










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.19 n.2 Córdoba mayo/ago. 2009
TRABAJOS ORIGINALES
Patrón espacial de la regeneración de dos especies leñosas en un sector aprovechado de la Selva Paranaense
Martín A Pinazo*, Fabio A Moscovich, Claudio Dummel & Otto E Knebel
INTA, EEA Montecarlo, Misiones. Argentina.
INTA, EEA Montecarlo, Av. Libertador 2472, (3384) Montecarlo, Misiones, Argentina. Email: mpinazo@montecarlo.inta.gov.ar
Recibido: 5 de diciembre de 2008;
Fin de arbitraje: 3 de mayo de 2009;
Revisión recibida: 2 de julio de 2009;
2da. Revisión recibida: 16 de julio de 2009;
Aceptado: 17 de julio de 2009
RESUMEN. Los patrones de regeneración de las especies vegetales determinan la estructura de la comunidad madura y brindan información que posibilita el aprovechamiento racional de las especies más valiosas. En este trabajo se analizó el patrón espacial de la regeneración de Cabralea canjerana (Vell.) Mart. y Balfourodendron riedelianum (Eng.) Eng. en un área aprovechada de la Selva Paranaense en la provincia de Misiones, Argentina. De manera específica se evaluó la aparición eventual de agrupamientos en la regeneración, la manifestación de patrones atribuibles a potenciales procesos denso-dependientes en relación a los individuos conespecíficos adultos y patrones compatibles con la existencia de procesos de facilitación por parte de individuos adultos del resto de las especies. Los renovales de ambas especies presentaron patrones agregados hasta los 15 m. La regeneración y los individuos adultos conespecíficos de C. canjerana mostraron relaciones positivas a distancias entre los 5 y 10 m, mientras que la regeneración de B. riedelianum presentó un patrón al azar a corta distancia y asociación positiva a partir de los 5 m respecto a los individuos conespecíficos adultos. La relación de los renovales respecto a los individuos adultos de todas las especies presentó relaciones positivas en C. canjerana hasta los 10 m, mientras que B. riedelianum se asoció a partir de los 10 m de distancia. Los patrones de la regeneración de C. canjerana podrían ser atribuidos a los mecanismos de dispersión primaria y secundaria de la especie. En el caso de B. riedelianum, los patrones encontrados podrían originarse por la limitación en la dispersión de las semillas, lo cual pone en evidencia su dependencia de la presencia de individuos adultos para la regeneración.
Palabras clave: Distribución espacial; Índice O; Dispersión de semillas; Cabralea canjerana; Balfourodendron riedelianum.
ABSTRACT. Spatial pattern of regeneration of two woody species in a logged area in Paranaense Forest: The regeneration patterns of plants determine the structure of mature communities and provide usefull information to ensure the rational use of valuables species. In this study, the spatial regeneration pattern of Cabralea canjerana (Vell.) Mart. and Balfourodendron riedelianum (Eng.) Eng. was studied in a remnant sector of Paranaense Forest in Misiones province, Argentine. We evaluated the regeneration clustering, also the related patterns with conspecific adults trees potentially linked to density dependent process; and the existence of facilitation process related to the adult trees from all species. All trees were mapped in 4 replications of 100 x 100 m plots. Univariate and bivariate point pattern analysis were carried out using Wiegand & Moloney "O" ring index. Both species have showed a regeneration in aggregated pattern for distances up to 15 m. A positive relationship was verified for distances between 5 and 10 m for C. canjerana regeneration and conespecific adult trees; and the regeneration of B. riedelianum showed a random pattern in short distances (<5 m) and a positive association for larger distances of 10 m with their conspecific adult trees. The relationship between C. canjerana regeneration and adult trees from all species was positive at distances of 10 m, whereas B. riedelianum showed association at 10 m onward. The observed regeneration patterns of C. canjerana could be due to the primary and secondary dispersal mechanism of the species. On the other side, the observed patterns of B. riedelianum may represent a seed dispersal limitation, but the regeneration depends on the presence of adult trees of this species.
Keywords: Spatial distribution; O ring index; Seed dispersal; Cabralea canjerana, Balfourodendron riedelianum.
INTRODUCCIÓN
El estudio de los patrones espaciales de plantas son un paso fundamental para comprender o para determinar el efecto de sucesos pasados sobre el patrón presente y poder generar hipótesis sobre los procesos biológicos o ambientales que los determinan (Dale 1999; Levine 1992). Los patrones espaciales son el resultado de procesos diferentes que operan en distintas escalas (Condit et al. 2000; Wiegand & Moloney 2004; Wiegand et al. 2007) y guardan relación con los síndromes de regeneración propios de las especies (e.g., modos de dispersión, tolerancia a la sombra, establecimiento, supervivencia y desarrollo) y con la heterogeneidad ambiental (Condit et al. 2000; Seidler & Plotkin 2006).
