










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.19 n.3 Córdoba sept./dic. 2009
COMUNICACIÓN BREVE
Hábitos alimentarios de larvas de Trichoptera (Insecta) de una cuenca subtropical
María C Reynaga*
* CONICET, Facultad de Ciencias Naturales, Universidad Nacional de Tucumán, Tucumán, Argentina.
CONICET, Facultad de Ciencias Naturales, Universidad Nacional de Tucumán. Miguel Lillo 205, (4000) Tucumán, Argentina. Email: celina_reynaga@yahoo.com
Recibido: 13 de mayo de 2009;
Fin de arbitraje: 7 de septiembre de 2009;
Revisión recibida: 1 de octubre de 2009;
Aceptado: 14 de octubre de 2009
RESUMEN. Los tricópteros constituyen un elemento importante en el flujo de energía y en la dinámica de nutrientes de los ambientes acuáticos debido a que sus larvas despliegan una diversidad amplia de adaptaciones tróficas y explotan una gran variedad de microhábitats. Sin embargo, se dispone de escasa información sobre sus hábitos alimentarios en la región Neotropical. En este trabajo se analizan y definen los grupos funcionales de larvas de tricópteros de una cuenca subtropical andina. Las larvas fueron colectadas en época de aguas bajas e identificadas hasta el nivel de género. Mediante análisis de contenido estomacal se realizaron los perfiles dietarios por género. La mayoría de los géneros prefirieron más de un ítem alimenticio, registrándose en todos los casos materia orgánica particulada fina. Metrichia, Hydroptila, Oxyethira y Marilia se clasificaron como colectores-recolectores; Nectopsyche se clasificó como colector-recolector y triturador secundario. Mortoniella y Helicopsyche presentaron un hábito raspador primario y colector-recolector secundario. Chimarra se clasificó como colector-filtrador. Smicridea y Polycentropus resultaron depredadores de invertebrados y recolectores de partículas retenidas en las redes. Sólo cuatro de los diez géneros analizados se correspondieron con las categorías informadas para otras regiones.
Palabras clave: Grupos funcionales alimenticios; Contenido estomacal; Yungas; Tucumán; Análisis de "cluster".
ABSTRACT. Feeding habits of Trichoptera (Insecta) larvae of the subtropical Basin: The functional feeding groups of aquatic invertebrates indicate both the type of food source used and the feeding mechanism involved. The groups considered were: collectors (gatherers and filterers), shredders, scrapers and predators. Trichoptera larvae display a diversity of trophic adaptations and exploit a variety of microhabitats, but information on their feeding habits in the Neotropical region is still incomplete. The main objective of this paper was to analyze the feeding habits and define the functional feeding groups of Trichoptera recorded in a subtropical Andean basin. The larvae were collected during the low water period and identified to the genus level. Diet profiles were obtained through gut content analysis. Schoener's Proportional Similarity Index was applied to assess the similarity between diet profiles. A complete linkage dendrogram was obtained and significant clusters (dietary patterns) were identified. The majority of taxa examined prefer more than one food item, being fine particle organic matter present throughout all genera. Metrichia, Hydroptila, Oxyethira and Marilia are categorized as collectors-gatherers. Nectopsyche is defined as collector-gatherer and secondarily shredder. Mortoniella and Helicopsyche are primarily scrapers and secondarily collectors-gatherers. Chimarra shows a collector-filterer habit. Smicridea and Polycentropus are basically predators, but they also feed on particles trapped in nets. Only four of the ten genera analyzed here correspond to the functional groups already reported for other regions.
Keywords: Functional feeding groups; Gut contents analysis; Yungas; Tucumán; Cluster analysis.
