trampas, las estimaciones de abundancia se hicieron multiplicando por dos el valor de la trampa remanente. A pesar de que este método tiene un error asociado, la corrección fue hecha sólo en 5% de los casos. La abundancia también fue analizada para cada fecha de muestreo por separado sin comparaciones entre distintas fechas, debido a que no se cumplían los supuestos de las pruebas de medidas repetidas.
Con el fin de explorar las posibles asociaciones entre los depredadores y el total de artrópodos realizamos regresiones lineales entre la riqueza de depredadores y la de artrópodos, y entre sus abundancias. Los datos fueron transformados logarítmicamente cuando fue necesario para alcanzar normalidad y homogeneidad de varianza. Se calculó la potencia de todas las pruebas estadísticas para estimar la probabilidad de cometer un error de tipo II.
RESULTADOS
Patrones generales de riqueza y abundancia
Se identificó un total de 375 morfoespecies y 39169 individuos. El orden mejor representado fue Coleoptera, con 45% del total de las especies. Dentro de este orden, 21 especies pertenecieron a la familia Carabidae (5.6% del total de las especies identificadas). Los siguientes grupos mejor representados fueron Araneae (16.3%), Heteroptera (13%) e Hymenoptera (8.8%) (ver Apéndice en www.ecologiaaustral.com.ar). Los depredadores, en su mayoría Coleoptera y Araneae, constituyeron 37.6% de todas las morfoespecies. El grupo funcional de los fitófagos fue el siguiente mejor representado (17.1% del total). Debido a la imposibilidad de alcanzar un nivel taxonómico adecuado, 34.4% de las especies no fue asignadas a ningún grupo funcional. El orden Isopoda fue el más abundante (55%), seguido por Coleoptera (21%), Araneae (7%) y Orthoptera (7%) (ver Apéndice). Los Carabidae representaron 5% del total de abundancia. Los restantes órdenes estuvieron mucho menos representados, probablemente debido a que las trampas de caída no resultaron eficientes para muestrear el estrato en el que se encuentran esos órdenes. Los fitófagos representaron 65% del total de abundancia (ver Apéndice), en gran medida por el número elevado de isópodos recolectados (81% de este grupo). Los depredadores constituyeron 23% del total de abundancia. Las especies detritívoras y coprófagas fueron menos abundantes.
Efecto de las terrazas
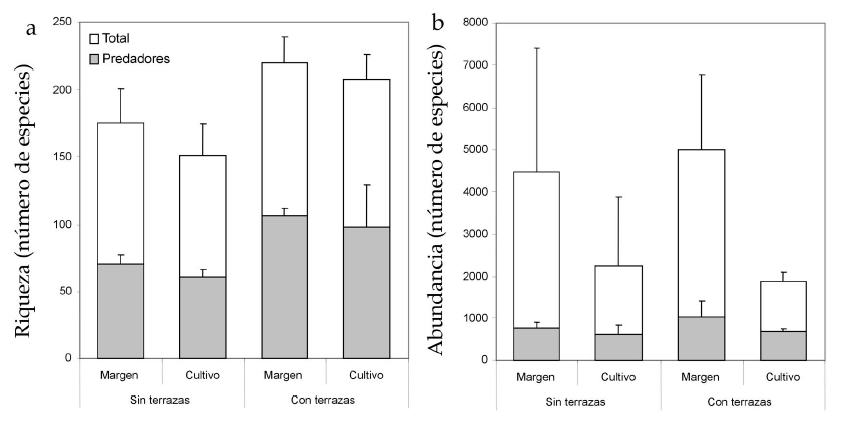
El control de la erosión por medio de terrazas no afectó la riqueza total de los márgenes (contraste de Tukey F1,4=2; P=0.23), mientras que aumentó la del cultivo (F1,8=11.01; P=0.009; Figura 2a). A su vez, el control de la erosión aumentó la riqueza de los depredadores, tanto en los márgenes (contraste de Tukey F1,4=47.31; P=0.002) como en el cultivo (F1,8=16.54; P=0.0028; Figura 2a). El control de la erosión no afectó la abundancia total de artrópodos ni al considerar el total del ciclo (F1,8=0.09; P=0.76; Figura 2b), ni al considerar cada fecha de muestreo por separado (Figura 3a). Tampoco afectó la abundancia de los depredadores cuando se analizó todo el ciclo del cultivo (F1,8=3.26; P=0.1; Figura 2b), pero el análisis de cada fecha por separado reveló una mayor abundancia en los cultivos con terrazas durante la primera fecha de muestreo (Figura 3b).
