










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.19 n.2 Córdoba mayo/ago. 2009
TRABAJOS ORIGINALES
Evaluación de estrategias de rehabilitación de pastizales áridos
Emiliano Quiroga*, Lisandro Blanco & Enrique Orionte
* INTA, Estación Experimental Agropecuaria La Rioja, Argentina.
INTA, Estación Experimental Agropecuaria La Rioja, CC 26, CP 5380, Chamical, La Rioja, Argentina.
Dirección actual: INTA, EEA Catamarca, Ruta Provincial 33, km 4, (4705) Sumalao, Valle Viejo, Catamarca, Argentina. Email: equiroga@correo.inta.gov.ar
Recibido: 19 de noviembre de 2008;
Fin de arbitraje: 15 de marzo de 2009;
Revisión recibida: 20 de abril de 2009;
Aceptado: 25 de mayo de 2009
RESUMEN. En los pastizales del Chaco Árido argentino se ha perdido biodiversidad y productividad a causa del sobrepastoreo por animales domésticos. El objetivo de este trabajo fue evaluar algunos tratamientos de rehabilitación que podrían aumentar la densidad de gramíneas forrajeras en un pastizal sobrepastoreado de la región. A partir de la idea de que la compactación del suelo, la falta de semillas en el banco y la falta de mantillo son factores que limitan el establecimiento de las plántulas, realizamos un experimento factorial a campo que combinó poceado del suelo, agregado de semillas y acción conjunta de reducción de competencia y agregado de mantillo de arbustos. Durante cuatro años evaluamos la densidad de plántulas y plantas de las gramíneas agregadas y del total de gramíneas forrajeras. Sólo el agregado de semillas aumentó la densidad de plántulas y plantas de estas especies. Durante el primer año registramos mayor densidad de plántulas en las parcelas con agregado de semillas (principalmente Pappophorum spp.) y una mayor densidad de plantas adultas durante los primeros dos años (atribuible al establecimiento casi exclusivo de P. philippianum). A partir del tercer año (año seco) se observó una disminución de 70-80% en la densidad de plantas en todos los tratamientos, si bien persistieron las diferencias debidas al agregado de semillas. Nuestros resultados sugieren que, además de las condiciones climáticas adversas, la falta de semillas en el banco del suelo restringe la recuperación de los pastizales sobrepastoreados en el Chaco Árido, y que Pappophorum philippianum sería una especie promisoria a utilizar con fines de rehabilitación.
Palabras clave: Gramíneas forrajeras; Densidad de plantas; Recuperación de la vegetación; Áreas degradadas; Chaco Árido.
ABSTRACT. Testing rehabilitation strategies for arid rangelands: Overgrazing by domestic cattle has caused loss of plant diversity and primary productivity in rangelands of the Arid Chaco (Argentina). The objective of our study was to evaluate a number of rehabilitation strategies that might increase the density of forage grasses in an overgrazed rangeland in this region. Based on the hypothesis that soil compaction, lack of seeds and litter cover constrain plant establishment we performed a field factorial experiment, that combined soil pitting, seed addition and the combined effect of competition release and litter addition. During four years, we evaluated seedling and plant density of added grasses and all forage grasses. Seed addition was the only factor that increased seedling and plant density, with Pappophorum spp. and P. philippianum accounting for the main variation in seedling and plant density, respectively. During the third year (a dry year), there was 70-80% decrease in plant density in all treatments, although the differences due to the addition of seeds was still apparent. Our results suggest that, in addition to adverse climatic conditions, the lack of seeds in the soil bank restricts the recovery of overgrazed rangelands in the Arid Chaco. Pappophorum philippianum seems to be a promising species for the rehabilitation of these rangelands.
Keywords: Forage grasses; Plant density; Vegetation recovery; Degraded areas; Arid Chaco.
INTRODUCCIÓN
Los ecosistemas áridos y semiáridos cubren una gran superficie del planeta. Su degradación a causa del uso antrópico es un problema serio y su rehabilitación es de gran importancia desde el punto de vista productivo y ecológico (Aronson et al. 1993; Maestre et al. 2001). Las estrategias más frecuentes para recuperar la vegetación en áreas degradadas incluyen la siembra de especies forrajeras (González & Dodd 1979; Passera et al. 1992; Blanco et al. 2005), tratamientos para mejorar las características del suelo (González & Dodd 1979; Blanco et al. 2005), o la creación de micrositios favorables para el establecimiento y el crecimiento de las plantas (Passera et al. 1992; Handa & Jefferies 2000; Zuleta et al. 2003). En general, la probabilidad de lograr resultados exitosos en estudios orientados a recuperar la vegetación de áreas degradadas tiende a disminuir con la marginalidad del ambiente en el cual se realizan [i.e., menor precipitación media anual, menor disponibilidad de nutrientes en el suelo, estado de degradación avanzado, etc. (Westoby et al. 1989; Holmgren et al. 2001)].
