










Services on Demand
Journal

Article
 Portuguese (pdf)
Portuguese (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.19 no.3 Córdoba Sept./Dec. 2009
ARTÍCULO ORIGINAL
Esclerofilia, taninos e insetos herbívoros associados a Copaifera lagsdorffii Desf. (Fabaceae: Caesalpinioideae) em área de transição Cerrado-Caatinga no Brasil
Jhonathan O Silva1, *, Fabiene M Jesus2, Marcílio Fagundes2 & G Wilson Fernandes3
1. Ecologia Evolutiva/DBG, CCBS/ Universidade Estadual de Montes Claros, Caixa Montes Claros-MG, Brasil.
2. Biologia da Conservação/ DBG, CCBS/ Universidade Estadual de Montes Claros, Montes Claros-MG, Brasil.
3. Ecologia Evolutiva & Biodiversidade/DBG, ICB/Universidade Federal de Minas Gerais, Belo Horizonte MG, Brasil.
*Ecologia Evolutiva/DBG, CCBS/ Universidade Estadual de Montes Claros, Caixa Postal 126, 39401-089. Montes Claros-MG, Brasil. Email: jhonathanos@gmail.com
Recibido: 7 de abril de 2009;
Fin de arbitraje: 25 de junio de 2009;
Revisión recibida: 11 de agosto de 2009;
Aceptado: 22 de septiembre de 2009
RESUMEN. A riqueza e abundância de herbívoros associados a uma planta hospedeira são influenciadas por variações nas defesas físicas e químicas da planta as quais são mediadas pela qualidade do habitat. Neste estudo, 36 indivíduos de Copaifera langsdorffii distribuídos equitativamente entre habitats de Cerrado "sensu stricto", Matas Ciliares e Floresta Estacional Decidual foram usadas para avaliar os efeitos do habitat na esclerofilia foliar, produção de taninos e na fauna de insetos herbívoros associados. Observouse maior riqueza e abundância de herbívoros mastigadores em plantas dos habitats de Mata Ciliar e Cerrado comparativamente à Floresta Estacional Decidual. Porém, a riqueza e abundância de insetos herbívoros sugadores não diferiu entre os habitats. Não foi encontrada relação entre a riqueza ou abundância de herbívoros com a concentração de taninos e a esclerofilia foliar de C. langsdorffii. Estas variáveis também não variaram entre os habitats. É provável que nesses ambientes a esclerofilia não esteja relacionada apenas com a herbivoria, e que alguns herbívoros podem não ser afetados negativamente ou beneficiados pela concentração de taninos encontrada. O lançamento de folhas por C. langsdorffi apenas no fim da estação seca na Floresta Decidual pode constituir um mecanismo de escape de herbivoria. Porém, para uma análise mais detalhada há necessidade de estudos a longo prazo sobre nos diversos tipos de ambientes nos quais esta espécies esta distribuída bem como da pressão exercida pelos seus herbívoros.
Palabras clave: Defesa química; Estresse higrotérmico; Massa foliar especifica.
ABSTRACT. Sclerophylly, tannins and herbivorous insects associated with Copaifera lagsdorffii Desf. (Fabaceae:Caesalpinioideae) in a Cerrado-Caatinga transition, Brazil: The richness and abundance of free-feeding insect herbivores may vary according to leaf sclerophylly, tannin concentration, and habitat conditions. We sampled 12 individuals of Copaifera langsdorffii in three different habitat physiognomies found in a cerrado-caatinga transition area in northern Minas Gerais, Brazil: 1) Cerrado ("sensu stricto"), 2) Riparian Forest, and 3) Seasonally Dry Forest (n=36 individuals). Higher richness and abundance of chewing herbivores were observed on C. langsdorffii in the Riparian Forest and Cerrado compared to Seasonally Dry Forest. On the other hand, the richness and abundance of sap-sucking herbivores did not differ among the habitats. No statistically significant differences were observed in leaf sclerophylly and tannin concentration among physiognomies, as well as no relationship was found among tannin concentration, leaf sclerophylly and richness and abundance of any of the insect guilds studied. The flushing of leaves by C. langsdorffi at the end of the dry season in the deciduous forest may be a mechanism to escape herbivores. Otherwise, for a detailed analysis long term studies at the vegetation formation where the host plant species is distributed and the selective pressures exerted by its herbivores are called for.
Keywords: Chemical defenses; Hygrothermal stress; Specific leaf mass.
INTRODUÇÃO
Os resultados das interações planta-herbívoro são afetados por variações nas condições bióticas e/ou abióticas locais (White 1969; Danks 2002). Os insetos herbívoros compreendem vários grupos com diferentes formas de alimentação que respondem distintamente a qualidade do hospedeiro e do ambiente (Fernandes & Price 1988; Weis & Berembaum 1989; Ribeiro et al. 1998). Variações na disponibilidade de água (White 1969; Fernandes & Price 1988, 1991) e nutrientes do solo (Coley et al. 1985) afetam a qualidade da planta que pode estender seus efeitos para os níveis tróficos superiores (Fernandes et al. 1999).