El tipo de hábitat afectó la riqueza total de artrópodos, que resultó mayor en los márgenes que en los cultivos (F1,8=5.52; P=0.04), mientras que no afectó la de los depredadores (F1,8=0.96; P=0.35; Figura 2a). Por su parte, el tipo de hábitat afectó la abundancia total de artrópodos, que fue más del doble en los márgenes que en los cultivos (F1,8=8.41; P=0.02; Figura 2b y Figura 3a), mientras que no afectó la de los depredadores al considerar el total del ciclo del cultivo (F1,8=3.26; P=0.1; Figura 2b). En cambio, sí se detectaron diferencias de abundancia de depredadores al analizar cada fecha por separado. En estos casos, los depredadores fueron más abundantes en los márgenes hacia el comienzo y el final del ciclo del cultivo (muestreo 1: F1,8=3.26; P=0.1; muestreo 5: F1,8=3.26; P=0.1; Figura 3b). Desde el comienzo de la floración hasta el estadio de llenado de grano, la abundancia de depredadores no difirió entre ambos tipos de hábitat. Las pruebas de potencia realizadas cuando no se detectaron diferencias significativas entre tratamientos mostraron una potencia muy baja en todos los casos (ninguna potencia fue mayor de 50%).
Figura 2. Valores promedio (+Desvío Estándar) de riqueza (panel a) y abundancia (panel b) del total de artrópodos y de depredadores. Datos combinados de cinco fechas de muestreo.
Figure 2. Total and predator Mean (+ S.D.) richness (A) and abundance (B). Pooled data of five sampling dates.

Figura 3. Valores promedio (± Desvío Estándar) de abundancia total (panel a) y de depredadores (panel b), discriminando cada fecha de muestreo (la fecha corresponde al incio del muestreo).
Figure 3. Total (a) and predator (b) mean (+S.D.) abundance in each sampling date (dates correspond to the beginning of the sampling period). *= significant for factor habitat type. **= significant for factor soil erosion control.
La riqueza de depredadores se correlacionó con el número total de especies (r =0.92; P<0.001), mientras que su abundancia estuvo menos correlacionada con la abundancia total (r=0.58; P=0.047). Por otro lado, no se encontraron correlaciones significativas entre los Carabidae y el total de artrópodos para abundancia (r=0.34; P=0.27) ni para riqueza específica (r=0.31; P=0.33).
DISCUSIÓN
Nuestros resultados muestran que el control de la erosión mediante terrazas favorece la riqueza de artrópodos en los cultivos, aunque no su abundancia. Los ambientes de margen presentaron mayor riqueza y abundancia de especies que los ambientes de cultivo, a excepción de los depredadores. Estos resultados tienen consecuencias importantes para el manejo de los lotes de la región, ya que indican que las terrazas benefician tanto el control de la erosión como la conservación de de la biodiversidad en agroecosistemas.
