










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.20 n.1 Córdoba ene./abr. 2010
ARTÍCULOS ORIGINALES
Forestación con Polylepis australis en suelos erosionados de las Sierras Grandes de Córdoba: evaluación del uso de terrazas y vegetación nodriza
Marcos A Landi * & Daniel Renison
Cátedra de Ecología, Universidad Nacional de Córdoba-CONICET, Córdoba, Argentina.
Cátedra de Ecología, Universidad Nacional de Córdoba-CONICET, C.C. 122, (5000) Córdoba, Argentina. Tel.: +54-351-476-6510.
Email: marcoslandi00@yahoo.com.ar
Recibido: 16 de julio de 2009;
Fin de arbitraje: 14 de febrero de 2010;
Revisión recibida: 20 de marzo de 2010;
Aceptado: 27 de marzo de 2010
RESUMEN. La erosión de los suelos y la consecuente aparición de cárcavas constituyen un problema grave en las montañas del Centro Argentino. La forestación con la especie nativa Polylepis australis es una de las tecnologías disponibles para detener y revertir el deterioro. Sin embargo, la escasa fertilidad y retención del agua por parte de los suelos limitan el éxito de este tipo de manejo. En este trabajo evaluamos el papel de la construcción de terrazas de piedra y la implantación de vegetación potencialmente nodriza como facilitadoras del establecimiento de P. australis. Realizamos un ensayo factorial de implantación de plantines y de semillas de P. australis en cárcavas con distinto grado de protección por terraza y/o vegetación nodriza (césped, Poa stuckertii y sin vegetación) en dos tipos de suelo con menor y mayor profundidad (cárcava sobre horizonte A1 y cárcava sobre horizonte II). Evaluamos la supervivencia y crecimiento de los plantines, el establecimiento de las semillas y el crecimiento de plántulas luego de dos/tres años. La supervivencia total de los plantines fue de 96% y resultó similar para todas las situaciones. El efecto de la vegetación nodriza sobre el crecimiento de los plantines dependió del tipo de suelo. En las cárcavas sobre suelo menos profundo el uso de terrazas con P. stuckertii fue positivo, mientras que en las ubicadas sobre suelos más profundos observamos un efecto positivo solo en las terrazas sin vegetación nodriza. Independientemente de los tratamientos aplicados, una proporción muy baja de las semillas (inferior a 1%) se estableció como plántulas, mientras que su crecimiento fue mayor en las terrazas vegetadas con P. stuckertii. El crecimiento de las plántulas fue seis veces más lento que el de los plantines. Concluimos que la mejor forma de facilitar la implantación de P. australis en cárcavas de las Sierras Grandes sería mediante el uso de plantines y no de semillas. Recomendamos el empleo de terrazas con vegetación nodriza de P. stuckertii en los suelos menos profundos y de terrazas sin vegetación nodriza en los más profundos.
Palabras clave: Reforestación; Facilitación; Establecimiento de plántulas; Plantines.
ABSTRACT. Forestation with Polylepis australis BITT. in eroded soils of the Sierras Grandes of Córdoba: The mountains of Central Argentina have serious soil erosion problems including the formation of erosion gullies. The forestation with native Polylepis australis trees could be used to stop soil erosion and restore degraded areas. However their establishment in erosion gullies is hampered due to low soil fertility and water content. We evaluated whether P. australis establishment could be facilitated by the construction of stone terraces and the use of plants with potential nurse effects. We sowed and planted P. australis in a factorial design which included different degrees of protection by terraces and/or implanted vegetation types (lawns, Poa stuckertii and control without vegetation) and in top layer and deep soils exposed by gully erosion (A1 and II horizon). After 2/3 years, we evaluated survival and growth of planted P. australis, and establishment and growth of P. australis seedlings derived from the sowing. Survival of the planted P. australis was 96% without differences between treatments. The effects of the protection treatment on growth were dependant on soil depth. In the top soils the use of terraces with P. stuckertii improved seedling growth, while in deeper soils only terraces with no vegetation improved growth. Seedling establishment from the sowing experiment was very low (less than 1%) with no treatment effect, while their growth was best in terraces with P. stuckertii. Growth of seedlings was 6 times slower than the growth of planted P. australis. We conclude that the best method to implant P. australis in gullies of the Sierras Grandes is planting and not seeding. We suggest the construction of terraces with P. stuckertii as a nurse plant in the top soils and the construction of terraces without vegetation in deep soils.