En líneas generales, la regeneración tiende a estar agrupada, mientras que los individuos mayores presentan una disminución del grado de agrupamiento a medida que aumenta el tamaño de los individuos (Batista & Maguire 1998; Condit et al. 2000). El agrupamiento de los individuos puede ser el resultado de limitaciones en la dispersión de semillas o de la heterogeneidad ambiental (Condit et al. 2000; Nathan & Muller-Landau 2000; Seidler & Plotkin 2006), aunque la existencia de mecanismos denso dependientes puede generar la disminución del grado de agregación de los individuos (Janzen 1970; Connell 1971; Condit et al. 2000). Las especies cuyas semillas son dispersadas por el viento tienden a presentar patrones agregados debido principalmente a limitaciones en la dispersión. Mientras tanto, las especies cuyas semillas son dispersadas por animales por lo general presentan patrones menos agrupados o regulares (Seidler & Plotkin 2006). Sin embargo, la dispersión por parte de algunos vertebrados puede generar patrones agrupados verificándose procesos de dispersión directa, especialmente en especies con frutos de alto valor energético (Wenny 2001). Por otra parte, las diferencias en la tolerancia a la sombra pueden determinar agrupamientos que coinciden con claros naturales, como en el caso de especies pioneras o demandantes de luz (Pearson et al. 2003), mientras que en el caso de especies tolerantes a la sombra se puede esperar que se den patrones al azar o agregados en función de las características particulares de las diferentes especies o de las situaciones ambientales (Franklin & Rey 2007).
Actualmente, la Selva Paranaense, como parte integral del denominado Bosque Atlántico, es uno de los ecosistemas más amenazados y forma parte de iniciativas de conservación (Olson & Dinerstein 2002). Hasta el presente, esta formación boscosa ha sido sometida a la extracción selectiva de los individuos de las especies comerciales mediante la técnica denominada "diámetro mínimo de corta", que supone que los individuos por debajo del diámetro permitido para la corta podrán desarrollarse y mantener el estado productivo del bosque (Matthews 1989; Lamprecht 1990). Sin embargo, los daños producidos durante la extracción sumados a tasas excesivas de corta y la falta de acciones complementarias de manejo, generan bosques degradados con una excesiva proliferación de cañas de las especies Chusquea ramosissima y Merostachys clausenii que, junto con la proliferación de lianas Adenocalymna sp. y Arrabidaea sp., impiden la regeneración de las especies arbóreas del bosque por períodos de tiempo largos (Campanello et al. 2007a; Campanello et al. 2007b). Balfourodendron riedelianum (Eng.) Eng. y Cabralea canjerana (Vell.) Mart. son especies de importancia económica de la Selva Paranaense, y por lo tanto están sometidas a una fuerte presión de corta.
En estos ambientes, los individuos remanentes juegan un rol fundamental sobre la lluvia de semillas, ya sea como perchas de dispersores o por el aporte de sus propias semillas (Rother et al. 2009), y también pueden actuar como facilitadores de la regeneración (Campanello et al. 2007b). Es por ello que la generación de conocimientos relativos a los patrones espaciales de las especies son de interés para comprender la dinámica de estos bosques, con el objeto de desarrollar técnicas de manejo que permitan su aprovechamiento racional (Batista & Maguire 1998; Fajardo et al. 2006). El objetivo del presente trabajo es examinar los patrones espaciales de la regeneración de dos especies comerciales y las relaciones de la regeneración con los individuos adultos conespecíficos y del resto de las especies. En particular, se pretende verificar la aparición de agrupamientos en la regeneración, la manifestación de patrones atribuibles a potenciales procesos denso dependientes en relación a los individuos conespecíficos adultos y patrones compatibles con la existencia de procesos de facilitación por parte de individuos adultos del resto de las especies.
MATERIALES Y MÉTODOS
Sitio y especies estudiados
El estudio se llevó a cabo en un sector remanente de 500 ha de bosque nativo en el Campo Anexo Manuel Belgrano, perteneciente al Instituto Nacional de Tecnología Agropecuaria (Misiones, Argentina) (26º03'20'' S; 53º48'08'' W). En la actualidad, este campo experimental constituye el único remanente de bosque nativo en el área junto con la reserva estricta San Antonio, con la que limita al norte. La agricultura y ganadería de pequeña escala son los usos predominantes en las áreas linderas. La vegetación predominante corresponde a la región Selva Paranaense, Selva de Laurel, Guatambú y Pino (Cabrera 1976). El bosque fue sometido al aprovechamiento selectivo durante los años 1950-1955 y hasta la actualidad no se verificaron nuevas extracciones ni prácticas silvícolas. Se encuentra dominado por cañas en los claros de corta, mientras que en los sectores con mayor cobertura la densidad de cañas disminuye (observación personal). La presencia de cañas y lianas es abundante, a pesar de que los valores de área basal son mayores a los comúnmente se hallan en sitios aprovechados (Tabla 1) y podría clasificarse como un área poco disturbada según los trabajos de Placci et al. (1992) y López-Cristóbal et al. (1996). La temperatura media anual del área es 21 ºC, con una amplitud media mensual de 10 ºC. La precipitación media anual es 2000 mm, distribuida de manera uniforme durante el año. En el invierno pueden darse heladas de manera poco frecuente (Cabrera 1976). Los suelos son lateríticos, derivados de rocas basálticas y corresponden a los Kandiudultes (USDA 2006).