La mayor riqueza de especies en los lotes con terrazas que en los cultivos sin terrazas pudo deberse a que los cultivos con terrazas tenían una mayor relación borde/área. Esto proporcionaría una mayor diversidad de hábitat y favorecería la colonización del cultivo desde las terrazas. Incluso, la riqueza de depredadores en las terrazas fue mayor que en los márgenes de los lotes sin terraza, lo que demuestra el beneficio de estos ambientes como reservorio de controladores de plagas. Estos resultados coinciden con los de De la Peña et al. (2003), quienes encontraron que las especies de carábidos son más abundantes en sitios con mayor densidad de márgenes (i.e., mayor heterogeneidad). Irmler (2003), por otro lado, encontró una correlación negativa entre la riqueza de carábidos y el tamaño del lote, y argumentó que el largo de los márgenes podría ser una causa de este patrón. Por el contrario, Fahrig & Jonsen (1998) no encontraron una relación entre el tamaño de lotes de alfalfa y la riqueza de familias de insectos.
La mayor riqueza de especies que encontramos en los márgenes con respecto a los cultivos posiblemente se deba a la mayor diversidad vegetal de los ambientes de margen. Esta mayor diversidad podría ofrecer una mayor variedad de nichos ecológicos para distintas especies de artrópodos. Estos resultados coinciden con los de otros autores que encontraron una mayor riqueza de artrópodos en márgenes con mayor diversidad vegetal (Thomas & Marshall 1999; Varchola & Dunn 1999; Asteraki et al. 2004). Los depredadores, en cambio, tuvieron igual riqueza en ambientes marginales que en cultivos, lo cual sugiere que ambos tipos de hábitat ofrecen la misma cantidad de nichos para este tipo de organismos. Este patrón indica que la composición de la comunidad en su estructura trófica varió entre ambientes, con mayor proporción de especies depredadoras en los cultivos que en los márgenes y terrazas. Esto pudo deberse a que las especies presa son más abundantes en el cultivo y permitirían sostener una red trófica con más especies depredadoras. La mayor abundancia de artrópodos encontrada en este trabajo en los ambientes de margen coincide con otros estudios similares realizados en márgenes de cultivos con arañas (Lemke & Poheling 2002; Liljesthröm et al. 2002), Carabidae (Varchola & Dunn 1999), depredadores (Kemp & Barret 1989; Dennis & Fry 1992) y mariposas (de Snoo 1999). Al igual que en el caso de la riqueza, esto podría deberse a una mayor diversidad estructural del hábitat en los márgenes, la cual -se ha encontrado- aumenta la abundancia de artrópodos (Borges & Brown 2001; pero ver Perner et al. 2005).
Cuando se analizó cada fecha de muestreo por separado, la falta de diferencias desde el comienzo de la floración hasta el estadio de llenado de grano pudo haberse relacionado con condiciones microclimáticas dentro del lote que favorecerían la dispersión de individuos, como lo han sugerido Dennis et al. (2000) para depredadores. La diferencia de abundancia luego de la aplicación de insecticidas pudo deberse a que las poblaciones más afectadas fueron las de los cultivos. Estas diferencias desaparecieron en el siguiente muestreo, posiblemente debido a que el efecto del insecticida había pasado y nuevos individuos se dispersaron desde los hábitats marginales.
La abundancia absoluta de depredadores resultó similar tanto en los márgenes como en los cultivos, y fue independiente de la presencia o no de las terrazas. Sin embargo, la proporción de depredadores respecto del total fue mayor en los cultivos, lo que indica un cambio en la composición trófica de la comunidad entre los distintos ambientes. Estas diferencias mostraron una estacionalidad a lo largo del ciclo del cultivo, ya que los depredadores fueron más abundantes en los márgenes al comienzo y al final del ciclo. Este patrón pudo deberse al movimiento de individuos entre los márgenes y el cultivo, dado que los márgenes actúan como fuente o sumidero según cuán atractivos sean para los artrópodos (Corbett & Plant 1993).