INTRODUCCIÓN
El estudio de la biodiversidad involucra dimensiones múltiples, que incluyen la variabilidad genética, taxonómica y funcional de los organismos (Heino 2005). Las funciones ecológicas pueden ser descriptas a través de atributos asociados a las especies, que son capaces de reflejar su adaptación a las condiciones ambientales (Townsend & Hildrew 1994). En particular, el conocimiento de los grupos funcionales alimenticios de los ecosistemas lóticos contribuye a una mejor comprensión del funcionamiento de tales sistemas (Cummins 1973). Este método de asignación de especies a grupos funcionales tróficos está basado sobre la asociación entre los mecanismos morfológicos y de comportamiento alimentario y el tipo de alimento ingerido, y permite agrupar a los insectos acuáticos en las categorías de colectores (recolectores y filtradores), trituradores, raspadores y depredadores. Una de sus ventajas es su capacidad para representar la dependencia de la comunidad lótica hacia recursos alimenticios particulares, lo cual permite detectar tanto los patrones de cambio que suceden a lo largo del sistema lótico, desde la zona de cabecera hasta los tramos inferiores (Vannote et al. 1980), como las alteraciones promovidas por el impacto antrópico (Cummins 1993). Esta clasificación funcional fue desarrollada para el hemisferio norte, por cuanto la designación funcional correcta de los grupos taxonómicos representativos en la región Neotropical está en proceso de ajuste (Cummins et al. 2005).
Las larvas de Trichoptera constituyen un elemento importante en el flujo de energía y en la dinámica de nutrientes de los ambientes acuáticos debido a que despliegan una diversidad amplia de adaptaciones tróficas y a que explotan una variedad de microhábitats (Flint et al. 1999). La gran importancia de este grupo en la estructura trófica de los ríos contrasta con la escasa información disponible sobre los hábitos alimentarios de las larvas de Trichoptera en la región Neotropical (Albariño & Valverde 1998; Posada-García & Roldán-Pérez 2003; Motta & Uieda 2004; Díaz Villanueva & Trochine 2005; Cummins et al. 2005; Gil et al. 2006; Tomanova et al. 2006; Reynaga & Rueda Martín, en prensa).
El objetivo de este trabajo consiste en determinar los hábitos alimentarios y definir los correspondientes grupos funcionales de los estadios larvales de tricópteros registrados en la Cuenca del Río Lules (Tucumán, Argentina). La asignación de las especies de macroinvertebrados a un grupo funcional trófico servirá en el futuro para entender un río subtropical andino desde el punto de vista funcional.
MATERIALES Y MÉTODOS
La Cuenca del Río Lules (26°36' S, 65°45' W) es una cuenca subtropical andina que posee una extensión de 820 km2. La vegetación característica de la cuenca es de selva de montaña perteneciente a la región fitogeográfica de Las Yungas (Brown 2000). Esta vegetación se distribuye en una franja altitudinal que se extiende entre los 400 y los 3000 m.s.n.m. El clima es subtropical con régimen monzónico, y presenta un contraste fuerte entre la mitad cálida del año, con tormentas eléctricas, y la mitad fría (de abril a setiembre), en la que sólo se registra 20% de la precipitación anual (Hunzinger 1997). A pesar del porcentaje bajo de lluvia que cae en el invierno, las condiciones de humedad están preservadas gracias a la condensación del aire húmedo, un fenómeno denominado precipitación horizontal. La temperatura también muestra una estacionalidad marcada, con valores promedio de 25 °C en verano y 12 °C en invierno.
Las larvas de Trichoptera fueron recolectadas en siete ríos distribuidos en la Cuenca del Río Lules, durante la época de aguas bajas (setiembre de 2005 y 2006). Las muestras se obtuvieron con red Surber (0.09 m2 de superficie y 300 µ de abertura de malla), y se fijaron con alcohol al 75%. Los tricópteros fueron identificados hasta el nivel de género por medio de las claves disponibles para la región (Angrisano 2001). Se procuró obtener hasta 10 individuos por cada género encontrado en las muestras, a los que se les midió el largo de cuerpo y el ancho de cabeza, para establecer el perfil dietario del estadio correspondiente.