En la región del Chaco Árido, ubicada en el noroeste de la Argentina, la ganadería de cría bovina y caprina es la actividad agropecuaria más difundida e involucra a unos 10000 productores (Diez et al. 1991; Blanco et al. 2005). La dieta del ganado se basa casi exclusivamente sobre los recursos forrajeros del pastizal nativo (Anderson et al. 1980). Actualmente, la productividad forrajera de estos ambientes se encuentra por debajo de su potencial debido al deterioro de la vegetación a causa del sobrepastoreo (Biurrun 1988), que afecta en especial a las gramíneas forrajeras (Anderson et al. 1980). A pesar de la importancia productiva y ecológica que representa este problema, faltan estudios que evalúen técnicas orientadas a aumentar la cobertura de este grupo de especies en los campos ganaderos de la región. En un sitio clausurado al pastoreo por 20 años ubicado en el ecotono entre el Chaco Árido y el Monte, Passera et al. (1992) lograron un mejoramiento de la capacidad forrajera mediante la remoción de arbustos (principalmente Larrea cuneifolia) y la creación de mantillo con el material vegetal removido. En cambio, no encontraron que el agregado de semillas de gramíneas nativas promoviera la recuperación forrajera del ecosistema, y sugieren que en dicho ambiente, la competencia de los arbustos sería un factor más limitante que la disponibilidad de semillas de gramíneas nativas. Sin embargo, como éstos y otros autores han reconocido (Holmgren et al. 2001; Handa et al. 2002; Suding et al. 2004), en áreas sobrepastoreadas, la importancia de la competencia ejercida por arbustos podría jugar un papel menor frente a otras limitantes tales como la compactación del suelo, la falta de semillas en el banco del suelo o la falta de sitios seguros para el reclutamiento de plántulas.
El objetivo del presente estudio fue evaluar distintos tratamientos de rehabilitación sobre un pastizal degradado por sobrepastoreo en la región del Chaco Árido, sobre la base de la hipótesis que en ambientes sobrepastoreados, la compactación del suelo, la falta de semillas en el banco del suelo, la competencia de los arbustos y la falta de mantillo son factores que limitan el establecimiento de las gramíneas forrajeras nativas (González & Dodd 1979; Call & Roundy 1991; Passera et al. 1992; Zuleta et al. 2003; Blanco et al. 2005).
MATERIALES Y MÉTODOS
Sitio de estudio
La región del Chaco Árido posee una superficie aproximada de 100000 km² y se encuentra enmarcada por un conjunto de sierras pampeanas que superan los 2000 m sobre el nivel del mar (Morello et al. 1985). El clima es subtropical seco, de veranos cálidos con 20 a 25 días con temperaturas superiores a 40 °C, e inviernos templados con solo 5 a 10 días con temperaturas bajo cero (Prohasca 1959). La temperatura media mensual es de 11 ºC en julio y 26 ºC en enero. La precipitación media anual varía entre 600 mm en el este y 300 mm en el oeste (Morello et al. 1985), con el 80% concentrado entre noviembre y marzo. En general, los suelos poseen textura gruesa, aunque su granulometría disminuye desde las sierras a las zonas más bajas. Los contenidos de materia orgánica son bajos (<1% en los primeros 20 cm de suelo) y el pH varía por lo general entre neutro y básico (Gómez et al. 1993). El tipo de vegetación es un arbustal xerófilo subtropical, con una estructura arbórea discontinua dominada por Aspidosperma quebracho blanco y Prosopis spp. (Anderson et al. 1980; Morello et al. 1985). Entre los arbustos se destacan los géneros Larrea, Mimozyganthus, Senna y Capparis. Por su parte, el estrato herbáceo esta conformado principalmente por gramíneas C4 perennes de los géneros Trichloris, Pappophorum, Digitaria y Setaria (Ragonese & Castiglioni 1970; Morello et al. 1985).
La experiencia se realizó en un área muy degradada por sobrepastoreo debido a su cercanía a una aguada. El área pertenece al campo ganadero "La Dormida", al oeste del Chaco Árido (30º46'12'' S - 66º52'48'' W), sobre la isoyeta de 300 mm, zona en la que se exacerban las limitaciones ambientales (Cabido et al. 1993; Blanco et al. 2008). La precipitación total durante las estaciones de crecimiento 2002/2003, 2003/2004 y 2005/2006 fue apenas superior al promedio del lugar (320 mm, para el período 1986-2000, Agencia de Extensión Rural de INTA Portezuelo). Inclusive, durante la primera estación de crecimiento (un mes después del inicio del ensayo) tuvo lugar una lluvia de 115 mm. Por su parte, la temporada 2004/2005 fue muy seca (Figura 1).
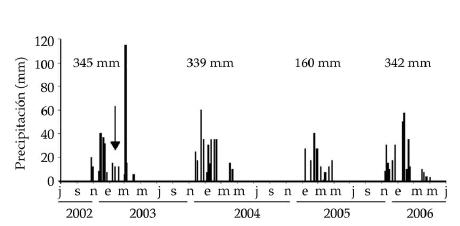
Figura 1. Precipitaciones caidas en el sitio experimental entre julio 2002 y junio 2006, en el sector cercano a la aguada del campo "La Dormida" (30.77º S, 60.80º W). Cada línea vertical representa un evento; valores en la parte superior: precipitaciones anuales totales (períodos julio-junio). Precipitación media anual: 320 mm (período 1986-2000, Agencia de Extensión Rural, INTA Portezuelo). La flecha señala la fecha de aplicación de los tratamientos.
Figure 1. Precipitation in the experimental site (sector near to the watering point of the "La Dormida" cattle ranch; 30.77º S, 60.80º W), from July of 2002 to June of 2006. Each rain event has been represented by a vertical line; values in the upper portion of the figure: yearly total precipitation (July to June). Mean annual precipitation: 320 mm (period 1986-2000, Agencia de Extensión Rural INTA Portezuelo). The arrow shows the date of the treatments.