Tabla 1. Parámetros estructurales de las parcelas estudiadas*. DM: diámetro promedio; D.Máx: diámetro máximo.
Table 1. Structural parameters of the study plots*. DM: mean DHB; D.Máx: maximun DHB

Cabralea canjerana (cancharana) presenta una distribución amplia y una elevada frecuencia en la región neotropical, y se extiende desde Costa Rica hasta el sur de Brasil y el norte de Argentina (Cabrera 1976; Pennington 1981). La cancharana es considerada como tolerante a la sombra durante su estadio juvenil (Carvalho 1994). Los individuos maduros alcanzan 20-30 m de altura y 150 cm de diámetro a la altura del pecho (Gartland & Parusini 1990). La maduración de los frutos tiene lugar entre septiembre y enero, y el pico de maduración sucede en noviembre-diciembre. Las semillas son dispersadas por un grupo diverso de especies de aves (Pizo 1997), y las hormigas cumplen un papel importante como dispersores secundarios y facilitadores de la germinación (Pizo & Oliveira 1998, 2001). Los frutos son capsulas globosas dehiscentes, con 1-6 diásporas a las que corresponden 1 ó 2 semillas envueltas en un arilo fino. El arilo de C. canjerana posee un contenido elevado de lípidos, lo cual la ubica dentro del grupo de plantas con frutos altamente energéticos (Pizo 1997; Pizo & Oliveira 2001) y convierte a las semillas en recursos muy atractivos para aves, roedores e insectos predadores. Además, se verifican niveles elevados de depredación después de la dispersión, por sobre procesos de depredación en la planta madre o de pre-dispersión (Pizo 1997).
Balfourodendron riedelianum (guatambú) está presente en Paraguay, Brasil y el noreste de Argentina, y es dominante del canopeo en la región (Cabrera 1976). Es considerada de hábito intermedio entre las categorías heliófilas y esciófilas y tolera sombra parcial durante su estadio juvenil (Carvalho 1994). Los individuos maduros alcanzan 20-25 m de altura y 80 cm de diámetro a la altura del pecho (Gartland & Parusini 1997). Los frutos son sámaras leñosas indehiscentes, con 3-4 alas semicirculares y de dispersión anemocórica (Eibl et al. 1990). La presencia de frutos maduros sucede desde abril hasta septiembre, con picos de producción en julio. Para ninguna de estas especies se dispone de información relacionada a la edad de fructificación y variaciones temporales en la producción de frutos.
Recolección de datos
Se establecieron cuatro parcelas de 100 m x 100 m en dos áreas de bosque nativo. En cada área se instalaron transectas cada 10 m para obtener una grilla de 10 m x 10 m. El sistema de grilla fue utilizado para realizar el mapeo de todos los individuos con diámetro a la altura del pecho (DAP) >10 cm y, además, todos los renovales de C. canjerana y B. riedelianum con DAP<10 cm y altura >1.3 m. Cada árbol fue ubicado por medio de una cinta métrica con una precisión de 10 cm, y luego fueron referidos a un sistema de coordenadas x-y. Todos los individuos con DAP>10 cm fueron identificados en el nivel de especie.
Análisis de datos
El análisis de los patrones espaciales de cada una de las especies se realizó por medio del estadístico "O" de Wiegand & Moloney, que se relaciona con la función K de Ripley (Wiegand & Moloney 2004). La función K de Ripley es el número esperado de puntos en un círculo de radio r y centro en un punto arbitrario, mientras que el estadístico "O" reemplaza a los círculos por anillos, con la ventaja de poder aislar los efectos en clases de distancia determinados. Mientras tanto, la función K de Ripley presenta efectos acumulativos que generan dificultades en la interpretación en los eventos a larga distancia con los eventos verificados a corta distancia (Condit et al. 2000; Wiegand & Moloney 2004). El cálculo del estadístico "O" implica la utilización de un anillo de radio r y ancho dr que es centrado en cada punto del patrón, en el que se cuentan el número de puntos vecinos que se encuentran dentro del mismo. Para n individuos distribuidos en un área A, la densidad (d=n/A) es el número promedio de individuos por unidad de área. La función O(r) es el número esperado de puntos dentro de un anillo de radio r centrado en un punto arbitrario del patrón.
El estadístico "O" es análogo a la función K de Ripley, y se define como:
O(r) = g(r)* λ
donde g(r) es:
g(r) = [dK(r)/dr]/(2.π.r)
donde K(r) es el valor de la función de Ripley definido como:
K(r) = λ-1 * E(r)
E: número esperado de puntos (e.g., árboles) a distancia r de un punto determinado (e.g., árboles). λ: intensidad del patrón de puntos.