Keywords: Reforestation; Facilitation; Seedling establishment; Planting.
INTRODUCCIÓN
En los ambientes muy erosionados una proporción importante del suelo se pierde como consecuencia de la formación de cárcavas (Morgan & Mngoezulu 2003). Cuando las cárcavas poseen un desarrollo incipiente su avance puede detenerse retirando los disturbios que les dieron origen (Morgan 1996), pero cuando los procesos de formación de cárcavas son avanzados, la interrupción del disturbio suele ser insuficiente para revertirlas (Bin et al. 2003). Estos casos requieren de una intervención más intensa, como, por ejemplo, la implantación de vegetación, que protege al suelo del impacto de las gotas de lluvia, disminuye su densidad aparente, y aumenta la estabilidad del suelo, la proporción de macro poros, la tasa de infiltración y retención del agua, y el contenido de materia orgánica (Morgan 1996; Cèrda et al. 1998; Bachmeier et al. 2006; Zhao et al. 2007).
La erosión de los suelos en las Sierras Grandes de Córdoba del Centro Argentino es particularmente grave. La porción más alta de estas sierras presentan roca madre expuesta por la erosión de sus suelos en casi un cuarto de su superficie (Cingolani et al. 2004). Algunos estudios sugieren que una porción importante de las Sierras Grandes (aproximadamente 30%), poseía bosques cuya canopia era dominada por Polylepis australis BITT. (tabaquillo). Estos bosques fueron degradados por acción del fuego y la ganadería, y en la actualidad ocupan alrededor de 12% de la superficie. La mayoría de estos bosques se encuentra en etapas sucesionales tempranas, y con procesos de erosión de sus suelos entre incipientes y graves (Renison et al. 2006, 2010 a,b; Cingolani et al. 2008). Este proceso de erosión reduce la capacidad de regeneración de esta especie leñosa (Renison et al. 2004, 2005; Torres et al. 2008; Zimmermann et al. 2009).
La restauración de estos ambientes degradados mediante la reforestación con especies arbóreas nativas es limitada ya que, además de P. australis, existe otra especie con posibilidades, Maytenus boaria MOLINA (Renison et al. 2010a), pero presenta dificultades para la recolección y germinación de sus semillas (Costa 2008). Experiencias previas indican que la presencia de micrositios con suelo desnudo, pendientes pronunciadas, y abundantes en arcillas y arena (como los que existen en las cárcavas del lugar), afecta de manera negativa el crecimiento de P. australis. En estas condiciones, los plantines de P. australis sólo crecen entre 4 y 6 cm de altura por año (Renison et al. 2005). Por lo tanto, una opción para su implantación en cárcavas podría ser la adecuación de la superficie de la cárcava para que retenga las semillas provenientes de árboles semilleros cercanos (o las sembradas en forma manual), y para que aumente su probabilidad de establecimiento. La implantación manual de semillas de P. australis resulta una opción viable debido a su facilidad de recolección y, en condiciones como éstas, su uso podría ser una alternativa sencilla y de bajo costos (Doust et al. 2006). Si bien una serie de estudios indican que su siembra es viable en zonas donde el suelo se ha estabilizado (Kennedy et al. 2001; Rey et al. 2005), ciertas condiciones ambientales como la luz y el calor excesivos y el stress hídrico (que son frecuentes en el interior de las cárcavas) pueden impedir la germinación de las semillas y el establecimiento de las plántulas (Torres et al. 2008). Las dificultades en el desarrollo de la vegetación dentro de las cárcavas se debe a que el suelo es escaso en materia orgánica, nitrógeno, micro y macro nutrientes, y a que su capacidad para almacenar humedad es baja (Bin et al. 2003; Burylo et al. 2007). Al mismo tiempo, los problemas de compactación también son frecuentes y dificultan el crecimiento de las raíces, lo cual restringe el número de especies vegetales que puede crecer allí (Hudson 1982). Además, la acumulación de agua de lluvia evita que la vegetación se implante (Morgan 1996; Körner 2002; Burylo et al. 2007).