Los depredadores se pudieron mover al cultivo durante su floración y primeros estadios de llenado de granos, cuando sus presas son allí más abundantes (Massaro et al. 1992). En los períodos en los que las condiciones en el cultivo no fueron favorables, los depredadores pudieron haber buscado refugio en los márgenes. Se ha visto en otros trabajos que los márgenes pueden actuar como refugios ante la aplicación de insecticidas (Lee et al. 2001) o durante períodos de sobreinvernación (Lemke & Poehling 2002). A pesar de que este estudio no fue diseñado para detectar movimientos de poblaciones entre distintos tipos de hábitat, otros autores demostraron que el intercambio de individuos en los bordes de cultivos es un evento común (Duelli et al. 1990; Thomas et al. 2002). Dennis (1991), trabajando con trampas de caída direccionales, encontró un movimiento neto de depredadores hacia el cultivo. Algunas especies estudiadas fueron más abundantes en el margen del lote al comienzo de la estación, luego de lo cual sus valores se nivelaron. Estudios posteriores deberían concentrarse en probar la existencia de estos movimientos entre terrazas y cultivos de soja, para lo cual es necesario otro diseño experimental distinto al que utilizamos en este trabajo.
La falta de diferencias de abundancia entre los márgenes y el cultivo sugiere que las condiciones en ambos ambientes no eran tan desfavorables para los depredadores como pensamos en un principio. Este estudio se llevó a cabo en lotes cultivados bajo el sistema de siembra directa, que -en general- favorece la diversidad de insectos en cultivos (Marasas et al. 2001; Zaccagnini & Calamari 2001). Por lo tanto, es posible que haya existido un gradiente suave entre las condiciones ambientales en los márgenes y en los cultivos de soja, lo que facilitó la inmigración de artrópodos al cultivo (Lemke & Phoeling 2002).
Debido a la dificultad en la identificación de especies de artrópodos se propuso el uso de taxones indicadores para facilitar el monitoreo de biodiversidad (Büchs 2003). Aún cuando se usa un único método de muestreo (trampas de caída), el número de especies e individuos recolectados resulta demasiado grande para una evaluación eficiente de la biodiversidad. En este estudio propusimos dos grupos potenciales como bioindicadores: depredadores y carábidos. Nuestros resultados sugieren que los depredadores podrían funcionar como buenos indicadores de la riqueza total de artrópodos. Por el contrario, su valor como indicadores de abundancia es menos confiable, probablemente a causa de que, por su posición trófica, los depredadores estuvieron menos representados en los datos. Esto los hace naturalmente menos abundantes. Sin embargo, los patrones de los resultados de este estudio no son siempre similares para los depredadores y para el total de artrópodos. Es en cierta medida, era esperable que distintos grupos funcionales tuvieran respuestas diferentes al ambiente o, incluso, dentro de un mismo grupo funcional (Lindenmayer et al. 2000). Las especies de Carabidae y arañas, por ejemplo, tienen respuestas individualistas que no son extrapolables a todo el conjunto de especies (Pearce & Venier 2006). Por esta razón, si el objetivo del uso de indicadores es sobre efectos del ambiente, no sería recomendable usar los depredadores. Nuestro diseño experimental presentó la ventaja de ser representativo de las condiciones reales al trabajar en lotes a escala comercial, ya que los estudios en parcelas experimentales (por lo general de pocos metros cuadrados) conllevan el riesgo de una sobreestimación de los efectos de borde, al tener una mayor relación borde/área (Corbett & Plant 1993). Las debilidades de nuestra aproximación, en cambio, están dadas por el número reducido de repeticiones, que podría haber limitado nuestra capacidad de detectar algunos efectos.
Apéndice. Abundancia de morfoespecies y roles funcionales. Det: detritívoros, Fit: fitófagos, Pre: depredadores, Par: parasitoides y Mic: micetófagos.
Appendix. Morphospecies abundance and functional roles. Det: detritivores, Fit: phytophages, Pre: predators; Par: parasitoids; Mic: mycetophages.