Diseño experimental y variables medidas
Se realizó un experimento con diseño en bloques completos aleatorizados (n=6) y arreglo factorial de tres factores: poceado, agregado de semillas y de mantillo (cada uno con dos niveles: si-no). Los tratamientos evaluados resultaron de la combinación de dichos factores, lo que determinó un total de 8 tratamientos (testigo; poceado; semilla; mantillo; poceado y semilla; semilla y mantillo; poceado y mantillo; poceado, semilla y mantillo). Durante el experimento, el sitio fue clausurado para evitar el ingreso del ganado. El poceado removió la compactación superficial del terreno; el agregado de semillas aumentó la disponibilidad de semillas de gramíneas forrajeras en el banco del suelo, y la aplicación de mantillo simultáneamente incrementó la existencia de sitios favorables para el reclutamiento de plántulas y redujo la competencia ejercida por los arbustos (dado que el mantillo era de ramas de arbustos obtenidas en cada una de las parcelas tratadas). Los tratamientos se aplicaron el 28 de enero de 2003, en parcelas de 20 m x 10 m, en el siguiente orden: poceado, agregado de semillas y aplicación de mantillo.
El poceado se realizó con un rolo constituido por un cilindro de 1.1 m de ancho y 0.9 m de diámetro en el cual se insertan en forma radial brazos de acero con puntas en forma de pala (peso total: 244 kg), traccionado por un burro, teniendo especial cuidado de no dañar la vegetación leñosa y con una densidad aproximada de 8 pozos/m², cada uno de 5-10 cm de diámetro y alrededor de 5 cm de profundidad.
El agregado de semillas consistió en distribuir de forma manual una mezcla de semillas de las siguientes gramíneas forrajeras perennes nativas: Trichloris crinita, Digitaria californica y Pappophorum spp. (mezcla de P. caespitosum y P. philippianum) (Passera et al. 1992), con una densidad de 200 semillas "germinables"/m² en cada caso; es decir, un total de 600 semillas "germinables"/m². Las semillas se cosecharon durante el año previo al inicio del ensayo, de poblaciones de la región. Las de Trichroris crinita y Digitaria californica pertenecían a una colección establecida en la Experimental de INTA La Rioja (30º28'57'' S - 66º07'26'' W) y las de Pappophorum caespitosum y P. philippianum se recolectaron en márgenes de la Ruta Nacional 79 (31º19'11'' S - 66º16'58'' W y 30º21'16'' S - 66º18'13'' W, respectivamente). Antes de iniciar el ensayo se realizaron pruebas de germinación en cajas de Petri (3 cajas con 100 semillas/especie) con papel de filtro humedecido con agua para mantener condiciones de humedad adecuadas para la germinación. Se mantuvieron a 25-30 ºC (Marone et al. 2000) y se contaron las semillas germinadas hasta el cese de la germinación (alrededor de 30 días). El promedio de germinación acumulada fue: Trichloris crinita=50%; Digitaria californica= 33%; Pappophorum caespitosum=50%; Pappophorum philippianum=51%.
La aplicación de mantillo se realizó cortando ramas de arbustos ubicados dentro de la parcela (principalmente Larrea divaricada, Larrea cuneifolia y Mymoziganthus carinatus), y distribuyéndolas sobre el terreno fuera de la proyección de su copa. Al momento de la aplicación de este tratamiento los arbustos se encontraban en activo crecimiento, por lo que poseían follaje verde. La cobertura aérea "en pie" de arbustos en parcelas con este tratamiento se redujo en promedio 15% (de 40% a 25%). El mantillo cubrió aproximadamente 25% de la superficie de la parcela [mediciones realizadas en junio 2003, en 5 marcos de 0.1 m2/parcela, mediante el método de Daubenmire (1959)].
Para caracterizar el stand de gramíneas forrajeras previo a la aplicación de los tratamientos, en cada parcela se midió la densidad de cada especie en cuatro marcos de 0.1 m² colocados cada 2.5 m sobre una transecta de 10 m. Luego de aplicados los tratamientos, la densidad de gramíneas forrajeras se midió en cada parcela al finalizar cada estación de crecimiento durante cuatro años (junio de 2003, 2004, 2005 y 2006), en cinco marcos de 0.1 m² ubicados cada 2 m sobre una transecta fija de 10 m. Las plántulas (individuos con un único macollo, sólo se observaron en junio de 2003) se contaron por separado de las plantas (de 2 ó más macollos). Para los análisis sólo se tuvieron en cuenta las gramíneas forrajeras perennes. De todas formas, durante las mediciones fue raro observar gramíneas anuales, y las pocas que fueron halladas carecían de valor forrajero (Bouteloua aristidoides y Aristida adscencionis). Se calculó la densidad de plántulas y de plantas de cada uno de los siguientes grupos: a) cada especie de gramínea agregada b) total de gramíneas agregadas y c) total de gramíneas forrajeras.
Análisis de los datos
Los datos se analizaron sobre la base de un modelo mixto (Littel et al. 1996), mediante el procedimiento MIXED del programa estadístico SAS (SAS Institute 1996). Como factores de efectos fijos del modelo se incluyeron: poceado, semilla, mantillo, año (este último únicamente para plantas ya que sólo se observaron plántulas en junio de 2003), y las interacciones dobles, triples y cuádruples (esta última sólo para plantas) resultantes de las combinaciones posibles entre ellos. Por su parte, se incluyeron como factores de efecto aleatorio los bloques y las parcelas (este último sólo para plantas) anidadas en la interacción poceado x semilla x mantillo (Littel et al. 1996). El análisis de la densidad total de plantas de gramíneas forrajeras se realizó usando como covariable la densidad medida para dicho grupo antes de aplicarse los tratamientos. Para las otras variables no se utilizó covariable debido a que los análisis preliminares mostraron que las densidades medidas antes del ensayo no eran significativas (P>0.15) al ser incluidas en el modelo. En cada análisis se utilizó la opción KENWARDROGER para estimar los grados de libertad, y la sentencia LSMEANS para estimar medias y errores estándar (SAS Institute 1996).