El análisis de alimentación se realizó a través de la disección ventral del tórax para aislar el estomodeo y mesodeo (Peckarsky 1996). El contenido de cada espécimen fue montado con glicerina y observado bajo aumento de 200X con ocular graduado. Se seleccionaron 15 transectas al azar, en las que se registró cada ítem alimenticio para la estimación de su porcentaje de cobertura según Reynaga & Rueda Martín (en prensa). Las categorías alimenticias que se consideraron fueron: partículas de sedimento, materia orgánica particulada fina (MOPF<1 mm), materia orgánica particulada gruesa (MOPG>1 mm), micrófitas y restos de invertebrados.
Se evaluó el solapamiento de nicho trófico entre pares de espectros alimenticios a través del índice de Similitud Proporcional de Schoener (SP) (Schoener 1970), el cual se obtuvo restando a la unidad la mitad de la distancia de Manhattan entre los perfiles alimenticios dados en frecuencias relativas. Así, para el par de especímenes (1, 2) el índice es:
![]()
Los valores de SP están contenidos en el intervalo [0;1]. Cero indica la exclusión mutua en la partición del recurso y 1 indica la superposición completa. Se adoptó la sugerencia de Wallace (1981) al considerar el valor 0.6 de SP como umbral de similitud dietaria significativa en términos biológicos.
En función de los valores apareados de solapamiento se construyó un dendrograma de disimilitud con el método de ligamiento completo (Sokal & Rohlf 1995). Los respectivos agrupamientos de perfiles tróficos fueron identificados al usar el valor 0.6 como línea de corte. Finalmente, se atribuyó el grupo funcional alimenticio para cada género atendiendo, en forma conjunta, tanto al modo de adquisición del alimento como su pertenencia a los grupos (o patrones) de perfiles alimenticios antes definidos. Los gráficos y los análisis se hicieron a través del entorno R para matemáticas y estadística (R Development Core Team 2008). Los "scripts" quedan libremente disponibles y pueden ser solicitados a la autora por correspondencia electrónica.
RESULTADOS Y DISCUSIÓN
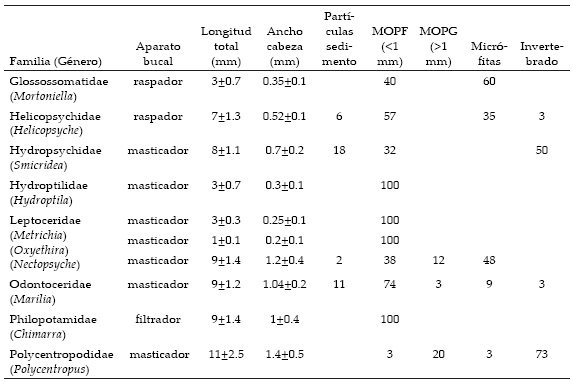
El área de estudio alberga un total de once géneros de tricópteros, distribuidos en nueve familias diferentes. De ese total, se excluyó del análisis al género Atopsyche debido a que su grupo funcional depredador ya ha sido tratado (Reynaga & Rueda Martín, en prensa). En términos generales, la MOPF fue el ítem alimenticio dominante, presente en la dieta de todos los géneros analizados, mientras que la MOPG fue ingerida en proporción menor (Tabla 1).
Tabla 1. Composición dietaria y medidas morfológicas para los géneros de Trichoptera encontrados en el área de estudio. La frecuencia relativa de los items alimenticios está expresada en porcentajes.
Table 1. Average diet composition and morphological measurements of Trichoptera genera found throughout the study area. Relative frequencies for feeding items are in percentage.
El dendrograma permitió extraer siete grupos de patrón alimentario (Figura 1). El Grupo 1 estuvo integrado por individuos que registraron una afinidad de 100% hacia la MOPG, mientras que el Grupo 2 presentó una dieta compuesta por partes semejantes de MOPF e invertebrados (Figura 2). Los individuos con una afinidad mayor a 80% por los invertebrados conformaron el Grupo 3. En el Grupo 4 predominaron las micrófitas (rango 80-100%) y en menor proporción MOPF. El Grupo 5 estuvo integrado en forma primaria por partículas de sedimento y MOPF. El Grupo 6 quedó disociado del resto por la dominancia de MOPF, mientras que el Grupo 7 presentó el perfil más heterogéneo, signado por MOPF (54%), micrófitas (26%), partículas de sedimento (13%) e invertebrados (6%) (Figura 2). La afinidad de cada género hacia los grupos mostró que, por un lado, el Grupo 6 concentró la mayor abundancia y riqueza de géneros, mientras que por otro lado, el género Smicridea presentó los patrones alimenticios más heterogéneos (Tabla 2).