AGRADECIMIENTOS
Agradecemos a las autoridades y personal de la EEA Paraná del INTA por las facilidades provistas, a los dueños de los campos en los que fueron conducidos los estudios, y a los extensionistas W. Mancuso, R. de Carli y G. Pasgal por facilitar la comunicación con los agricultores. N. Calamari ayudó con el relevamiento y selección de lotes. B. Jaubert, L. Cappellacci, P. Garaffa, L. Solari y M. Viola asistieron en el trabajo de campo y laboratorio. A. Cicchino, G. Montero, M. Ramírez y A. Saluso identificaron algunos especímenes recolectados. A. Rosón muestreó la vegetación e identificó las especies. S. Perelman asistió en los análisis estadísticos. Tres revisores han mejorado versiones anteriores de este manuscrito. Este estudio formó parte de la tesis de licenciatura de FW y fue financiado por el Proyecto INTA – USFWS # 2534.
BIBLIOGRAFÍA
ALTIERI, MA. 1999. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 74: 19- 31. [ Links ]
ASTERAKI, E J; BJ HART; TC INGS & WJ MANLEY. 2004. Factors influencing the plant and invertebrate diversity of arable field margins. Agric. Ecosyst. Environ. 102:219-231 [ Links ]
BORGES, PAV & VK BROWN. 2001. Phytophagous insects and web-building spiders in relation to pasture vegetation complexity. Ecography 24: 68- 82. [ Links ]
BÜCHS, W. 2003. Biodiversity and agri-environmental indicators - general scopes and skills with special reference to the habitat level. Agric. Ecosyst. Environ. 98:35-78. [ Links ]
BURKART, R; NO BÁRBARO; RO SÁNCHEZ & DA GÓMEZ. 1999. Eco-regiones de la Argentina. Administración de Parques Nacionales, 42 pp. [ Links ]
CARMONA, DM & DA LANDIS. 1999. Influence of refuge habitats and cover crops on seasonal activity-density of ground beetles (Coleoptera: Carabidae) in field crops. Environ. Entomol. 258: 1145-1153. [ Links ]
CHAO, A & SM LEE. 1992. Estimating the number of classes via sample coverage. J. Am. Stat. Assoc. 87:210-217. [ Links ]
CHAO, A & TJ SHEN. 2003. Program SPADE (Species Prediction And Diversity Estimation). Programa y Guía del Usuario publicado en http://chao.stat.nthu.edu.tw. [ Links ]
CHAPIN III, FS; ES ZABALETA; VT EVINER; RL NAYLOR; PM VITOUSEK ET AL. 2000. Consequences of changing biodiversity. Nature 405:234-242. [ Links ]
CORBETT, A & RE PLANT. 1993. Role of movement in the response of natural enemies to agroecosystem diversification: a theoretical evaluation. Environ. Entomol. 22:519-531. [ Links ]
DE LA PEÑA, NM; A BUTET; Y DELETTRE; P MORANT & F BUREL. 2003. Landscape context and carabid beetles (Coleoptera: Carabidae) communities of hedgerows in western France. Agric. Ecosyst. Environ. 94:59-72. [ Links ]
DE SNOO, JR. 1999. Unsprayed field margins: effects on environment, biodiversity and agricultural practice. Landscape Urban Plan. 46:151-160. [ Links ]
DENNIS, P; GLA FRY & A ANDERSEN. 2000. The impact of field boundary habitats on the diversity and abundance of natural enemies in cereals. Pp. 193-212 en: Ekbom, B; M Irwin & Y Roberts (eds.). Interchanges of insects. Kluwer Academic Publishers, Netherlands. [ Links ]
DENNIS, P & GLA FRY. 1992. Field margins: can they enhance natural enemy population densities and general arthropod diversity on farmland? Agric. Ecosyst. Environ. 40:95-115. [ Links ]