Este trabajo parte de la necesidad de identificar técnicas que permitan mejorar el establecimiento a partir de semillas y el crecimiento de plantines de P. australis dentro de las cárcavas. Proponemos que estas mejoras podrían obtenerse a través de la combinación de obras de infraestructura que estabilicen el suelo y detengan los procesos de erosión (Morgan 1996; Renison et al. 2005; Burylo et al. 2007) con la implantación de vegetación adaptada a las condiciones de estrés (Castro et al. 2002; Gasque & García-Fayos 2004; Gómez-Aparicio et al. 2004; Sánchez-Velásquez et al. 2004). La sombra de la vegetación nodriza podría proteger a las semillas y a los plantines de las condiciones ambientales hostiles que se generan dentro de las cárcavas, en particular la alta temperatura y la radiación (Suzán et al. 1996; Maestre et al. 2003). A su vez, las condiciones generadas por la vegetación nodriza promoverían una mayor humedad del suelo e, incluso, protegerían de las heladas (Weid & Galen 1998; Bisigato & Bertiller 1999; Maestre et al. 2003; Caveries et al. 2005). En trabajos previos realizados en las Sierras Grandes de Córdoba se encontró que los renovales de P. australis siempre están asociados a las copas de los individuos adultos, que podrían ejercer un efecto nodriza sobre los renovales (Enrico et al. 2004; Torres et al. 2008). A su vez, en estos sitios la mayor cobertura vegetal del suelo estimula el crecimiento de los plantines (Renison et al. 2005). Por lo tanto, la evidencia local disponible apoya la idea de que el establecimiento y el desarrollo de P. australis en las cárcavas podría mejorarse mediante el uso de terrazas y de vegetación con potencial para actuar de nodriza. Los objetivos de este trabajo fueron: (1) evaluar la supervivencia y el crecimiento de plantines de P. australis en cárcavas ubicadas sobre suelos con distinto desarrollo (someros y más profundos) y bajo distintas alternativas de protección por terrazas vegetadas y sin vegetación; (2) evaluar el establecimiento a partir de semillas en relación con el de plantines, en sistemas con terraza.
MÉTODOS
Área de estudio y características de las cárcavas
El cordón de las Sierras Grandes de Córdoba es considerado una isla biogeográfica debido a su clima y biota distinta a la región Chaco Serrana que la rodea, y de la cual se sobreeleva más de 1000 m (Cabido et al. 1998). El paisaje consiste en mosaicos de vegetación que se dividen en tres grandes dominios. El primero está formado por los remanentes de bosques restringidos a afloramientos graníticos, protegidos del ganado y del fuego (Cingolani et al. 2004, 2008; Renison et al. 2006). El segundo está compuesto por pastizales de Poa stuckertii con vegas de Eleocharis albibracteata, pajonales de Deyeuxia hieronymi y céspedes de Alchemilla pinnata y Carex fuscula. Por último, encontramos los roquedales, compuestos por pajonales de Festuca tucumanica mezclados con afloramientos rocosos, afloramientos rocosos naturales con roca expuesta por erosión y pavimentos de erosión (Cingolani et al. 2004). La precipitación media anual es 854 mm, y la mayor parte (83%) ocurre entre octubre y marzo (Renison et al. 2002). La temperatura media anual es 8 ºC, y no se registran meses libres de heladas (Cabido 1985).
El área de estudio se ubica en la parte alta del macizo de Los Gigantes (2100 m.s.n.m.; 31°25' S, 64°48' W), situado en el extremo norte de las sierras. El suelo es un Hapludol éntico-lítico con desarrollo incipiente, escasa capacidad de retención de humedad y alta susceptibilidad a la erosión (Gorgas & Tassle 2003; Cingolani et al. 2008). Posee dos horizontes desarrollados: un horizonte superficial tipo A1 de color oscuro, con alto contenido de materia orgánica y textura franco-arenosa (que de ahora en más se denominará "suelo superficial"), y un horizonte II de color rojizo, con menor contenido de materia orgánica, fuertemente ácido y una mayor acumulación de arcillas (Gorgas & Tassle 2003), que en adelante se dominará "suelo profundo". El estudio se realizó en una superficie de casi 20 ha que fue alambrada para excluir al ganado y reforestada parcialmente cuatro años antes del estudio. El área posee pendientes pronunciadas de hasta 20 grados, es recorrida por un sistema de cárcavas que abarca una gran parte de la superficie, y se estima que 60% de los suelos (aproximadamente) ya se ha perdido por erosión. Debido a la protección contra el ganado se han comenzado a desarrollar pastizales densos de P. stuckertii en el fondo del valle, mientras que las laderas poseen un desarrollo incipiente. Los experimentos fueron realizados en cárcavas activas, con bordes de erosión que oscilaron entre 0.5 y 4 m de altura, y una pendiente de hasta 45 grados. Cuando los suelos son profundos, la superficie activa de la cárcava puede dejar expuesto el horizonte profundo, además del horizonte superficial.