El estadístico "O" para patrones bivariados, por ejemplo cuando se analiza la distribución de un evento (i.e., regeneración) respecto a otro evento puntual (i.e., árboles adultos conespecíficos), el estadístico se calcula mediante aproximación numérica como:
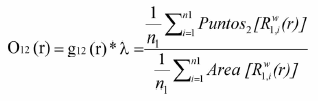
donde: n1: número total de puntos del patrón 1; R1,iw(r): anillo con radio r y ancho w centrado en el in punto del patrón 1; Puntos2: puntos del patrón 2 en la región definida por el anillo R1,iw(r). Área: determina el área de la región x. Si los puntos presentan una distribución de Poisson, la cual asume que los puntos están distribuidos al azar, el valor esperado de O(r) se iguala a la densidad o intensidad del patrón.
El patrón espacial de la regeneración de cada especie se realizó a través de análisis univariados, mientras que para explorar las relaciones entre la regeneración y árboles con DAP>10 cm conespecíficos y respecto a los individuos con DAP>10 cm de todas las especies, se realizaron análisis bivariados. Se estimaron los intervalos de confianza usando el método de simulación Monte Carlo mediante la realización de 99 simulaciones, representando los límites de confianza al 99% (Haase 1995). La realización de simulaciones Monte Carlo implica mantener la densidad de puntos del patrón y una distribución de los individuos en el espacio en función al modelo nulo utilizado (Haase 1995). Para el análisis univariado de C. canjerana se utilizó el modelo espacial completo al azar (CSR) como modelo nulo, el cual distribuye al azar los puntos del patrón. En el caso de B. riedelianum se optó por aplicar el método heterogéneo de Poisson debido a las diferencias en la densidad o intensidad de los puntos observados en las parcelas (ver información suplementaria en www.ecologiaaustral.com.ar). El modelo nulo se generó mediante la aplicación del método de ventana móvil (Wiegand & Moloney 2004). En los análisis bivariados se aplicó el modelo de condición antecedente como modelo nulo, el cual mantiene fijo el patrón (e.g., árboles adultos) contra el cual se desea analizar el patrón de interés (e.g., regeneración) que se distribuye al azar (Wiegand & Moloney 2004). Si el valor del estadístico "O" se mantiene dentro del intervalo de confianza generado mediante las simulaciones, se supone una distribución al azar para los análisis univariados, mientras que en los análisis bivariados esto significaría la ausencia de relación entre ambos patrones. Valores por encima del intervalo de confianza revela asociación en los análisis univariados, en cuanto que por debajo del intervalo de confianza indican segregación. Los análisis se realizaron con el software "Programita" (Wiegand & Moloney 2004), aplicando la metodología propuesta por Diggle (2003), que permite combinar diferentes parcelas para obtener los resultados estadísticos.
RESULTADOS
Los análisis espaciales univariados mostraron patrones agregados para la regeneración de ambas especies. C. canjerana presentó una agregación significativa a distancias de hasta 22 m, con un pico hasta los 10 m. En escalas mayores, la especie no presentó desviaciones significativas del modelo nulo propuesto (Figura 1). Por otra parte, B. riedelianum mostró agregación a distancias de hasta 15 m, con un pico hasta los 5 m (Figura 1).
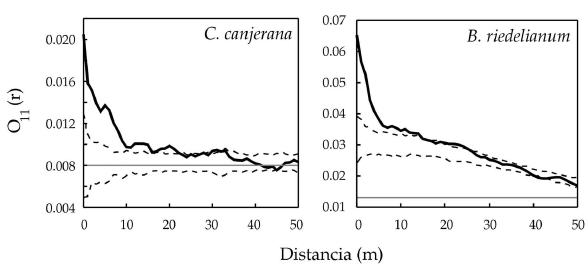
Figura 1. Análisis univariado de la regeneración de C. canjerana y B. riedelianum mediante el estadístico "O" de Wiegand & Moloney. En línea punteada limites de confianza al 99%; en línea gris "λ" o intensidad de puntos del área en estudio; en línea sólida negra valores del O11(r) para las diferentes escalas.
Figure 1. Univariate analysis of regeneration of C. canjerana, and B. riedelianum by means of the Wiegand & Moloney "O" index. Dashed lines are the 99% confidence limits; grey lines are "λ" or the point intensity of the study area and the solid lines are the O11(r) values at the different scales.
Los análisis bivariados de la regeneración y los individuos adultos conespecíficos (DAP>10 cm) presentaron diferentes patrones para ambas especies. La regeneración de C. canjerana mostró una tendencia a un patrón agregado a distancias hasta los 10 m, y otro pico entre los 25 a 35 m. Sin embargo, resultaron significativas las desviaciones alrededor de los 5 a 10 m en cercanías de los individuos conespecíficos (P<0.01), con otro pico significativo entre los 25-30 m (P<0.01) (Figura 2). La regeneración de B. riedelianum presentó tendencia a una asociación negativa (repulsión), pero no significativa, con los individuos conespecíficos adultos (DAP>10 cm) a distancias de hasta 5 m. A partir de los 5 m y hasta los 35 m aproximadamente, se verificó una relación positiva significativa (P<0.01) (Figura 2).