RESULTADOS
Sólo se observaron plántulas en junio de 2003, y todas correspondieron a las especies agregadas. Las parcelas con agregado de semillas mostraron mayor densidad total de plántulas que las parcelas sin agregado de semillas (Tabla 1; Figura 2). La mayor parte de esta diferencias estuvo asociada a la presencia de plántulas del género Pappophorum, seguida por Digitaria californica, y con un escaso aporte por Trichloris crinita (Tabla 1). Contrariamente a lo esperado, no se observó efecto significativo del poceado y de la aplicación de mantillo sobre la densidad de plántulas. Tampoco se observó interacción entre los factores semilla, poceado y mantillo (Tabla 1).
Tabla 1. Resultados del análisis del modelo mixto para densidad de plántulas de Pappophorum spp., Digitaria californica, Trichloris crinita, y el total de gramíneas agregadas. Se muestran los valores de F y de probabilidad para los factores de efectos fijos (en negrita: P<0.05) y los valores de Z y de probabilidad para los factores de efectos aleatorios.
Table 1. Results for the mixed model analysis for seedling density of Pappophorum spp., Digitaria californica, Trichloris crinita and total added grasses. F and probability values for fixed-effect factors are showed (in bold: P<0.05), Z and probability values for random-effect factors are also indicated.
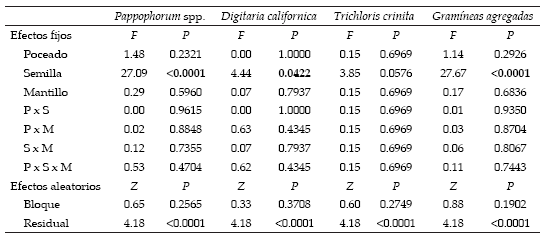
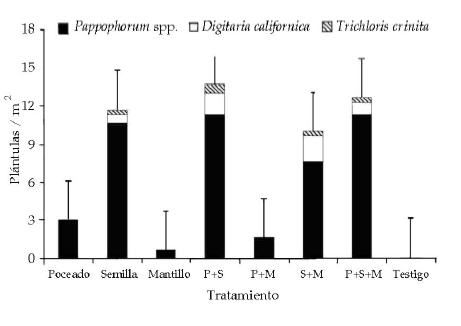
Figura 2. Densidad media de plántulas de cada especie agregada (plántulas/m2) en los distintos tratamientos (junio 2003, única fecha en que se censaron plántulas). Las líneas verticales sobre cada barra corresponden a los EE de la densidad total de especies agregadas. Los valores de P figuran en la Tabla 1.
Figure 2. Mean seedling density of each added species (seedlings/m2) in the evaluated treatments (June 2003, only date in which seedlings were recorded). Vertical lines represent SE for the total density of added species. P-values are shown in Table 1.
El aporte de semillas permitió incrementar la densidad de plantas de Pappophorum philippianum (Tabla 2). La diferencia entre las parcelas con y sin agregado de semillas fue de 2 a 3 plantas/m2 durante los dos primeros años, y de 1 planta/m2 durante los dos últimos (Figura 3). A partir del tercer año (año seco, 2004/2005) se registró una disminución en la cantidad de plantas de P. philippianum (-3 a -5 plantas/m2) en todos los tratamientos (Tabla 2). Pese su pequeña magnitud, la densidad en las parcelas con agregado de semillas duplicó la densidad en las parcelas sin agregado. Por otra parte, ni el poceado, ni la aplicación de mantillo, ni sus interacciones tuvieron efecto sobre la densidad de plantas de P. philippianum (Tabla 2).
Tabla 2. Resultados del modelo mixto para densidad de plantas de Pappophorum philippianum, total de gramíneas agregadas y total de gramíneas forrajeras. Se muestran los valores de F y de probabilidad para los factores de efectos fijos (en negrita: P<0.05) y los valores de Z y de probabilidad para los factores de efectos aleatorios. Para densidad de plantas del total de gramíneas forrajeras se usó como covariable la densidad de dicho grupo medida antes del inicio del ensayo.
Table 2. Results of the mixed model analysis for plant density of Pappophorum philippianum,total added grasses and total forage grasses. F and probability values for fixed-effect factors are showed (in bold: P<0.05), Z and probability values for random-effect factors are also indicated. For the total density of forage grasses we used as covariate the density of this group measured at the beginning of the experiment.
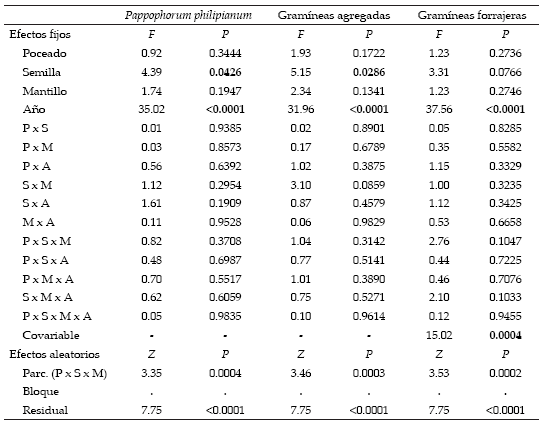
No se observaron plantas de Pappophorum caespitosum en ninguna de las parcelas. Las densidades de Digitaria californica y Trichloris crinita fueron muy bajas (<0.8 y <0.2 plantas/ m2, respectivamente) y no fueron modificadas por el poceado, el agregado de semillas, la aplicación de mantillo o los años ni se observó efecto interactivo de estos factores (Tabla 2). Como consecuencia, la densidad total de plantas de gramíneas agregadas tuvo un comportamiento similar a la de Pappophorum philippianum: sólo varió por efecto del agregado de semillas y de los años (Tabla 2). La diferencia en la densidad total de plantas de gramíneas agregadas entre parcelas con y sin agregado fue de 2 a 3 plantas/m2 durante los dos primeros años y de 1 a 1.5 plantas/m2 en los dos últimos. También se observó una disminución (-3 a -5 plantas/m2) a partir del año seco (2004/2005) en todos los tratamientos (Figura 3). El poceado, la aplicación de mantillo y las interacciones evaluadas no tuvieron efecto sobre la densidad total de plantas de gramíneas agregadas (Tabla 2).