Descrever as interações entre e dentro dos níveis tróficos é imprescindível para compreender os padrões de distribuição das espécies e estabelecer estratégias para a conservação dos recursos naturais (Basset et al. 2003). No Brasil, estudos relacionados a padrões de diversidade têm enfocado questões amplas, como a biodiversidade do Cerrado, Mata Atlântica e Floresta Amazônica (Myers et al. 2000). Estudos que enfocam padrões de distribuição de insetos herbívoros associados a um hospedeiro particular entre habitats ainda são escassos (mas veja Fernandes & Price 1992; Ribeiro et al. 1998; Cuevas-Reyes et al. 2004). Desta forma, questões relativas a variações na defesa de plantas entre habitats e seus efeitos na fauna de herbívoros associada ainda permanecem sem respostas.
A esclerofilia é caracterizada como um conjunto de defesas físicas com efeitos negativos na herbivoria (Turner 1994). Este mecanismo de defesa é comum em plantas crescidas em solos pobres em nutrientes, baixa disponibilidade de água e alta irradiação solar (Gonçalves-Alvim et al. 2006). Habitats xéricos tendem a apresentar vegetação esclerófila com elevada longevidade foliar e aumento da razão C/N (Janzen 1974; Bryant et al. 1983; Fernandes & Price 1991, 1992). Nestas condições as plantas produzem maior quantidade de compostos secundários de alto peso molecular e de caráter cumulativo, como os taninos (Coley et al. 1985). Os taninos são compostos de defesa química constitutiva cujo efeito é dependente da dosagem; sendo caracterizados principalmente por sua função anti-herbivoria, precipitando proteínas no trato intestinal dos herbívoros (Coley & Barone 1996; Kutchan 2001). Altas concentrações de taninos e características esclerófilas podem constituir barreiras para a alimentação de herbívoros generalistas especialmente em ambientes xéricos (Fernandes & Price 1988).
Este estudo teve como objetivo avaliar o efeito da esclerofilia foliar e da concentração de taninos na riqueza e abundância de herbívoros de vida livre associados a Copaifera langsdorffii Desf. (Fabaceae: Caesalpinioideae) entre habitats de Cerrado, Mata de Galeria e Floresta Estacional Decidual em uma área de transição na região sudeste do Brasil. Três hipóteses foram testadas: (i) indivíduos de C. langsdorfii presentes em habitats xéricos apresentam maiores esclerofilia foliar e concentração de taninos; (ii) a abundância e riqueza de insetos herbívoros de vida livre associados a C. langsdorffii são maiores nas plantas presentes em habitats mésicos; e (iii) a abundância e riqueza de herbívoros apresentam relação inversa com a esclerofilia foliar e a concentração de taninos.
MATERIAL E MÉTODOS
Área de Estudo
O trabalho foi desenvolvido na Área de Proteção Ambiental do Rio Pandeiros (15º36'02.7" S, 44º40'07.5" W), município de Januária, norte do estado de Minas Gerais, Brasil. Esta unidade de conservação está inserida na transição entre os biomas da Caatinga e do Cerrado, cobrindo uma área de cerca de 210000 ha. As formações vegetais dominantes na APA do Rio Pandeiros são o Cerrado "sensu stricto", as Matas de Galerias e as Florestas Estacionais Deciduais. O clima é do tipo Aw (Segundo Köppen) com temperatura média anual de 23.6 ºC e precipitação média anual de 1057.4 mm (IGA 2006; Santos et al. 2007).
O Cerrado é um tipo de savana, apresentando árvores tortuosas (3-6 m) e vegetação xeromórfica enquanto a Caatinga é um tipo de vegetação com folhas reduzidas e muitos espinhos, dentre a qual as florestas decíduas apresentam como principal adaptação ao es tresse hídrico à queda de folha sincronizada durante a estação seca (Ribeiro & Walter 1998). O Cerrado apresenta grande déficit hídrico e suas plantas apresentam menor taxa de crescimento e folhas caracteristicamente esclerófilas (Ribeiro et al. 1998; Gonçalves-Alvim 2006). As Matas de Galeria correspondem a habitats higrotermicamente menos estressados, com plantas de crescimento mais rápido e folhas menos esclerofilas, que favorecem a colonização por insetos herbívoros de vida livre (Fernandes & Price 1991, 1992). As Florestas Estacionais Deciduais são formações vegetais muito pouco estudadas, as árvores deste habitat tendem a apresentar altas taxas de crescimento e folhas pouco esclerófilas (Ribeiro & Walter 1998; Sanchez-Azofeifa et al. 2005).