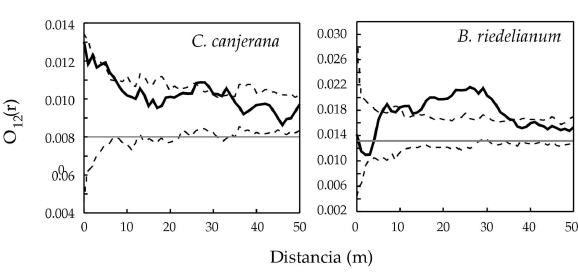
Figura 2. Análisis bivariado de la regeneración de C. canjerana y B. riedelianum e individuos conespecíficos con DAP>10 cm mediante el estadístico "O" de Wiegand & Moloney. En línea punteada limites de confianza al 99%; en línea gris "λ" o intensidad de puntos del área en estudio; en línea sólida negra valores del O12(r) para las diferentes escalas.
Figure 2. Bivariate analysis of regeneration of C. canjerana, and B. riedelianum and conspecific trees with DBH>10 cm by means of the Wiegand & Moloney "O" index. Dashed lines are the 99% confidence limits; grey lines are "λ" or the point intensity of the study area and the solid lines are the O12(r) values at the different scales.
Los análisis bivariados de la regeneración de ambas especies y los individuos de DAP>10 cm de todas las especies presentes resultaron en asociaciones positivas en ambos casos. La interacción de la regeneración de C. canjerana y los árboles adultos de todas las especies presentó una tendencia positiva en todas las escalas de análisis, con picos significativos hasta los 10 m y entre los 20 y 40 m (P<0.01) (Figura 3). Por su parte la regeneración de B. riedelianum mostró una asociación positiva a partir de los 15 m de distancia de los individuos adultos (P<0.01) (Figura 3).

Figura 3. Análisis bivariado de la regeneración de C. canjerana y B. riedelianum e individuos de todas las especies con DAP>10 cm mediante el estadístico "O" de Wiegand & Moloney. En línea punteada limites de confianza al 99%; en línea gris "λ" o intensidad de puntos del área en estudio; en línea sólida negra valores del O12(r) para las diferentes escalas.
Figure 3. Bivariate analysis of regeneration of C. canjerana, and B. riedelianum and trees of all species with DBH>10 cm by means of the Wiegand & Moloney "O" index. Dashed lines are the 99% confidence limits; grey lines are "λ" or the point intensity of the study area and the solid lines are the O12(r) values at the different scales.
DISCUSIÓN
Los patrones espaciales de la regeneración de ambas especies reflejarían principalmente los procesos de dispersión particulares a cada una de ellas y la posible manifestación de procesos denso dependientes y de facilitación en cada caso en particular. El patrón de la regeneración de B. riedelianum evidenciaría la limitación en la dispersión de sus semillas y la ocurrencia de potenciales procesos denso-dependientes en cercanías a los adultos de esta especie. Mientras tanto, el patrón espacial de la regeneración de C. canjerana resultó en una asociación significativa con los adultos conespecíficos como resultado de la acción de procesos de dispersión secundaria, y en posibles mecanismos de facilitación por parte de los individuos del resto de las especies. C. canjerana presenta dispersión por animales, principalmente aves, siendo esperable patrones al azar de acuerdo a lo expresado por Batista & Maguire (1998), Condit et al. (2000) y Seidler & Plotkin (2006). Sin embargo, las diferentes especies de aves que consumen los frutos presentan comportamientos de alimentación diferentes que pueden generar patrones diferentes al modelo nulo propuesto. Mientras que algunas especies consumen la semilla entera, que de esta manera debe pasar por el tracto digestivo, otras especies consumen solo el arilo sobre la misma planta, y otras -en cambio- se trasladan a diferentes plantas para consumirlo (Pizo 1997). Estos comportamientos particulares de las diferentes especies de aves involucradas en la dispersión pueden generar patrones agrupados cerca de los árboles madre y en los sitios preferidos utilizados como perchas (Wenny & Levey 1998; Wenny 2001). En este último proceso algunas especies son dispersores más eficientes, mientras que otras dejan caer varios frutos antes de poder consumirlos (Pizo 1997) y, por lo tanto, la mayoría de frutos y semillas caen o son depositados en las cercanías de las plantas madres. En esta situación se verifican procesos severos de predación por parte de insectos y roedores en conjunción con procesos de dispersión secundaria por partes de hormigas de los géneros Pachycondila, Odontomachus, Pheidole y Solenopsis entre otros (Pizo & Oliveira 1998, 2001).