Figura 3. Dinámica de la densidad promedio de plantas de Pappophorum philippianum (arriba), de especies de gramíneas agregadas (centro), y de especies de gramíneas forrajeras (abajo). Los EE de las medias fueron: 1.5 (arriba), 1.6 (centro) y 1.8 (abajo). Los valores de P figuran en la Tabla 2. Gráficos en parte superior derecha de cada figura: densidad promedio en parcelas con y sin agregado de semillas; EE: 0.7 (arriba), 0.8 (centro) y 0.9 (abajo).
Figure 3. Dynamics of average plant density of Pappophorum philippianum (upper), added grasses (center), and forage grasses (bottom). Estimated SE for these means were 1.5 (upper), 1.6 (center), and 1.8 (bottom). P-values are shown in Table 2. Inserted graphics in upper right corner of each figure: average plant density on plots with and without addition of seeds; SE were: 0.7 (upper), 0.8 (center) and 0.9 (bottom).
La densidad de gramíneas forrajeras nativas de especies no agregadas (Diplachne dubia, Aristida mendocina, Gouinia paraguaiensis, Chloris ciliata, Trichloris pluriflora, Setaria hunzikeri, Setaria pampeana y Setaria lachnea) fue muy baja: <1.5 plantas/m2 durante el primer año y <1 planta/m2 en los años siguientes. En consecuencia, la densidad total de plantas de gramíneas forrajeras (agregadas + no agregadas) fue muy influenciada por la densidad de Pappophorum philippianum, y tendió a ser mayor en las parcelas con agregado de semillas que en las parcelas sin agregado (Tabla 2; Figura 3). La diferencia fue de 2 a 3 plantas/m2 durante los dos primeros años y de 1 plantas/m2 en los dos últimos. El poceado y la aplicación de mantillo no afectaron la densidad total de plantas de gramíneas forrajeras, que luego del año seco 2004/2005 se redujo de forma significativa en todos los tratamientos (-3 a -6 plantas/m2; Tabla 2; Figura 3). Ninguna de las interacciones evaluadas tuvo efecto sobre densidad total de plantas de gramíneas forrajeras (Tabla 2).
DISCUSIÓN
Este trabajo constituye el primer documento sobre la evaluación de técnicas orientadas directamente a la recuperación de los pastos nativos en la región del Chaco Árido argentino. El agregado de semillas fue el único tipo de intervención que permitió incrementar la densidad de plántulas en el corto plazo y que posibilitó duplicar la densidad de gramíneas forrajeras, pese a que (en términos absolutos) ese incremento fue muy pequeño. En algunos trabajos de rehabilitación realizados en ambientes semiáridos degradados de Sudáfrica (aproximadamente 600 mm de precipitación media anual) (Snyman 2003) y de Australia (Cole et al. 2005), la siembra de gramíneas también permitió elevar la densidad de plantas en el corto plazo.
El pulso de emergencia de plántulas tuvo lugar el primer verano, y fue bastante mayor en las parcelas con agregado de semillas. Sin embargo, también se observaron algunas plántulas en las parcelas que sólo fueron poceadas y/o cubiertas con mantillo (Figura 2). Tal vez la aplicación de mantillo, y en especial el poceado, crearon micrositios que actuaron como trampas para las semillas que fueron arrastradas (probablemente desde parcelas donde se agregaron semillas) por acción del viento (Marone et al. 1998a). Es poco factible que el poceado haya actuado removiendo y subiendo a la superficie semillas que se encontraban enterradas en el suelo (>2 cm de profundidad), y por lo tanto con poca posibilidad de emergencia, ya que las especies agregadas forman bancos de semillas superficiales (<2 cm de profundidad), con dormición escasa o nula (las viables pueden germinar apenas diseminadas) y de persistencia limitada en el tiempo [banco de semillas "transitorio" (Marone et al. 1998a, 2000; De Souza Maia et al. 2006)]. En estudios realizados en un pastizal de características similares y con especies de pastos C4 entre las que se encuentran las agregadas en nuestro ensayo, Marone et al. (1998b, 2000) encontraron que la granivoría por parte de aves y pequeños mamíferos consume por año el 50% de las semillas del banco del suelo, concentrándose en los meses de otoño e invierno. Ello sugiere que en nuestro estudio, la granivoría podría haber diezmado la reserva de semillas del suelo luego del primer año, y restringido la emergencia de plántulas a partir del segundo año.
Como se demostró en otras regiones semiáridas del mundo, nuestros resultados sugieren que las condiciones ambientales (en especial las precipitaciones), ejercen un control muy importante sobre la dinámica de la vegetación (Holmgren et al. 2001), y sobre el éxito de los tratamientos de rehabilitación (Snyman 2003; van den Berg & Kellner 2005). Se ha descrito el rol de años lluviosos y años secos sobre la dinámica de las comunidades vegetales (Holmgren et al. 2001) y se ha demostrado el efecto "residual" (que persiste de un año al siguiente) de las precipitaciones sobre la productividad primaria. Un año seco puede modificar las características de una comunidad vegetal a través de disminuir la producción de semillas, la emergencia y el establecimiento de plántulas, la supervivencia de plantas adultas, el desarrollo de área foliar y/o el macollaje de los pastos. Esto restringe la productividad de la comunidad en la estación de crecimiento siguiente (Villalba & Veblen 1998; Oesterheld et al. 2001). Inclusive, un año de escasas precipitaciones puede producir cambios persistentes en la composición florística de la comunidad ya que las distintas especies existentes pueden presentar distinta respuesta a periodos sequía (Holmgren & Scheffer 2001).