Os efeitos do habitat na esclerofilia, na produção de taninos e na fauna de insetos herbívoros de vida livre associados a Copaifera langsdorffii foram avaliados comparando-se estas variáveis em 36 árvores distribuídas equitativamente entre os habitats de Cerrado (n=12), Floresta Estacional Decidual (n=12) e Mata Ciliar (n=12). Os habitats amostrados na área de estudo eram adjacentes entre si. À medida que se distancia do rio Pandeiros é observada a formação de florestas decíduas nos solos onde há afloramentos de calcário (500-750 m), sendo estas florestas secas circundadas por solos arenosos que apresentam vegetação de Cerrado. Em alguns casos, podem ser observadas formações em que a floresta seca abrange até a margem do rio. Para cada habitat foram escolhidos três sítios amostrais, sendo escolhido quatro indivíduos de C. langsdorffii por sítio. As árvores selecionadas em cada habitat apresentavam altura entre 5 e 8 m, Circunferência a Altura do Peito (CAP) acima de 20 cm, boas condições fitossanitárias (ausência aparente de doenças e infestações parasitárias) e distância mínima entre os indivíduos de 15 m. As coletas foram realizadas durante três dias consecutivos, sendo amostrado um habitat por dia. O número total de plantas amostradas praticamente equivale a população local de C. langsdorffii em cada sítio estudado. O trabalho foi realizado no mês de outubro de 2007, quando C. langsdorffi apresentava folhas novas e completamente expandidas, sendo este período evidenciado como o de preferência para o ataque de herbívoros (Ribeiro & Pimenta 1991; veja também Feeny 1976; Coley & Barone 1996). O período amostral corresponde ao fim da estação seca, na qual C. langsdorffii era a única espécie vegetal com folhas verdes presente na Floresta Estacional Decidual.
A Planta Hospedeira
Copaifera langsdorffii Desf. (Fabaceae: Caesalpinioideae), conhecida popularmente como pau d'óleo ou copaíba, é uma espécie arbórea de até 35 m de altura (Ribeiro et al. 2001). A árvore apresenta deciduidade marcante no período de julho a agosto. O brotamento das folhas ocorre nos meses de setembro a outubro, podendo ser facilmente observado pelo aspecto avermelhado da copa (Pedroni et al. 2002). No Brasil, a espécie ocorre nos estados de Minas Gerais, Goiás, Mato Grosso, São Paulo e Paraná (Lorenzi 1992), em fisionomias de cerrado "sensu stricto", matas de galeria, matas secas e cerradões (Almeida et al. 1998), onde é atacada por vários herbívoros (Pedroni 1993; Almeida et al. 2006).
Esclerofilia Foliar
Trinta folhas maduras (completamente expandidas) de cada planta foram coletadas aleatoriamente ao redor da copa das árvores e prensadas. Estas folhas foram levadas para o laboratório e digitalizadas com o auxílio do software ImageJ para determinação da área foliar (Rasband 2006). Em seguida, as folhas foram secas por 72 h a 60 ºC, e então pesadas em balança analítica. A esclerofilia foliar foi estimada através do cálculo da Massa Foliar Especifica (MFE=peso seco/área foliar) (Turner 1994; Madeira et al. 1998; Gonçalves-Alvim et al. 2006).
Quantificação de Taninos
Os taninos foram quantificados através do método de difusão radial (Hagerman 1987). Assim, trinta folhas maduras (completamente expandidas) foram coletadas de cada planta e dessecadas em estufa por 72 h a 45 ºC. Neste método, o sobrenadante das amostras (50 mg com 1 mL de metanol), após centrifugação (11000 RPM/15 min) foi aplicado diretamente para a difusão em gel: 20 µL do extrato, adicionados à placa Petri contendo uma mistura de ágar e proteína (albumina bovina). O extrato foi colocado em círculos uniformes de 2.8 mm de diâmetro e eqüidistantes 1.5 cm perfurados na agarose. As placas foram seladas com parafilme e incubadas a 30 ºC por 96 h. O extrato de tanino após reação com a albumina forma um precipitado opaco no qual o diâmetro ao quadrado é proporcional à concentração de taninos no extrato (Hagerman 1987). Foi utilizado ácido tânico para a construção da curva padrão.
Amostragem dos Herbívoros de Vida Livre
Os insetos de vida livre foram coletados pelo método de batimento entomológico proposto por Basset (1999) e modificado por Ribeiro et al. (2005). Assim, foram realizadas dez batidas em três ramos por planta. Os ramos selecionados apresentavam tamanhos similares (cerca de 30 cm de comprimento e folhas em estágios similares de maturação) e estavam posicionados em diferentes partes da copa, de forma a homogeneizar os efeitos das diferentes condições micro-climática ao longo do dossel. As coletas em todos os ambientes foram realizadas no período da manhã (8:00 às 12:00 h). Os insetos coletados foram levados para o laboratório, onde foram agrupados em duas guildas (sugadores e mastigadores), separados em morfoespécies e identificados até o menor nível taxonômico possível. Foram considerados como sugadores, insetos adultos e juvenis das sub-ordens Auchenorryncha, Sternorryncha e demais Heteroptera herbívoros. Os herbívoros mastigadores foram compostos por insetos adultos das ordens Coleoptera, Orthoptera e Phasmida, incluindo larvas da ordem Lepidoptera (Borror et al. 2002).