Las hormigas cumplen un rol fundamental en el éxito del establecimiento de las plántulas, principalmente en cercanías de las plantas madres ya que facilitan la germinación, ocultan las semillas dificultando el acceso de predadores de las mismas y reducen la posibilidad de ataque de patógenos mediante la extracción del arilo de las mismas (Pizo & Oliveira 1998, 2001). En particular, los géneros Pachycondila y Odonthomachus pueden transportar semillas hasta 10 m (Passos & Oliveira 2002), distancia que coincide con el patrón de agrupamiento alrededor de los árboles padres. Estos procesos definirían los patrones de la regeneración de la especie ya que, por un lado la actividad de las hormigas facilitaría el establecimiento en las cercanías de las plantas madres y determinarían el pico de agregación alrededor de las mismas a distancias menores a los 10 m, mientras que los agrupamientos a mayores distancias estarían determinados principalmente por el comportamiento de las aves involucradas en la dispersión de las semillas (Wenny 2001; Alcántara et al. 2000; Wenny & Levey 1998; Pizo 1997). En este sentido, y por tratarse de un área aprovechada y representar un segmento de bosque, es esperable que la fauna involucrada en la dispersión haya sido impactada, alterando los procesos de dispersión de la especie (Jansen & Zuiderna 2001). En el caso particular de C. canjerana, se reportan alteraciones en la diversidad de aves involucradas en la dispersión de esta especie en sitios fragmentados (Pizo 1997). Este efecto de la fragmentación podría esperase en el área de estudio y por lo tanto la mayor abundancia de algún dispersor podría generar un patrón particular como el observado. Sin embargo, las aves dispersoras pueden presentar vuelos a mayores distancias y generar agrupamientos a distancias mayores a los 50 m que no sean detectados en el presente estudio. Por otra parte, la asociación con los adultos de todas las especies a distancias de hasta 10 m podría relacionarse al carácter tolerancia a la sombra de esta especie y a la dispersión por parte de aves que depositan las semillas bajo las copas de los árboles, mientras que la asociación verificada entre los 20 y 40 m podría deberse a la existencia de diferentes agrupamientos de la regeneración en el área de estudio.
B. riedelianum, especie de dispersión anemocórica cuyo patrón espacial resultó marcadamente más agregado respecto a C. canjerana en cortas distancias, reflejaría limitaciones en la dispersión de las semillas en concordancia con lo expresado por Seidler & Plotkin (2006) y Condit et al. (2000) para especies de árboles en bosques tropicales y a lo observado en Cedrela fissilis (Alcántara et al. 1997). La falta de tendencia en la cercanía de los individuos conespecíficos adultos podría deberse a la existencia de procesos denso-dependientes de predación de semillas, herbivoría y ataque de patógenos que limitaría la regeneración en la cercanía de los adultos (Janzen 1970; Connell 1971; Augspurger 1984; Wills et al. 1997). Sin embargo, este proceso se manifestaría a distancias pequeñas de menos de 5 m de los individuos conespecíficos. Este efecto denso-dependiente a escalas pequeñas, menores a 10 m, también fue documentado en otros bosques tropicales (Augspurger 1983; Condit et al. 1994; Blundell & Peart 1998). Por otra parte, la asociación positiva entre los 10 a 35 m reflejaría la limitación en la dispersión de las semillas a mayores distancias y explicaría la tendencia a regenerar alrededor de los árboles padres. La relación positiva con los individuos adultos de todas las especies refuerza la clasificación de Carvalho (1994) como especie tolerante a la sombra e indicaría limitaciones de la especie para regenerar en sitios abiertos o dominados por lianas y bambúes, donde es posible verificar una intensa competencia de estos con los renovales (Campanello et al. 2007a; Campanello et al. 2007b) y/o presentar condiciones ambientales estresantes para la especie (Campanello et al. 2008). Sin embargo, el agrupamiento a partir de los 15 m respecto a los adultos de todas las especies, y con un pico a los 30 m, podría significar preferencias a condiciones más iluminadas respecto a C. canjerana. En este sentido, Campanello (2004) menciona valores de hasta 30% de radiación como valores típicos de radiación donde se desarrolla la regeneración de B. riedelianum, mientras que Carvalho (1994) clasifica a esta especie como tolerante parcial de sombra durante los estadios juveniles.
A partir de los resultados obtenidos es posible realizar algunas recomendaciones de manejo de estas especies. Por un lado, el agrupamiento de la regeneración de C. canjerana en torno a los individuos adultos conespecíficos y del resto de las especies hasta distancias de 10 m, sugieren la factibilidad de realizar el manejo de esta especie mediante la liberación de la regeneración por medio de la corta de los adultos e individuos de otras especies a modo de tratamientos suplementarios, manteniendo una distancia entre árboles maduros remanentes no mayor a 60 m. Por otra parte, en el caso de B. riedelianum, que presentó agrupamientos a partir de los 10 m, la corta de individuos conespecíficos no aseguraría la liberación de los renuevos y por lo tanto se deberían considerar cortas de liberación suplementarias. De esta manera, se contribuiría a asegurar la presencia de renovales en toda el área en el caso de que estas especies constituyan el objetivo del manejo forestal. Finalmente, y en el caso de ambas especies, los sitios sometidos a disturbios como cortas excesivas podría condicionar el establecimiento de renovales y comprometer la producción en ciclos de corta futuras, tanto por la falta de individuos adultos, que aseguren la disponibilidad de semillas, como así también por la generación de situaciones que dificultan la regeneración de especies comerciales.