En nuestro estudio, las bajas precipitaciones durante la estación de crecimiento 2004/2005 redujeron a un tercio (o menos) la densidad de gramíneas forrajeras. Sin embargo, nuestros resultados no deben ser extrapolados automáticamente a otras situaciones sin antes tener en cuenta la combinación particular de condiciones climáticas (Marone et al. 2000) que tuvieron lugar durante este ensayo: un gran evento de lluvia a un mes del inicio, y un tercer año notablemente seco. Por otra parte, los tratamientos se aplicaron al final de enero, y la primera lluvia importante sucedió recién un mes más tarde. Ello determinó que el pulso de germinación de las semillas se diera tarde en la estación de crecimiento (que se extiende entre noviembre y marzo). Esto podría haber influido en que gran parte de las plántulas emergidas no alcanzaran tamaño suficiente para afrontar las heladas y la ausencia de lluvias del invierno (Petruzzi et al. 2003). En observaciones realizadas durante el invierno 2003 se pudo ver la alta tasa de mortalidad de plántulas emergidas meses antes (observación personal). El hecho de que las plántulas censadas en junio 2003 no hayan vuelto a ser censadas o no hayan llegado al estado de plantas al año siguiente es consistente con esa observación. Posiblemente, una siembra más temprana (noviembre o diciembre) habría permitido una emergencia temprana, con lo cual las plántulas habrían alcanzado un mayor tamaño antes del período crítico invernal.
El corte de ramas para creación de mantillo, el pasaje de un rolo para hacer pozos en el terreno, e incluso las combinaciones de las técnicas evaluadas, no se tradujeron en ningún aumento de la densidad de plantas de pastos forrajeros. Por el contrario, en otros ambientes semiáridos degradados del mundo se encontró que la remoción de suelo con arados, escarificadores o palas de mano (Snyman 2003; Cole et al. 2005; van den Berg & Kellner 2005) y la aplicación de una generosa cobertura de ramas sobre el suelo [2000-4000 g/m2 (Zuleta et al. 2003)] permitieron aumentar la densidad y/o cobertura de la vegetación en el corto plazo. Es probable que la intensidad del poceado que utilizamos en nuestro experimento haya sido demasiado conservativa, ya que el pasaje del rolo sólo removió un 4% de la superficie de las parcelas (8 pozos/m2 y un diámetro de pozo de 0.08 m (8 pozos/ m2 x 0.005 m2/ pozo x 100 = 4%). Tal vez, un movimiento de suelo mayor habría favorecido una mayor infiltración del agua de lluvia, un mejor contacto entre las semillas y el suelo, y un mejor desarrollo del sistema radicular de las plántulas.
La aplicación de cantidades de mantillo relativamente bajas podría haber sido insuficiente para mejorar las condiciones microambientales necesarias para el establecimiento de nuevas plantas (Fowler 1986). En ambientes áridos, esto podría implicar poco cambio de temperatura y del estado hídrico de la superficie del suelo y de las plántulas, factores involucrados en su supervivencia (Fowler 1986; Xiong & Nilsson 1999). Entre las especies que usamos como mantillo estuvieron dos congéneres de Larrea tridentata (L. divaricada y L. cuneifolia), especie del sur-oeste de Norteamérica que posee sustancias alelopáticas en sus hojas (fenoles y ácidos) que pueden ser trasladadas al suelo por la lluvia (Hyder et al. 2002). Sin embargo, en nuestro estudio no observamos ningún efecto negativo del mantillo sobre la cantidad de plántulas (Tabla 1) o de plantas (Tabla 2). Por esto deducimos que el mantillo usado no ha influido negativamente sobre los pastos.
Las diferencias en densidad de plantas entre parcelas con y sin agregado de semillas se explican principalmente por la presencia de plantas de Pappophorum philippianum, pese a que su aporte en la mezcla de semillas agregada fue la mitad que el aporte de las otras especies (excepto en el caso de su congénere P. caespitosum). En cambio, el agregado de Pappophorum caespitosum, Digitaria californica y Trichloris crinita no contribuyó a incrementar la densidad de plantas forrajeras. Esto sugeriría una mayor adaptación de Pappophorum philippianum para establecerse en esta zona (McKay et al. 2005). Tal aptitud ya había sido observada en las mediciones realizadas antes de iniciar este experimento, cuando se vio que esta especie mostró una densidad mayor respecto de las demás forrajeras. Por otro lado, P. philippianum es la especie de menor valor forrajero del conjunto agregado (Anderson et al. 1980). Esto muestra el compromiso que puede existir al momento de seleccionar materiales para incrementar la productividad forrajera en ambientes áridos, ya que las especies de mayor valor forrajero, por tener una mayor proporción de hojas y tejidos más tiernos, pueden no ser las mejor adaptadas para la supervivencia (Stebbins 1981).