Obtención de semillas y plantines y diseño experimental
La recolección de semillas se realizó en los meses de máxima fructificación de P. australis, entre enero y febrero de 2004. Se escogieron bosquecillos de la parte alta del macizo de Los Gigantes, cuyas semillas presentan un poder germinativo que varía entre 20 y 80% (Seltmann et al. 2007). En mayo del mismo año las semillas se sembraron en bandejas dentro de un invernadero; en julio las plántulas fueron trasplantadas a tubos individuales de 5 cm de diámetro y 15 cm de alto (ver Renison et al. 2005).
En enero de 2005 se inició el experimento con la implantación de 240 plantines sobre los bordes activos de las 14 cárcavas seleccionadas para el estudio. La altura de los plantines, desde la base a la yema apical más distante, era de 5.3 cm ±0.23 cm. Los plantines se repartieron de manera equitativa en un experimento factorial de dos factores (n=240): tipo de suelo (superficial y profundo) y protección (sin terraza, con terraza y sin vegetación, con terraza y césped, con terraza y Poa stuckertii). Cada terraza se construyó con pala retirando tierra del borde de la cárcava, dejando una superficie nivelada de 60 cm x 60 cm, la cual fue protegida con piedras en los costados. Las terrazas se ubicaron cada 3 m, y los tratamientos se seleccionaron al azar. Los panes de césped tenían un tamaño de 15 cm x 15 cm, y una altura de 10 cm, y estaban integrados por especies herbáceas (en su mayoría por Alchemilla pinnata y Carex spp.) y también por especies de otras comunidades (Cingolani et al. 2003). Los individuos de P. stuckertii trasplantados tenían una altura de 1 m. Los transplantes de césped y pajonal de P. stuckertii provinieron de sitios aledaños que se juzgaron como no susceptibles a la erosión. Luego, en marzo de 2005, se sembraron 600 semillas de P. australis dentro de cada una de las terrazas, excepto en los sitios sin terraza, ya que las semillas no eran retenidas en ellos. Las semillas se esparcieron manualmente alrededor de cada plantín y entre la vegetación implantada, y no se cubrieron con tierra. En julio de 2007 se evaluó la supervivencia y altura de los plantines y el número y la altura de las plántulas provenientes de las siembras.
Análisis estadísticos
La supervivencia de los P. australis plantados se analizó por medio de una regresión logística con variables categóricas (protección y horizonte de suelo), se utilizó la prueba "a posteriori" de Wald para detectar diferencias significativas entre tratamientos y la prueba Chi cuadrado de Pearson para determinar el ajuste del modelo (Hair et al. 1999; Quinn & Keough 2002). El crecimiento de los plantines y las plántulas se analizó por medio de un análisis de la varianza con dos factores (tipo de protección con cuatro niveles para el caso de los plantines y con tres niveles para el de las plántulas, y tipo de suelo con dos niveles) y sus interacciones. En este caso, la variable de respuesta fue el crecimiento anual calculado como el cociente entre la diferencia de las alturas de los plantines al inicio y al final del experimento y el tiempo transcurrido. Se utilizó la prueba de Duncan para detectar diferencias significativas entre medias. El número de plántulas provenientes de las siembras se analizó por medio de una regresión de Poisson con variables categóricas (protección y horizonte de suelo). Los datos se transformaron (ln) para obtener la normalidad de los residuales. Se realizó una prueba Chi cuadrado de Pearson para detectar la falta de ajuste del modelo y la prueba de Wald para detectar diferencias significativas entre los tratamientos (Hair et al. 1999; Quinn & Keough 2002). Todos los análisis estadísticos se realizaron con INFOSTAT 2008. Debido a la destrucción de terrazas y plantines por parte de visitantes, 11 plantines sin terrazas y ocho protegidos con terrazas no fueron incluidos en el análisis.
RESULTADOS
La supervivencia total de los plantines luego de tres años fue de 95.5%, y el crecimiento anual promedio fue 5.3 cm (±0.31 cm). Se contó un total de 390 plántulas vivas provenientes de las siembras, las cuales crecieron anualmente un promedio de 0.8 cm (±0.01 cm), durante las dos primeras estaciones de crecimiento. En general, la vegetación implantada de césped y P. stuckertii colocados en las terrazas sobrevivió.