Análise dos dados
Para verificar se a riqueza e abundância de herbívoros mastigadores e sugadores variam entre os habitats, em função da esclerofilia e da concentração de taninos foram construídos modelos lineares generalizados (GLM). Os modelos foram submetidos a análise de resíduos para verificar a adequação do modelo à distribuição de erro assumida (Crawley 2002). Foi utilizada distribuição de erro Poisson, corrigida para Quasi-Poisson quando os dados eram sobredispersos (Crawley 2002). Para avaliar a esclerofilia e a concentração de taninos entre os habitats também foram realizados GLM, ajustados a uma distribuição de erro normal. Após o término das análises, os termos qualitativos não significativos foram agrupados através de análises de contraste. As análises foram realizadas através do software R v2.6.2 (R Development Core Team 2008).
RESULTADOS
A esclerofilia foliar de C. langsdorffii não variou estatisticamente entre os habitats de Cerrado (11.3±0.57 mg/cm2), Floresta Decidual (10.0±0.95 mg/cm2) e Mata Ciliar (9.0±0.37 mg/cm2) (F2,33=2.704; P=0.081). Similarmente, a concentração de taninos também não diferiu estatisticamente entre os habitats de Mata Ciliar (3.3±0.09 mg/g), Cerrado (3.2±0.11mg/g) e Floresta Decidual (3.1±0.07 mg/g) (F2,33=3.753; P=0.073).
Foi amostrado um total de 375 insetos herbívoros de vida livre distribuídos em 34 morfoespécies. A abundância (239 indivíduos) e a riqueza de insetos mastigadores (19 morfoespécies) foi maior que a de sugadores (136 indivíduos e 15 morfoespécies). Dentre os insetos mastigadores, a ordem Coleoptera foi a mais representativa (oito famílias), sendo que a família Curculionidae foi a mais rica e abundante (seis morfoespécies e 199 indivíduos). A guilda de insetos sugadores foi composta com maior riqueza pelas famílias Psyllidae e Tingidae, ambas com 3 morfoespécies, enquanto que a família Psyllidae foi mais abundante, com 69 indivíduos amostrados (Tabela 1).
Tabela 1. Abundância e riqueza das guildas de insetos de vida livre associados a Copaifera langsdorffii nos habitats de Cerrado, Mata Ciliar e Floresta Decidual localizados em área de transição Cerrado-Caatinga no médio São Francisco, norte de Minas Gerais, Brazil (n=36).
Table 1. Abundance and richness of free-feeding insect herbivores associated with Copaifera langsdorffii in three different habitats: Cerrado, Gallery Forest and Deciduous Forest in Cerrado-Caatinga transition, Minas Gerais, Brazil (n=36).

As diferentes guildas responderam de forma distinta aos habitats. A abundância e a riqueza de mastigadores foram maiores no Cerrado e Mata Ciliar, quando comparados a Floresta Decidual (Tabela 2; Figura 1 a-b). Entretanto, a riqueza e a abundância de insetos sugadores não variaram entre os habitats estudados (Tabela 2; Figura 2 a-b). Além disto, observouse que a riqueza e a abundância de ambas as guildas de insetos herbívoros não variaram em função da esclerofilia foliar, concentração de taninos ou interação destas variáveis entre os o habitat (Tabela 2).
Tabela 2. Modelos Lineares Generalizados completos para avaliar o efeito da concentração de taninos, massa foliar específica (MFE) e habitat da planta hospedeira sobre a abundância e riqueza de insetos herbívoros mastigadores e sugadores associados a Copaifera langsdorffii (n=36) em zona de transição Cerrado-Caatinga no norte de Minas Gerais, Brasil. A distribuição de erro é de Quasi-Poisson, exceto para a riqueza de sugadores que é Poisson.
Table 2. Deviance analyses of the complete models to evaluate the effects of habitat, specific leaf mass (SLM), and tannin concentration on response variables (abundance and richness of chewing and sap-sucking insect herbivores) associated with Copaifera langsdorffii (n=36) in Cerrado-Caatinga transition in Minas Gerais, Brazil. The error distribution is Quasi-Poisson except for Sap-sucking richness which is Poisson.