Nuestros resultados, obtenidos en un sitio sobrepastoreado, brindan información complementaria a la de Passera et al. (1992) en un sitio clausurado al ganado por 20 años. La evaluación conjunta de los resultados obtenidos en ambos trabajos sugiere que el sobrepastoreo podría cambiar el orden de importancia relativo de las limitantes al incremento de la productividad de las gramíneas (Wisley & Polley 2003). En el sitio clausurado al pastoreo durante 20 años se manifestaría en mayor medida el efecto de competencia de los arbustos sobre los pastos (Reader & Best 1989; Goldgerg & Novoplansky 1997). Es esperable, además, que el banco de semillas de pastos forrajeros sea más abundante debido a la presencia de una cantidad mayor de plantas adultas bien desarrolladas y capaces de semillar (O´Connor & Pickett 1992; Marone et al. 2000; Haretche & Rodríguez 2006). Eso explicaría que Passera et al. (1992) hayan encontrado mayor respuesta a la remoción de arbustos que al agregado de semillas. Por el contrario, en el sitio sobrepastoreado se podría esperar que el efecto de competencia de los arbustos sea más leve, e incluso que cobren más fuerza las relaciones de facilitación de los arbustos sobre los pastos [protección contra el pisoteo y consumo animal, existencia de islas de fertilidad y mayor humedad en el suelo, resguardo de las plántulas contra condiciones ambientales extremas (Reader & Best 1989; Holmgren & Scheffer 2001)]. Así mismo, se podría esperar que en sitios degradados por sobrepastoreo, el banco de semillas de pastos forrajeros sea menos abundante (Bertiller 1992; O´Connor & Pickett 1992; Haretche & Rodríguez 2006). En este marco, nuestro trabajo sugiere que el agregado de semillas podría ser una intervención clave en los intentos para recuperar la capacidad forrajera en los ambientes sobrepastoreados del Chaco Árido.
AGRADECIMIENTOS
Este trabajo fue financiado por INTA La Rioja. Los autores quieren agradecer a: M. Almada, G. Berone, T. Flores, Don Andrada, E. Villagrán y D. Recalde por su colaboración en la instalación de la clausura y en la aplicación de los tratamientos; S. Cuello, E. Ávila y R. Ávila por su colaboración en distintas etapas de esta experiencia; la familia Mercado Luna por facilitar un espacio de su campo para realizar el ensayo; R. Golluscio y dos revisores anónimos por aportar valiosos comentarios para mejorar este manuscrito.
BIBLIOGRAFÍA
ARONSON, J; C FLORET; E LEFLOC`H; C OVALLE & R PONTANIER. 1993. Restoration and rehabilitation of degraded ecosystems in arid and semi-arid lands. I. A view from the south. Restoration Ecology 1: 8-17. [ Links ]
ANDERSON, DL; JA DEL AGUILA; A MARCHI; JC VERA; EL ORIONTE ET AL. 1980. Manejo del pastizal natural y producción ganadera. Parte 1. Páginas 1-61. Editorial INTA. Buenos Aires, Argentina. [ Links ]
BERTILLER, MB. 1992. Seasonal variation in the seed bank of a Patagonian grassland in relation to grazing and topography. Journal of Vegetation Science 3:47-54. [ Links ]
BIURRUN, FN. 1988. La Región de Los Llanos. El deterioro del Ambiente en la Argentina. Fundación para la Educación, la Ciencia y la Cultura. Buenos Aires, Argentina. [ Links ]
BLANCO, LJ; CA FERRANDO; FN BIURRUN; EL ORIONTE; P NAMUR ET AL. 2005. Vegetation responses to roller chopping and buffelgrass seeding in Argentina. Rangeland Ecology and Management 58:219-224. [ Links ]
BLANCO, LJ; MO AGUILERA; FN BIURRUN & JM PARUELO. 2008. Grazing effect on NDVI across an aridity gradient in Argentina. Journal of Arid Environments 72:764-776. [ Links ]
CABIDO, M; C GONZÁLEZ; A ACOSTA & S DÍAZ. 1993. Vegetation changes along a precipitation gradient in Central Argentina. Vegetatio 109:5-14. [ Links ]
CALL, CA & BA ROUNDY. 1991. Perspectives and processes in revegetation of arid and semiarid rangelands. Journal of Range Management 44: 543- 549. [ Links ]
COLE, I; ID LUNT & T KOEN. 2005. Effects of sowing treatment and landscape position on establishment of the perennial tussock grass Themeda triandra (Poaceae) in degraded eucalyptus woodlands in southeastern Australia. Restoration Ecology 13: 552-561. [ Links ]
DAUBENMIRE, R. 1959. A canopy-coverage method of vegetation analysis. Northwest Science 33:43–65. [ Links ]
DE SOUZA MAIA, M; FC MAIA & MA PÉREZ. 2006. Bancos de semillas en el suelo. Agriscientia 23: 33- 44. [ Links ]
DIEZ, HA; HF CALELLA; JC PEREZ; G PIGNATA; G BRONSTEIN ET AL. 1991. Acuerdo federal para el desarrollo rural del Chaco Árido. Documento Proyecto. Consejo Federal de Inversiones. [ Links ]
FOWLER, NL. 1986. Microsite requeriments for germination and establishment of three grass especies. American Midland Naturalist 115: 131- 145. [ Links ]
GOLDBERG, D & A NOVOPLANSKY. 1997. On the relative importance of competition in unproductive environments. The Journal of Ecology 85:4.9-418. [ Links ]
GÓMEZ, JC; HP CALELLA; RR CORZO & AA REYNOSO. 1993. Mapa de las subregiones de vegetación y suelo del Chaco Árido de La Rioja. Proyecto de Desarrollo Rural Integral de Los Llanos. Consejo Federal de Inversiones. [ Links ]
GONZÁLEZ, CL & JD DODD. 1979. Production responses of native and introduced grasses to mechanical brush manipulation, seeding, and fertilization. Journal of Range Management 32: 305- 309. [ Links ]