DENNIS, P. 1991. The temporal and spatial distribution of arthropod predators of the aphids Rhopalosiphum padi (L) and Sitobion avenae (F) in cereals next to field-margin habitats. Norwegian Journal of Agricultural Sciences 5:79-88. [ Links ]
DUELLI, P & MK OBRIST. 2003. Biodiversity indicators: the choice of values and measures. Agric. Ecosyst. Environ. 98:87-98. [ Links ]
DUELLI, P; MK OBRIST & DR SCHMATZ. 1999. Biodiversity evaluation in agricultural landscapes: above-ground insects. Agric. Ecosyst. Environ. 74: 33-64. [ Links ]
DUELLI, P; M STUDER; I MARCHAND & S JACOBS. 1990. Population movements of arthropods between natural and cultivated areas. Biol. Conserv. 54: 193-207. [ Links ]
FAHRIG, L & I JONSEN. 1998. Effect of habitat patch characteristics on abundance and diversity of insects in an agricultural landscape. Ecosystems 1:197-205. [ Links ]
FLEISHMAN, E; JR THOMPSON; R MAC NALLY; DD MURPHY & JP FAY. 2005. Using indicator species to predict species richness of multiple taxonomic groups. Conserv. Biol. 19:1125-1137. [ Links ]
IRMLER, U. 2003. The spatial and temporal pattern of carabid beetles on arable fields in northern Germany (Schleswig-Holstein) and their value as ecological indicators. Agric. Ecosyst. Environ. 98:141-151. [ Links ]
KATI, V; P DEVILLERS; M DUFRÊNE; A LEGAKIS; D VOKOU ET AL. 2003. Testing the value of six taxonomic groups as biodiversity indicators at a local scale. Conserv. Biol. 18:667-675. [ Links ]
KAUTZ, G; M ZIMMER & W TOPP. 2002. Does Porcellio scaber (Isopoda: Oniscidea) gain from coprophagy? Soil Biol. Biochem. 34:1253-1259. [ Links ]
KEMP, JC & GW BARRETT. 1989. Spatial patterning: impact of uncultivated corridors on arthropod populations within soybean agroecosystems. Ecology 70:114-128. [ Links ]
LEE, JC; FD MENALLED & DA LANDIS. 2001. Refuge habitats modify impact of insecticide disturbance on carabid beetle communities. J. Appl. Ecol. 38: 472-483. [ Links ]
LEMKE, A & HM POEHLING. 2002. Sown weed strips in cereal fields: overwintering site and "source" habitat for Oedothorax apicatus (Blackwall) and Erigone atra (Blackwall) (Araneae: Erigonidae). Agric. Ecosyst. Environ. 90:67-80. [ Links ]
LILJESTHRÖM, G; E MINERVINO; D CASTRO & A GONZÁLEZ. 2002. La comunidad de arañas del cultivo de soja en la Provincia de Buenos Aires, Argentina. Neotropical Entomology 31:197-210. [ Links ]
LINDENMAYER, DB; CR MARGULES & DB BOTKIN. 2000. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 14: 941-950. [ Links ]
LÖVEI, GL & KD SUNDERLAND. 1996. Ecology and behavior of ground beetles (Coleoptera: Carabidae). Annu. Rev. Entomol. 41:231-256. [ Links ]
MAC NALLY, R & E FLEISHMAN. 2004. A successful predictive model of species richness based on indicator species. Conserv. Biol. 18:646-654. [ Links ]
MARASAS, ME; SJ SARANDÓN & AC CICCHINO. 2001. Changes in arthropod functional group in a wheat crop under conventional and no tillage systems in Argentina. Appl. Soil Ecol. 18:61-68. [ Links ]
MARC, P; A CANARD & F YSNEL. 1999. Spiders (Araneae) useful for pest limitation and bioindication. Agric. Ecosyst. Environ. 74:229-273. [ Links ]