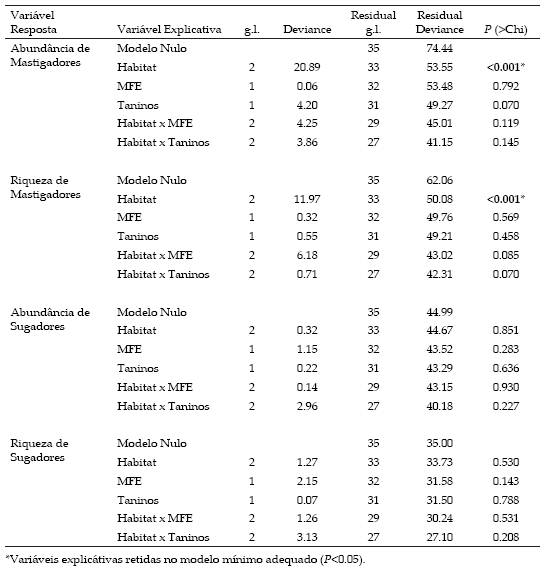

Figura 1. (a) Abundância e (b) riqueza média de insetos mastigadores associados a indivíduos de Copaifera langsdorffii em habitats de Cerrado, Mata Ciliar e Floresta Decidual localizados em área de transição Cerrado-Caatinga no norte de Minas Gerais, Brasil (n=36; média±EP).
Figure 1. (a) Abundance and (b) richness of chewing insects associated with Copaifera langsdorffii in three different habitats: Cerrado, Gallery Forest and Deciduous Forest in Cerrado-Caatinga transition in Minas Gerais, Brazil (n=36; mean±SE).

Figura 2. Abundância (a) e riqueza (b) média de insetos sugadores associados a indivíduos de Copaifera langsdorffii em habitats de Cerrado, Mata Ciliar e Floresta Decidual localizados em área de transição Cerrado-Caatinga no norte de Minas Gerais (n=36; média±EP).
Figure 2. (a) Abundance and (b) richness sap-sucking insects associated with Copaifera langsdorffii in three different habitats: Cerrado, Gallery Forest and Deciduous Forest in Cerrado-Caatinga transition in Minas Gerais, Brazil (n=36; mean±SE).
DISCUSSÃO
A composição das famílias de herbívoros encontradas neste estudo corrobora os resultados descritos por outros trabalhos realizados em florestas tropicais, na qual as famílias Curculionidae, Tingidae e Psyllidae estão entre as mais representativas (Basset 1999; Campos et al. 2006). Outros estudos também registraram um pequeno número de espécies de grande abundância e grande riqueza de espécies relativamente raras (Ribeiro et al. 1998; Varanda & Pais 2006), fato denominado comum para região tropical (Price et al.1995).
A abundância e riqueza de insetos mastigadores foram afetadas pelo habitat ocupado pela planta hospedeira. A abundância e riqueza de mastigadores foram menores na Floresta Decidual do que no Cerrado e Mata Ciliar, respectivamente. Em ambientes sazonais a estação seca é marcada pela deciduidade, o que pode conferir alto xeromorfismo a esses ambientes e baixa adaptabilidade para herbívoros de vida livre (Fernandes & Price 1992). Na estação seca o ambiente se torna ainda mais adverso devido a altas temperaturas, baixa umidade do ar e ausência quase total de folhas, podendo os mastigadores generalistas migrar para a Mata Ciliar e/ou Cerrado em busca de recursos (Stanton 1975; Neves et al. No prelo). Desta forma, nos habitats sempre-verdes, o dossel fechado pode gerar um microclima mais favorável a estes organismos, sendo menos afetados pela incidência de vento, temperatura e luz (Fernandes & Price 1988). De modo geral, quanto mais heterogêneo é um habitat, maior será a variedade de recursos oferecidos (Bell et al. 1991), o que também poderá favorecer a riqueza e abundância de mastigadores generalistas nos habitats de Cerrado e Mata Ciliar, uma vez que C. langsdorffi era a única fonte de recursos para folívoros na Floresta Decidual.
A abundância e riqueza de sugadores não variaram em função do habitat. Para insetos que se alimentam de seiva, a qualidade nutricional da planta é um fator importante na seleção do hospedeiro (Cook & Denno 1994). Insetos herbívoros sugadores, podem, portanto, apresentar maior especificidade ao hospedeiro do que mastigadores (Ribeiro & Pimenta 1991; Styrsky & Eubanks 2007). Alguns estudos demonstram sincronismo entre herbívoros sugadores e a fenologia da planta hospedeira (Ribeiro & Pimenta 1991), sugerindo menor efeito do habitat para esta guilda. Weis & Berembaum (1989) encontraram menor variação da riqueza de herbívoros sugadores do que mastigadores em relação à distribuição da planta hospedeira, enquanto Ribeiro et al. (1998) não observaram variação na abundância de herbívoros sugadores entre ambientes xérico e mésico dentro da mesma estação. Desta forma, sugadores podem ser mais especialistas e sofrerem menor efeito das variações ambientais entre habitats, provavelmente devido a pré-adaptações, não avaliadas neste estudo.