HANDA, IT & RL JEFFERIES. 2000. Assisted revegetation trials in degraded salt-marshes of the Hudson Bay lowlands. Journal of Applied Ecology 37:944-958. [ Links ]
HANDA, IT; R HARMSEN & RL JEFFERIES. 2002. Patterns of vegetation change and the recovery potential of degraded areas in a coastal marsh system of the Hudson Bay lowlands. Journal of Ecology 90: 86-99. [ Links ]
HARETCHE, F & C RODRÍGUEZ. 2006. Banco de semillas de un pastizal uruguayo bajo diferentes condiciones de pastoreo. Ecología Austral 16: 105- 113. [ Links ]
HOLMGREN, M; M SCHEFFER; E EZCURRA; JR GUTIERREZ & GMJ MOHREN. 2001. El Niño effects on the dynamics of arid ecosystems. Trends in Ecology and Evolution 16:89-94. [ Links ]
HOLMGREN, M & M SCHEFFER. 2001. El Niño as a window of opportunity for the restoration of degraded arid ecosystems. Ecosystems 4:151-159. [ Links ]
HYDER, PW; EL FREDRICKSON; RE ESTELL & ME LUCERO. 2002. Transport of phenolic compounds from leaf surface of creosotebush and tarbush to soil surface by precipitation. Journal of Chemical Ecology 28:2475-2482. [ Links ]
LITTELL, RC; GA MILLIKEN; WW STROUP & RD WOLFINGER. 1996. SAS for mixed models. SAS Institute, Cary, NC, USA. [ Links ]
MAESTRE, FT; S BAUTISTA; J CORTINA & J BELLOT. 2001. Potential for using facilitation by grasses to establish shrubs on a semiarid degraded steppe. Ecological Applications 11:1641-1655. [ Links ]
MARONE, L; BE ROSSI & ME HORNO. 1998a.Timing and spatial patterning of seed dispersal and redistribution in a South American warm desert. Plant Ecology 137:143-150. [ Links ]
MARONE, L; BE ROSSI & J LÓPEZ DE CASENAVE. 1998b. Granivore impact on soil-seed reserves in the central Monte desert, Argentina. Functional Ecology 12:640-645. [ Links ]
MARONE, L; ME HORNO & R GONZÁLEZ DEL SOLAR. 2000. Post-dispersal fate of seeds in the Monte desert of Argentina: patterns of germination in successive wet and dry years. Journal of Ecology 88:940-949. [ Links ]
MCKAY, JK; CE CHRISTIAN; S HARRISON & K RICE. 2005. "How local is local?" A review of practical and conceptual issues in the genetics of restoration. Restoration Ecology 13:432-440. [ Links ]
MORELLO, JH; J PROTOMASTRO; LA SANCHOLUZ & C BLANCO. 1985. Estudio macroecológico de Los Llanos de La Rioja. Serie del cincuentenario de la Administración de Parques Nacionales 5:1-53. [ Links ]
O´CONNOR, TG & GA PICKETT. 1992. The influence of grazing on seed production and seed banks of some African savannas grasslands. Journal of Applied Ecology 29:247-260. [ Links ]
OESTERHELD, M; J LORETI; M SEMMARTIN & OE SALA. 2001. Inter-annual variation in primary production of a semi-arid grassland related to previous-year production. Journal of Vegetation Science 12:137-142. [ Links ]
PASSERA, CB; O BORSETTO; RJ CANDIA & CR STASI. 1992. Shrub control and seeding influences on grazing capacity in Argentina. Journal of Range Management 45:480-482. [ Links ]
PETRUZZI, HJ; NP STRIZLER; EO ADEMA; CM FERRI & JP PAGELLA. 2003. Mijo Perenne. Publicación Técnica Nº 51 de la EEA Anguil. Ediciones INTA. [ Links ]
PROHASCA, FJ. 1959. El polo de calor de América del Sur. IDIA 141:27-30. [ Links ]
RAGONESE, AE & JC CASTIGLIONI. 1970. La vegetación del Parque Chaqueño. Sociedad Argentina de Botánica 11:133-160. [ Links ]
READER, RJ & BJ BEST. 1989. Variation in competition along an environmental gradient: Hieracium floribundum in an Abandoned Pasture. Journal of Ecology 77:673-684. [ Links ]
SAS INSTITUTE. 1996. SAS/STAT software: changes and enhancements through Release 6.11. SAS Institute, Cary, North Carolina, USA. [ Links ]
SNYMAN, HA. 2003. Revegetation of bare patches in a semi-arid rangeland of South Africa: an evaluation of various techniques. Journal of Arid Environments 55:417-432. [ Links ]
STEBBINS, GL. 1981. Coevolution of grasses and herbivores. Annals of the Missouri Botanical Garden 68:75-86. [ Links ]
SUDING, KN; KL GROSS & GR HOUSEMAN. 2004. Alternative states and positive feedbacks in restoration ecology. Trends in Ecology and Evolution 19:46-53. [ Links ]
VAN DEN BERG, L & K KELNER. 2005. Restoring degraded patches in a semi-arid rangeland of South Africa. Journal of Arid Environments 61: 497-511. [ Links ]
VILLALBA, R & TT VEBLEN. 1998. Influences of large-scale climatic variability on episodic tree mortality in northern Patagonia. Ecology 79:2624-2640. [ Links ]
WESTOBY, M; B WALKER & I NOY-MEIR. 1989. Opportunistic management for rangelands not at equilibrium. Journal of Range Management 42: 266-274. [ Links ]
WISLEY, BJ & HW POLLEY. 2001. Effects of seed additions and grazing history on diversity and productivity of subhumid grasslands. Ecology 84:920-931. [ Links ]
XIONG, S & C NILSSON. 1999. The effects of plant litter on vegetation: a meta-analysis. Journal of Ecology 87:984-994. [ Links ]
ZULETA, GA; MC LI PUMA & A BUSTAMANTE LEIVA. 2003. Initial effects of branching designs to restore semiarid shrub steppes in abandoned oil and gas locations of Patagonia, Argentina. 15th Annual International Conference of the Society for Ecological Restoration, Austin, Texas, USA. [ Links ]