La supervivencia de los plantines varió entre 89 y 96%; no fue afectada por el tipo de protección (P= 0.17, gl= 3), ni por el tipo de suelo (P= 0.81, gl= 1) y presentó un buen ajuste al modelo (χ2= 0.29). El crecimiento de los plantines reveló un efecto interactivo del tipo de protección y del tipo de suelo (Tabla 1). En ambos suelos, el uso de terraza con césped deprimió el crecimiento de los plantines con respecto a las terrazas sin vegetación; sin embargo, en el suelo superficial el uso de terrazas con P. stuckertii promovió el crecimiento de los plantines mientras que en el suelo profundo no lo afectó. Por el contrario, en este suelo la terraza sin vegetación mostró un impacto positivo significativamente superior al de las situaciones sin terraza y con terraza y césped (Figura 1).
Tabla. 1. Modelos de ANOVA para el crecimiento de plantines y plántulas.
Table 1. Models of ANOVA for the growth of seedling and plantlets.
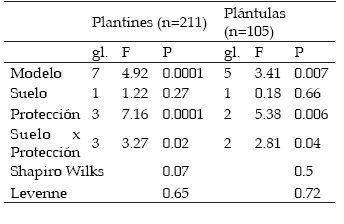

Figura 1. Crecimiento promedio anual (±error estándar) de P. australis luego de tres estaciones de crecimiento, con diferentes combinaciones de horizontes de suelo y tratamientos con terrazas de piedra y tipo de cobertura. C=control sin terraza, SV=terraza sin vegetación, Césped=terraza con césped multiespecífico, Poa=terraza con P. stuckertii. Letras diferentes indican diferencias significativas (P< 0.05).
Figure 1. Average annual growth (±standard error) of P. australis after three growing seasons, in different combinations of soil horizon and treatments with terraces and cover types. C=control, SV=terrace without vegetation, Césped=terrace with multispecific lawn, Poa=terrace with P. stuckertii tussocks. Different letters indicate significant differences (P < 0.05).
La cantidad de plántulas que sobrevivió luego de dos estaciones de crecimiento representó el 0.37% de las semillas sembradas y fue altamente variable entre parcelas. Ni el tipo de protección ni el suelo afectaron el número de plántulas presentes en terrazas con diferentes tipos de cobertura y suelo (gl= 5; vegetación P= 0.44; suelo P= 0.7; Pearson χ2= 0.18).
El crecimiento de las plántulas, evaluado solo para las 105 terrazas que presentaron plántulas mostró una interacción entre el tipo de protección y el tipo de suelo (Tabla 1, Figura 2). La interacción consistió en que la cobertura con P. stuckertii aumentó el crecimiento de las plántulas en las terrazas ubicadas en el suelo superficial, y que la vegetación con césped deprimió el crecimiento de las plántulas en las terrazas ubicadas sobre el suelo profundo (Figura 2).
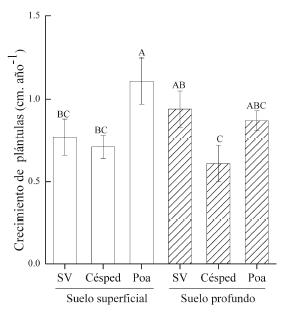
Figura 2. Crecimiento promedio anual (±error estándar) de las plántulas de P. australis luego de dos estaciones de crecimiento, con diferentes combinaciones de horizontes de suelo y tipos de cobertura. SV=terraza sin vegetación, Césped=terraza con césped multiespecífico, P=terraza con P. stuckertii Ce=césped, P=P. stuckertii. Letras diferentes indican diferencias significativas (P < 0.05).
Figure 2. Average annual growth (±standard error) of seedlings derived from sowing after two growing seasons, in different combinations of soil types and cover treatments. SV=terrace without vegetation Césped=terrace with multispecific lawn, Poa=terrace with P. stuckertii tussocks. Different letters indicate significant differences (P < 0.05).
DISCUSIÓN
Los resultados indican que la supervivencia de los P. australis que fueron plantados en cárcavas fue muy buena bajo todos los tratamientos y tipos de suelo, mientras que el crecimiento resultó superior en las terrazas con P. stuckertii para los suelos superficiales y en las terrazas sin vegetación para los suelos profundos. En cambio, las siembras repercutieron en porcentajes de establecimiento moderados pero con crecimientos posteriores excesivamente lentos.