Não foi encontrada nenhuma relação significativa entre riqueza e abundância de insetos herbívoros em função da esclerofilia foliar e da concentração de taninos. Da mesma forma, a esclerofilia foliar e a concentração de taninos não variaram significativamente entre os habitats de Cerrado, Mata Ciliar e Floresta Decidual. A esclerofilia não está relacionada apenas com a herbivoria, mas também na adaptação para conservação de água, nutrientes e proteção contra alta radiação solar (Fernandes & Price 1988, 1991; Turner 1994; Gonçalves-Alvim et al. 2006). Madeira et al. (1998) não encontraram variação na concentração de taninos na espécie Chamaecrista linearifolia (Fabaceae) que ocorria em diferentes altitudes. Em resultados semelhantes, Ribeiro et al. (1999) também não observaram variação na concentração de polifenóis em várias espécies de plantas esclerofilas em diferentes habitas mésico e xérico ao longo de um gradiente altitudinal tropical. Estes mesmos autores também não observaram variação na abundância e riqueza de insetos sugadores e mastigadores em função da concentração de taninos. Uma provável explicação é a pobreza nutricional regional, incluindo a Mata Ciliar, provavelmente devido ao lixiviamento provocado pelo rio Pandeiros em períodos de fortes tempestades e pela elevação do nível d'água (Janzen 1974). Já a ausência de relação entre herbívoros e a concentração de taninos, pode ser devido a esclerofilia impor uma barreira física principalmente para os mastigadores, o que pode mascarar o efeito direto dos taninos (Ribeiro et al. 1999); ou pelo simples fato de alguns insetos não sofrerem influencia da concentração de taninos encontrada, uma vez que o efeito é dependente da dosagem (Kucthan 2001).
Desta forma, o estresse higrotérmico pode ser um dos fatores que determinam a diversidade de herbívoros mastigadores entre os habitats nesta área de transição Cerrado-Caatinga. O lançamento de folhas por C. langsdorffii apenas no fim da estação seca pode favorecer o escape da herbivoria para os indivíduos presentes na Floresta Decidual (Feeny 1976; Coley & Barone 1996; Sloan et al. 2006). Isto é reforçado, uma vez que a menor diversidade de mastigadores foi encontrada durante este período para Floresta Decidual quando comparada ao Cerrado e Mata Ciliar. Por outro lado, herbívoros sugadores mostraram menor variação quanto ao habitat estudado. A esclerofilia mostrouse uma característica comum a planta hospedeira independente do habitat amostrado. Este fato pode ser reflexo da proximidade entre os habitats e de características como solos pobres e uma alta radiação solar, o que confere alta similaridade química, apesar da existência de um gradiente de umidade. Porém, estudos mais detalhados e a longo prazo são necessários para um melhor entendimento das relações herbívoros planta e dinâmica das defesas químicas de plantas em áreas de transição.
AGRADECIMENTOS
Agradecemos a Gisele C.O. Menino e a Magnel L. Oliveira pela ajuda durante o trabalho de campo, e em especial a Frederico S. Neves pelas críticas iniciais nas primeiras versões deste manuscrito e pela assistência em laboratório. Agradecemos também o Instituto Estadual de Florestas (IEF), a Companhia Energética do Estado de Minas Gerais (CEMIG) e ao CNPq (47.2811/2006-1, 30.9633/ 2007-9, 55.5980/ 2006-5) e aos Programa de Pós-graduação em Ciências Biológicas (Universidade Estadual de Montes Claros, MG) e de Pós-Graduação em Ecologia, Conservação e Manejo da Vida Silvestre (Universidade Federal de Minas Gerais).