Figura 1. Delimitación de grupos alimenticios a partir del dendrograma basado en la superposición absoluta de perfiles dietarios entre individuos. El índice de Schoener y el ligamiento completo se usaron como métrica y método de aglutinamiento, respectivamente. La línea punteada al valor 0.6 del índice sirve para guiar la selección de agrupamientos significativos.
Figure 1. Feeding groups derived from the dendrogram based on absolute differences between dietary profiles of individuals. The Schoener's Index and complete linkage were used as similarity measure and clustering method, respectively. The dotted line at the 0.6 index value is the guideline to obtain significant groupings.
Figura 2. Diagrama de barras esquemático indicando los perfiles alimenticios promedios para cada grupo (G) de la Figura 1.
Figure 2. Barplot showing the average feeding profiles for the groups (G) delineated out of the Figure 1.
Tabla 2. Ocurrencia de cada uno de los géneros en los agrupamientos (G) establecidos a partir del dendrograma de la Figura 2.
Table 2. Affinity of genera to the clusters (G) recognized along the dendrogram of Figure 2.

Los grupos alimenticios esbozados pueden relacionarse con las categorías preestablecidas de grupos funcionales, lo que permite complementar los datos de contenido estomacal con datos morfológicos y de comportamiento afines, y reforzar o reconsiderar esa asignación original. El perfil del Grupo 1 resultó compatible con individuos de hábito triturador, mientras que los perfiles de los Grupos 2 y 3 fueron compatibles con los hábitos filtradores y depredadores, respectivamente. El espectro dietario del Grupo 4 fue atribuible a organismos raspadores; por su parte, los perfiles dietarios de los grupos 5, 6 y 7 mostraron un predominio de MOPF y la presencia de partículas de sedimento que pueden vincularse con un hábito colector.
Algunos autores consideran que los insectos acuáticos son oportunistas debido a que ingieren una amplia variedad de ítemes alimenticios disponibles en su ambiente (Merrit & Cummins 1996; Covich 1988). En el presente estudio, la mayoría de los géneros prefirieron más de un ítem alimenticio, y en todos los casos analizados se registró la presencia de MOPF. Sin embargo, hubo diferencias en las proporciones de ingesta que se podrían relacionar con una alta disponibilidad de este recurso en el medio según observaran Tomanova et al. (2006).
Los resultados de este trabajo coinciden parcialmente con los documentados por otros autores. El género Chimarra pertenece al grupo colector-filtrador, indicado a partir de su perfil alimentario, dominado por MOPF, y de la morfología de su aparato bucal, apta para filtrar partículas a través de la membrana del labro, lo que coincide con otras evidencias (Wiggins 2004; Tomanova et al. 2006). Con respecto a la familia Hydroptilidae, los resultados obtenidos indican que los géneros Metrichia, Hydroptila y Oxyethira se asociarían con la categoría de colector-recolector. Tomanova et al. (2006) incluyen a los géneros de esta familia dentro del grupo de los raspadores, en primer lugar, y de los colectores-recolectores en menor cantidad. Wiggins & Mackay (1978) y Wiggins (1996) proponen el grupo funcional de los trituradores para el género Hydroptila. El género Mortoniella se asocia con un hábito raspador en primera instancia y colector-recolector de modo secundario, en coincidencia con lo planteado por Tomanova et al. (2006) y Wiggins (2004), lo cual indica que los Glossossomatidae accionan sus mandíbulas raspadoras para obtener el perifiton y las partículas finas que recubren las superficies de las rocas. La