BIBLIOGRAFÍA
AUGSPURGER, CK. 1983. Seed dispersal of the tropical tree, Platypodium elegans, and the escape of its seedlings from fungal pathogens. J. Ecol. 71: 759-771. [ Links ]
AUGSPURGER, CK. 1984. Seedling survival of tropical tree species: Interactions of dispersal distance, light-gaps and pathogens. Ecology 65:1705-1712. [ Links ]
ALCÁNTARA, JM; PJ REY; F VALERA & AM SÁNCHEZ-LAFUENTE. 2000. Factors shaping the seedfall pattern of a bird-dispersed plant. Ecology 81: 1937- 1950. [ Links ]
ALCÁNTARA, JM; N VERA; L SZCZIPANSKI; B EIBL & R RÍOS. 1997. Análisis de la dispersión de semillas de Cedrela fissilis en el bosque nativo de Misiones. Yvyrareta 8:16-21. [ Links ]
BATISTA, JLF & DA MAGUIRE. 1998. Modelling the spatial structure of tropical forest. For. Ecol. Manage.110:293-314. [ Links ]
BLUNDELL, AG & DR PEART. 1998. Distance-dependence in herbivory and foliar condition for juvenile Shorea trees in Bornean dipterocarp rain forest. Oecologia 117:151-160. [ Links ]
CABRERA, AL. 1976. Regiones Fitogeográficas Argentinas. Enciclopedia Argentina de Agricultura y Jardineria. Editorial ACME. Capital Federal. Buenos Aires. Argentina. 85 Pp. [ Links ]
CAMPANELLO, PI. 2004. Diversidad, crecimiento y fisiología de árboles en la Selva Misionera: efectos de los cambios en la estructura y funcionamiento del ecosistema producidos por la invasión de lianas y bambúseas. Tesis doctoral. Universidad Nacional de Buenos Aires. [ Links ]
CAMPANELLO, P; JF GARIBALDI; MG GATTI & G GOLDSTEIN. 2007a. Lianas in a subtropical Atlantic Forest: Host preference a tree growth. For. Ecol. Manage. 242:250-259. [ Links ]
CAMPANELLO, PI; MG GATTI; A ARES; L MONTTI & G GOLDSTEIN. 2007b. Tree regeneration and microclimate in a liana and bamboo-dominated semideciduous Atlantic Forest. For. Ecol. Manage. 252:108-117. [ Links ]
CAMPANELLO, PI; MG GATTI & G GOLDSTEIN. 2008. Coordination between water-transport efficiency and photosynthetic capacity in canopy tree species at different growth irradiances. Tree Physiol. 28: 85- 94. [ Links ]
CARVALHO, PER. 1994. Espécies florestais brasileiras: recomendações silviculturais, potencialidades e uso da madeira. Empresa Brasileira de Pesquisa Agropecuária; Centro Nacional de Pesquisa de Florestas - Colombo: EMBRAPA - CNPF; Brasília. Republica Federativa del Brasil. 640 Pp. [ Links ]
CONDIT, R; PS ASHTON; P BAKER; S BUNYAVEJCHEWIN; S GUNATILLEKE ET AL. 2000. Spatial pattern in the distribution of tropical tree species. Science 288: 1414-1418. [ Links ]
CONDIT, R; SP HUBBELL & RB FOSTER. 1994. Density dependence in two in a neotropical forest understory tree species. Ecology 75:671-680. [ Links ]
CONNELL, JH. 1971. On the role of natural enemies in preventing competitive exclusion in some marine animals and rain forest trees. Pp 298-312 en Den Boer, PJ & Gradwell (eds.). Dynamics of populations. Pudoc, Wageningen. Holanda. [ Links ]
DALE, MRT. 1999. Spatial pattern analysis in plant ecology. Third edition. Cambridge University Press. Cambridge. Inglaterra. 326 Pp. [ Links ]
DIGGLE, PJ. 2003. Statistical analysis of spatial point pattern. Second edition. Academic Press. Londres. Inglaterra. 168 Pp. [ Links ]
EIBL, BI; F SILVA & G OTTENWELLER. 1990. Caracterización bioclimática del Guatambú Blanco (Balfourodendron riedelianum (Engl.) Engl.) Yvyrareta 1:53-65. [ Links ]
FAJARDO, A; JM GOODBURN & J GRAHAM. 2006. Spatial pattern of regeneration in managed uneven-aged Ponderosa pine/Douglas-fir forests of Western Montana, USA. For. Ecol. Manage. 223:255-266. [ Links ]
FRANKLIN, J & SJ REY. 2007. Spatial patterns of tropical forest trees in Western Polynesia suggest recruitment limitations during secondary succession. J. Trop. Ecol. 23:1-12. [ Links ]
GARTLAND, HM & MG PARUSINI. 1990. Caracterización dendrométrica de treinta especies forestales de Misiones. Yvyrareta 1:5-28. [ Links ]