BIBLIOGRAFIA
ALMEIDA, SP; CEB PROENÇA; SM SANO & JF RIBEIRO. 1998. Cerrado: espécies vegetais úteis. Planaltina: Embrapa, 464 pp. [ Links ]
ALMEIDA, CIM; GLD LEITE; SL ROCHA; MML MACHADO & WCH MALDONADO. 2006. Fenologia e artrópodes de Copaifera langsdorffii Desf. no cerrado. Revista Brasileira de Plantas Medicinais 8:64-70. [ Links ]
BASSET, Y. 1999. Diversity and abundance of insect herbivores collected on Castanopsis acuminatissima (Fabaceae) in New Guinea: relationships with leaf production and surrounding vegetation. European Journal of Entomology 96:381-391. [ Links ]
BASSET, Y; V NOVOTNY; SE MILLER & RL KITCHING (EDS). 2003. Arthopods of tropical forests spatio-temporal dynamics and resource use in the canopy. Cambridge University Press, 474 pp. [ Links ]
BELL, SS; ER MACCONY & HR MSUHINSKY. 1991. Habitat structure: the physical arrangement of objects in space. The Journal of Ecology 79:558-558. [ Links ]
BORROR, DJ; CA TRIPLEHORN & NF JOHNSON (EDS). 2002. An introduction to the study of insects. Saunders College Publing, 875 pp. [ Links ]
BRYANT, JP; FS CHAPIN III & DR KLEIN. 1983. Carbon/Nutrient Balance of Boreal Plants in Relation to Vertebrate Herbivory. Oikos 40:357-368. [ Links ]
CAMPOS, RL; HL VASCONCENLOS; SP RIBEIRO; FS NEVES & JP SOARES. 2006. Relationship between tree size and insect assemblages associated with Anadenanthera macrocarpa. Ecography 29:442-450. [ Links ]
COLEY, PD; JP BRYANT & FS CHAPIN III. 1985. Resource availability and plant anti-herbivore defense. Science 230:895-899. [ Links ]
COLEY, PD & JA BARONE. 1996. Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systematics 27:305-35. [ Links ]
COOK, AG & RF DENNO. 1994. Planthopper/plant interactions: feeding behavior, plant nutrition, plant defense, and host plant specialization. Pp. 114-139 en: Denno, RF & TJ Perfect (eds.). Planthoppers: Their Ecology and Management. Chapman & Hall, 799 pp. [ Links ]
CRAWLEY, M. 2002. Statistical computing: An introduction to data analysis using S-Plus. John Wiley & Sons, 761 pp. [ Links ]
CUEVAS-REYES, P; M QUESADA; P HANSON; R DIRZO & K OYAMA. 2004. Diversity of gall-inducing insects in a Mexican tropical dry forest: the importance of plant species richness, life-forms, host plant age and plant density. Journal of Ecology 92:707-716. [ Links ]
DANKS, HV. 2002. Modification of adverse conditions by insects. Oikos 99:10-24. [ Links ]
FEENY, P. 1976. Plant apparency and chemical defense. Recent Advances in Phytochemistry 10: 1-40. [ Links ]
FERNANDES, GW & PW PRICE. 1988. Biogeographical gradients in galling species richness: tests of hypotheses. Oecologia 76:161-167. [ Links ]
FERNANDES, GW & PW PRICE. 1991. Comparison of tropical and temperate galling species richness: the role of environmental harshness and plant nutrient status. Pp. 91-115 en: PW Price; T Lewinsohn; GW Fernandes & WW Benson (eds.). Plant–animal interactions: evolutionary ecology in tropical and temperate regions. John Wiley, 640 pp. [ Links ]
FERNANDES, GW & PW PRICE. 1992. The adaptative significance of insect gall distribution: survivorship of species in xeric and mesic habitat. Oecologia 90: 14-20. [ Links ]
FERNANDES, GW; M FAGUNDES; RL WOODMAN & PW PRICE. 1999. Ant effects on three-trophic level interactions: Plant, galls, and parasitoids. Ecological Entomology 24:411-415. [ Links ]
GONÇALVES-ALVIM, SJ; G KORNDORF & GW FERNANDES. 2006. Sclerophylly in Qualea parviflora (Vochysiaceae): influence of herbivory, mineral nutrients, and waters status. Plant Ecology 187: 153-162. [ Links ]
HAGERMAN, AE. 1987. Radial diffusion method for determining tannin in plant extracts. Journal of Chemical Ecology 13:437-449. [ Links ]
IGA - INSTITUTO DE GEOCIÊNCIAS APLICADAS - GOVERNO DO ESTADO DE MINAS GERAIS. 2006. Áreas de Proteção Ambiental no Estado de Minas Gerais. Demarcação e Estudos para o Pré-Zoneamento Ecológico: Apa Bacia do Rio Pandeiros. Relatório Técnico, Instituto Estadual de Florestas (IEF), 271 pp. [ Links ]
JANZEN, DH. 1974. Tropical blackwater rivers, animals and mast fruiting by the Dipterocarpaceae. Biotropica 6:69-103. [ Links ]
KUTCHAN, TM. 2001. Ecological arsenal and developmental dispatcher. The paradigm of secondary metabolism. Plant Physiology 125: 58-60. [ Links ]