clasificación para Helicopsyche también fue la de raspador -en primera instancia- y colector-recolector secundario. Para este género, Wiggins & Mackay (1978) definen el grupo funcional como ramoneador y Cummins et al. (2005) como exclusivamente raspador. Este trabajo clasificó al género Marilia dentro del grupo funcional colector-recolector, mientras que Tomanova et al. (2006) lo clasifican como raspador y colector-recolector. Los Odontoceridae se consideran omnívoros debido a que presentan una dieta versátil que combina MOPF, algas y material animal (Wiggins 2004). Dentro de los grupos capaces de tejer redes, Smicridea y Polycentropus parecerían tener un rol depredador de invertebrados, tal vez procedentes de la deriva, y un hábito recolector de partículas retenidas en las redes. Por esta condición se los considera -en términos generales- como colectores-filtradores, clasificación discutida por Wiggins (2004) refiriendo a Hydropsychidae que capturan en sus redes invertebrados que derivan, otros que depredan sobre larvas de Simulidae, e incluso hace referencia a un grupo que exhibe búsqueda activa del alimento. Este último comportamiento ha sido empíricamente constatado mediante cría en laboratorio de individuos del género Smicridea ante la presencia de larvas de Chironomidae y congéneres de menor tamaño (Rueda Martín, comunicación personal). En lo que respecta al género Polycentropus, es clasificado funcionalmente como depredador (Merrit & Cummins 1996; Wiggins 1996), así como con hábitos de colector-filtrador (Cummins et al. 2005; Gil 2006). Nectopsyche es un género con hábitos nadadores y de acuerdo con los resultados aquí obtenidos se clasificaría como colector-recolector y, potencialmente, como triturador secundario. Cummins et al. (2005) diferencia a los Leptoceridae en sus grupos funcionales entre las formas que nadan (colectores) y las formas fijas (trituradores).
Merece destacarse que sólo cuatro de los diez géneros analizados (Chimarra, Mortoniella, Helicopsyche y Nectopsyche) se correspondieron con las categorías informadas para otras regiones, lo que sugiere que pretender vincular taxones con grupos funcionales precisos es una tarea que difícilmente alcance universalidad. Por el contrario, la calibración de las clasificaciones para el ámbito geográfico de interés parece una tarea ineludible, y que evitará asignaciones funcionales incorrectas. Se sugiere profundizar el estudio de otros insectos acuáticos a fin de complementar a la presente contribución y, de esta manera, aprovechar mejor el potencial analítico que pueden ofrecer las metodologías disponibles y basadas sobre grupos funcionales. De otro modo, la proyección de resultados a otros fines quedaría comprometida o invalidada.
AGRADECIMIENTOS
Por el apoyo brindado agradezco al Dr. H. Fernández, a la Dra. Odile Fossati, al Lic. D. Dos Santos, a la Lic. P. A. Rueda Martín, a los revisores anónimos y a la editora por sus valiosos comentarios. El trabajo fue realizado con el financiamiento del PICT 01-528 y CIUNT 26-G416.
BIBLIOGRAFÍA
ALBARIÑO, RJ & A VALVERDE. 1998. Hábito alimenticio del estado larval de Parasericostoma cristatum (Trichoptera: Sericostomatidae). Rev. Soc. Entomol. Argent. 57:131-135. [ Links ]
ANGRISANO, EB. 2001. Trichoptera. En: Fernández, H & E Domínguez (eds.), Guía para la determinación de los artrópodos bentónicos sudamericanos. Editorial Universitaria de Tucumán, Argentina. [ Links ]
BROWN, AD. 2000. Development threats to biodiversity and opportunities for conservation in the mountain ranges of the upper Bermejo river basin, NW Argentina and SW Bolivia. Ambio 29: 445-449. [ Links ]