HAASE, P. 1995. Spatial pattern analysis in ecology based on Ripley's K-function: Introduction and methods of edge correction. J. Veg. Sci. 6: 575- 582. [ Links ]
JANZEN, DH. 1970. Herbivores and the number of tree species in tropical forest. Am. Nat. 104: 501- 528. [ Links ]
JANSEN, PA & PA ZUIDERNA. 2001. Logging, seed dispersal by vertebrates, and natural regeneration of tropical timber trees. Pp. 35-59 en Fimbel, RA; A Grajal & JG Robinson (eds.). The cutting edge: Conserving wildlife in logged tropical forest. Columbia University Press. New York. USA. [ Links ]
LAMPRECHT, H. 1990. Silvicultura en los Trópicos. Agencia Alemana de Cooperación Técnica (GTZ). Eschborn. Alemania. 335 Pp. [ Links ]
LEVINE, SA. 1992. The problem of pattern and scale in ecology. Ecology 73:1943-1967. [ Links ]
LÓPEZ CRISTÓBAL, L; L GRANCE; D MAIOCCO & B EIBL. 1996. Estructura y composición florística del bosque nativo en el predio Guaraní. Yvyrareta 7:30-37. [ Links ]
MATTHEWS, JD. 1989. Silvicultural Systems. Oxford Science Publications. Oxford University Press. Oxford. Inglaterra. 284 Pp. [ Links ]
NATHAN, R & HC MULLER-LANDAU. 2000. Spatial pattern of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 15:277-285. [ Links ]
OLSON, DM & E DINERSTEIN. 2002. The global 200: Priority ecoregions for global conservation. Anna. Missouri Bot. Gard. 89:129-224. [ Links ]
PASSOS, L & PS OLIVEIRA. 2002. Ants affect the distribution and performance of seedlings of Clusia criuva, a primarily bird-dispersed rain forest tree. J. Ecol. 90:517-528. [ Links ]
PEARSON, TRH; DFRP BURSLEM; RE GOERIZ & JW DALLING. 2003. Interactions of gap size and herbivory on establishment, growth and survival of three species of neotropical pioneer trees. J. Ecol. 91:785-796. [ Links ]
PENNIGTON, TD. 1981. Meliaceae. Flor. Neotr. Mono. 28:234-242. [ Links ]
PIZO, MA. 1997. Seed dispersal and predation in two populations of Cabralea canjerana (Meliaceae) in the Atlantic Forest of southeastern Brazil. J. Trop. Ecol. 13:559-578. [ Links ]
PIZO, MA & PS OLIVEIRA. 1998. Interaction between ants and seeds of a nomyrmecochorus neotropical tree, Cabralea canjerana (Meliaceae), in the Altlantic Forest of Southeast Brazil. Am. J. Bot. 85:669-674. [ Links ]
PIZO, MA & PS OLIVEIRA. 2001. Size and lipid content of nonmyrmecochorous diaspores: effects on the interaction with litter-foraging ants in the Atlantic rain forest of Brazil. Plant Ecol. 157:37-52. [ Links ]
PLACCI, G; S ARDITI; P GIORGIS & A WUTHRICH. 1992. Estructura del palmital e importancia de Euterpe edulis como especie clave en el Parque Nacional Iguazú. Yvyrareta 3:93-108. [ Links ]
ROTHER, DC; RR RODRIGUES & MA PIZO. 2009. Effects of bamboo stands on seed rain limitation in a rainforest. For. Ecol. Manage. 257:885-892. [ Links ]
SEIDLER, TG & JB PLOTKIN. 2006. Seed dispersal and spatial pattern in tropical trees. Plos Biol. 4: 2132- 2137. [ Links ]
USDA. 2006. Keys to Soil Taxonomy. United States Department of Agriculture. Décima edición. Washington, DC. EEUU. 341 Pp. [ Links ]
WENNY, DG & DJ LEVEY. 1998. Directed seed dispersal by bellbirds in a tropical cloud forest. Proc. Natl. Acad. Sci. USA. 95:6204-6207. [ Links ]
WENNY, DG. 2001. Advantages of seed dispersal: A re-evaluation of directed dispersal. Evol. Ecol. Res. 3:51-74. [ Links ]
WIEGAND, T & KA MOLONEY. 2004. Rings, circles, and null models for point pattern analysis in ecology. Oikos 104:209-229. [ Links ]
WIEGAND, T; S GUNATILLEKE & N GUNATILLEKE. 2007. Species associations in a heterogeneous Sri Lankan Dipterocarp forest. Am.Nat. 170:77-95. [ Links ]
WILLS, CH; R CONDIT; RB FOSTER & SP HUBBELL. 1997. Strong density and diversity related effects help to maintain tree species diversity in a neotropical forest. Proc. Natl. Acad. Sci. USA 94:1252-1257. [ Links ]