LORENZI, H. 1992. Árvores brasileiras: Manual de identificação e cultivo de plantas arbóres nativas do Brasil. Nova Odessa: Plantarum, 352 pp. [ Links ]
MADEIRA, JA; KT RIBEIRO & GW FERNANDES. 1998. Herbivory, tannins and sclerophylly in Chamaecrista linearifolia (Fabaceae) along an altitudinal gradient. Brazilian Journal of Ecology 2:24-29. [ Links ]
MYERS, N; RA MITTERMEIER; CG MITTERMEIER; GAB FONSECA & J KENT. 2000. Biodiversity hotspots for conservation priorities. Nature 403:853-858. [ Links ]
NEVES, FS, LS ARAÚJO; M FAGUNDES; MM ESPÍRITO-SANTO; GW FERNANDES ET AL. 2009. Canopy herbivory and insect herbivore diversity in a dry forest-savana transition in Brazil. Biotropica (No Prelo). [ Links ]
PEDRONI, F. 1993. Ecologia da Copaíba (Copaifera langsdorffii Desf. Caelsapiniaceae) na Reserva Municipal de Santa Genebra, Campinas, SP. Dissertação (Mestrado em Ciências Biológicas-Biologia Vegetal). Universidade Estadual de Campinas, 122 pp. [ Links ]
PEDRONI, F; M SANCHEZ & AM SANTOS. 2002. Fenologia da copaíba (Copaifera Langsdorffii Desf. – Leguminosae, Caesalpinioideae) em uma floresta semidecídua no sudeste do Brasil. Revista Brasileira de Botânica 25:183-194. [ Links ]
PRICE, PW; IR DINIZ; HC MORAIS & ESA MARQUES. 1995. The abundance of insect herbivore species in the tropics: the high local richness of rare species. Biotropica 27(4):468-478. [ Links ]
R. DEVELOPMENT CORE TEAM. 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing, ISBN 3-900051-07-0, URL http://www.R-project.org. [ Links ]
RASBAND, WS. 2006. ImageJ, U. S. National Institutes of Health, Bethesda. http://rsb.info.nih.gov/ij. [ Links ]
RIBEIRO, JF & BMT WALTER. 1998. Fitofisionomias do bioma Cerrado. Pp. 89-166 en: Sano, SM & SP Almeida (eds). Cerrado: Ambiente e Flora. Planaltina: Embrapa Cerrados, 556 pp. [ Links ]
RIBEIRO, JF; CEL FONSECA & JC SOUSA-SILVA. 2001. Cerrado: Caracterização e Recuperação de Matas de Galeria. Planaltina: Embrapa Cerrados, 899 pp. [ Links ]
RIBEIRO, SP & HR PIMENTA. 1991. Padrões de abundância e de distribuição temporal de herbívoros de vida livre em Tabebuia ochracea (Bignoniaceae). Anais da Sociedade Entomológica do Brasil 20:428-448. [ Links ]
RIBEIRO. SP; MAA CARNEIRO & GW FERNANDES. 1998. Free-feeding insect herbivores along environmental gradients in Serra do Cipó: basis for a management plan. Journal of Insect Conservation 2:107-118. [ Links ]
RIBEIRO, SP; OA BRAGA, CHL SILVA & GW FERNANDES. 1999. Leaf polyphenols in brazilian melastomataceae: sclerophylly, habitats, and insect herbivores. Ecotropica 5:137-146. [ Links ]
RIBEIRO, SP; PP BORGES; C GASPAR; C MELO; ARM SERRANO; ET AL. 2005. Canopy insect herbivores in the Azorean laurisilva forests: key host plant species in a highly generalist insect community. Ecography 28:315-330. [ Links ]
SANCHEZ-AZOFEIFA, GA; M QUESADA; JP RODRÍGUEZ; JM NASSAR; KE STONER; ET AL. 2005. Research priorities for Neotropical dry forests. Biotropica 37:477-485. [ Links ]
SANTOS, RM; FA VIEIRA; M FAGUNDES & YRF NUNES. 2007. Riqueza e similaridade florística de oito remanescentes florestais no norte de Minas Gerais, Brasil. Revista Árvore 31:135-144. [ Links ]
SLOAN, SA; JK ZIMMERMAN & AB SABAT. 2006. Phenology of Plumeria alba and its herbivores in a tropical dry forest. Biotropica 39:195-201. [ Links ]
STANTON, N. 1975. Herbivore pressure on two types of tropical forests. Biotropica 7:8-11. [ Links ]
STYRSKY, JD & MD EUBANKS. 2007. Ecological consequences of interactions between ants and honeydew-producing insects. Proceedings of the Royal Society of London. Series B, Biological Sciences 274:151-164. [ Links ]
TURNER, IM. 1994. Sclerophylly: primarily protective? Functional Ecology 8:669-675. [ Links ]
VARANDA, EM & MP PAIS. 2006. Insect folivory in Didymopanax vinosum (Apiaceae) in a vegetation mosaic of Brazilian Cerrado. Brazilian Journal of Biology 66:671-680. [ Links ]
WEIS, AE & MR BERENBAUM. 1989. Herbivorous insects and green plants. Pp. 123-162 en: Abrahamson, WG (ed.). Plant-Animal Interaction, Graw-Hill, 520 pp. [ Links ]
WHITE, TCR. 1969. An index to measure weather induced stress of trees associated with out-breaks of psyllids in Australia. Ecology 50:905-909. [ Links ]