COVICH, AP. 1988. Geographical and historical comparisons of neotropical streams: biotic biodiversity and detrital processing in highly variable habitats. J. N. Am. Benthol. Soc. 7: 361-386. [ Links ]
CUMMINS, KM. 1973. Trophic relations of aquatic insects. Annu. Rev. Entomol. 18:183-203. [ Links ]
CUMMINS, KW. 1993. Invertebrates. Pp. 234-250 en: Calow, P & GE Petts (eds.), The Rivers Handbook. Blackwell Scientific, Oxford. [ Links ]
CUMMINS, KW; RW MERRIT & PCN ANDRADE. 2005. The use of invertebrates functional group to characterize ecosystem attributes in selected stream and rivers in south Brazil. Stud. Neotrop. Fauna Environ. 40:69-89. [ Links ]
DÍAZ VILLANUEVA, V & C TROCHINE. 2005. The role of microorganisms in the diet of Verger cf. limnophilus (Trichoptera: Limnephilidae) larvae in a Patagonian Andean temporary pond. Wetlands 25:473-479. [ Links ]
FLINT, OS; R HOLZENTHAL & SC HARRIS. 1999. Nomenclatural and systematic changes in the Neotropical Caddisflies (Insecta: Trichoptera). Insecta Mundi 13:73-84. [ Links ]
GIL, MA; PA GARELIS & EA VALLANIA. 2006. Hábitos alimenticios de larvas de Polycentropus joergenseni Ulmer, 1909 (Trichoptera: Polycentropodidae) en el Río Grande (San Luis, Argentina). Gayana 70: 206-209. [ Links ]
HEINO, J. 2005. Functional biodiversity of macroinvertebrates assemblage along major ecological gradients of boreal headwater streams. Freshwat. Biol. 50:1578-1587. [ Links ]
HUNZINGER, H. 1997. Hydrology of montane forests in the sierra de San Javier, Tucumán, Argentina. Mt. Res. Dev. 17:299-308. [ Links ]
MERRITT, RW & KW CUMMINS. 1996. An Introduction to the Aquatic Insects of North America. Kendall/Hunt. Iowa. [ Links ]
MOTTA, RL & VS UIEDA. 2004. Diet and trophic groups of an aquatic insect Community in a tropical stream. Braz. J. Biol. 64:809-817. [ Links ]
PECKARSKY, BL. 1996. Predator-prey interactions. Pp. 431-451 en: Hauer, FR & GA Lamberti (eds.), Methods in Stream Ecology. Academic Press, California. USA. [ Links ]
POSADA-GARCÍA, JA & G ROLDÁN-PÉREZ. 2003. Clave ilustrada y diversidad de las larvas de Trichoptera en el Nor-Occidente de Colombia. Caldasia 25: 169-192. [ Links ]
R DEVELOPMENT CORE TEAM. 2008. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. [ Links ]
REYNAGA, MC & PA RUEDA MARTÍN. En prensa. Trophic analysis of two species of Atopsyche (Trichoptera: Hydrobiosidae ). Limnologica. [ Links ]
SCHOENER, TW. 1970. Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 51(3):408-418. [ Links ]
SOKAL, RR & FJ ROHLF. 1995. Biometry: the principles and practice of statistics in biological research. 3ra edición. WH Freeman & Co., New York. Pp. 887. [ Links ]
TOMANOVA, S; E GOITIA & J HELEIC. 2006. Trophic levels and functional feeding groups of macroinvertebrates in neotropical streams. Hydrobiologia 556:251-264. [ Links ]
TOWNSEND, CR & AG HILDREW. 1994. Species traits in relation to a habitat templet for river systems. Freshwat. Biol. 31:265-275. [ Links ]
VANNOTE, RL; GW MINSHALL; KM CUMMINS; JR SEDELI & CE CUSHING. 1980. The river continuum concept. Can. J. Fish. Aquat. Sci. 37:130-137. [ Links ]
WALLACE JR, RK. 1981. An assessment of diet-overlap indexes. Trans. Am. Fish. Soc. 110:72-76. [ Links ]
WIGGINS, GB & RJ MACKAY. 1978. Some relationships between systematic and trophic ecology in neartic aquatic insects, with special referent to Trichoptera. Ecology 59:1211-1220. [ Links ]
WIGGINS, GB. 1996. Larvae of the north American Caddisfly Genera (Trichoptera). University of Toronto Press, Toronto. [ Links ]
WIGGINS, GB. 2004. Caddisflies. The Underwater Architects. University of Toronto Press, Toronto. [ Links ]


{kind=link}