MASSARO, RA; A MAI; G ROSSI & G PRIETO. 1992. Evolucion de poblaciones de insectos fitofagos en soja con sistemas de labranza minima y siembra directa en el sur de Santa Fe. Ciclo 1991/92. VIII Jornadas Fitosanitarias Argentinas, Paraná. [ Links ]
OERTLI, S; A MÜLLER; D STEINER; A BREITENSTEIN & S DORN. 2005. Cross-taxon congruence of species diversity and community similarity among three insect taxa in a mosaic landscape. Biol. Conserv. 126:195-205. [ Links ]
OLIVER, I & AJ BEATTIE. 1996. Designing a cost-effective invertebrate survey: a test of methods for rapid assessment of biodiversity. Ecol. Appl. 6:594-607. [ Links ]
PEARCE, JL & LA VENIER. 2006. The use of ground beetles (Coleoptera: Carabidae) and spiders (Araneae) as bioindicators of sustainable forest management: A review. Ecological Indicators 6: 780-793. [ Links ]
PERNER, J; C WYTRYKUSH; A KAHMEN; N BUCHMANN; I EGERER ET AL. 2005. Effects of plant diversity, plant productivity and habitat parameters on arthropod abundance in montane European grasslands. Ecography 28:429-442. [ Links ]
PIMENTEL, D; C WILSON; C MCCULLUM; R HUANG; P DWEN ET AL. 1997. Economic and environmental benefits of biodiversity. Bioscience 47:747-757. [ Links ]
SALUSO, A. 2004. Determinación del Nivel de Daño Económico y plan de decisión secuencial para el manejo de Armadillidium vulgare (Latreille, 1804) (Crustacea: Isopoda) en soja. Tesis de Magister, Universidad Nacional de La Rioja, 75 pp. [ Links ]
SOKAL, RR & FJ ROHLF. 1995. Biometry. The principles and practice of statistics in biological research. Freeman & Company, New York, 887 pp. [ Links ]
SOUSA, JP; JV VINGADA; S LOUREIRO; MM DA GAMA & AMVM SOARES. 1998. Effects of introduced exotic tree species on growth, consumption and assimilation rates of the soil detritivore Porcellio dilatatus (Crustacea: Isopoda). Appl. Soil. Ecol. 9: 399-403. [ Links ]
SWIFT, MJ; AMN IZAC & M VAN NOORDWIJK. 2004. Biodiversity and ecosystem services in agricultural landscapes - are we asking the right questions? Agric. Ecosyst. Environ. 104:113-134. [ Links ]
THOMAS, CFG; JM HOLLAND & NJ BROWN. 2002. The spatial distribution of carabid beetles in agricultural landscapes. Pp 305-344 en: Holland, JM (ed.). The agroecology of carabid beetles. Intercept, Andover, U.K. [ Links ]
THOMAS, CFG & EJP MARSHALL. 1999. Arthropod abundance and diversity in differently vegetated margins of arable fields. Agric. Ecosyst. Environ. 72:131-144. [ Links ]
VARCHOLA, JM & JP DUNN. 1999. Changes in ground beetle (Coleoptera: Carabidae) assemblages in farming systems bordered by complex or simple roadside vegetation. Agric. Ecoyst. Environ. 73: 41- 49. [ Links ]
ZACCAGNINI, ME & NC CALAMARI. 2001. Labranzas conservacionistas, siembra directa y biodiversidad. Pp. 29-68 en: Panigatti, JL; D Buschiazzo & H Marelli (eds.). Siembra Directa II. INTA, Buenos Aires. [ Links ]
ZACCAGNINI, ME; J DECARRE; A GOIJMAN; L SOLARI; R SUÁREZ ET AL. 2007. Efecto de la heterogeneidad ambiental de terrazas y bordes vegetados sobre la biodiversidad animal en campos de soja en Entre Ríos. Pp. 232 en: Caviglia, OP; OF Paparotti & MC Sasal (eds.). Agricultura Sustentable en Entre Ríos. Ediciones INTA. Buenos Aires. [ Links ]


{kind=link}