La supervivencia de los P. australis plantados en cárcavas fue exitosa ya que 95.5% de los plantines se hallaba con vida luego de tres estaciones de crecimiento, a pesar de que fueron transplantados con solo siete meses de edad y en las condiciones ambientales hostiles que se generan dentro de las cárcavas. En plantaciones realizadas bajo condiciones similares Renison et al. (2005) documentaron supervivencias algo menores (72%), que podrían deberse a las mayores precipitaciones del presente ensayo con respecto al citado estudio (CIRSA 2008). Queda por determinar si en años secos en donde la supervivencia es menor, el micrositio más húmedo generado por P. stuckertii podría facilitar la supervivencia, como sugieren Gómez-Aparicio et al. (2004) para una situación similar donde detectaron un efecto positivo de las plantas nodriza en la supervivencia solo en los años de sequía, mientras que en los años de mayores precipitaciones el efecto positivo desaparece.
Las tasas de crecimiento promedio de los P. australis plantados en cárcavas resultaron similares a las documentadas en experiencias previas (Renison et al. 2005). En el suelo superficial, sólo se observó un efecto positivo en terrazas con P. stuckertii, que mejoró la tasa de crecimiento anual, mientras que en el suelo profundo fue el uso de terrazas sin vegetación el tratamiento que tuvo un efecto positivo. Con el crecimiento de las plántulas que surgieron de las semillas de P. australis se observó un patrón similar al anterior. Los resultados anteriores indican que el tipo de horizonte juega un papel importante para determinar el efecto de la implantación de vegetación en las cárcavas. Este patrón podría deberse a que debajo de P. stucketii se genera un microambiente más sombrío y fresco; condiciones como estas permitirían un balance hídrico más favorable para la planta (Weid & Galen 1998; Caveries et al. 2005). Este hecho, combinado con alto contenido de nutrientes del suelo superficial (Renison et al. 2010b), podría haber desencadenado el crecimiento relativamente más rápido del tratamiento con P. stuckertii. El eventual mayor contenido de humedad en el suelo debajo de P. stuckertii podría no ser aprovechable en el suelo profundo, debido a que dicho horizonte posee un mayor contenido de arcilla (Gorgas & Tassle 2003) y, por lo tanto, posee un potencial mátrico más elevado. Esto le permite retener la humedad del suelo con más fuerza (Bachmeier et al. 2006). Castro et al. (2002) obtuvieron resultados similares a los de este trabajo en especies del género Pinus, y sugieren podría deberse a cambios en la calidad del suelo. En general, el uso de césped muestra efectos negativos sobre el crecimiento de las plántulas y plantines en ambos tipos de suelo, lo que podría deberse a que, además de brindar menor protección, podrían competir por los nutrientes y la humedad del suelo con las plántulas o con el plantín.
Dos estaciones de crecimiento después de la siembra, el número de plántulas de P. australis representaba el 0.37% de las semillas sembradas y no dependió de los tratamientos. Los porcentajes de germinación de P. australis siempre son bajos, incluso bajo condiciones óptimas en el laboratorio debido a que, en promedio, sólo 10% de las semillas tienen un embrión (Renison & Cingolani 1998; Renison et al. 2004; Seltmann et al. 2007). Entonces, si suponemos que de un establecimiento máximo posible del 10% se establecieron 0.37%; ello implica que 3.7% de las semillas con potencial para establecerse lograron hacerlo. Se considera que la siembra de semillas es más eficiente que la producción de plantines, y su posterior transplante a campo, dado que se espera que sobreviva más de un 1% de las semillas (Lamb et al. 1997). Por esta razón, nuestra experiencia con P. australis estaría dentro del rango aceptable. Sin embargo, el crecimiento de las plántulas fue seis veces más lento que el de los P. australis plantados, incluso bajo los mejores tratamientos, lo cual representa una desventaja importante a la hora de reducir la erosión en las cárcavas.
El número de plántulas observado en las terrazas podría responder a variaciones en los niveles de escorrentía a nivel de micrositio. Dentro de las cárcavas, los suelos son susceptibles de ser arrastrados por la escorrentía (Morgan 1996; Rey 2003; Burylo et al 2007), y si bien las terrazas brindan una protección contra ella, durante las tormentas se detectan pequeños flujos por las terrazas. Esto provoca que las plántulas y semillas sean removidas durante el proceso erosivo (Körner 2002; Burylo et al. 2007). Al mismo tiempo, las cárcavas se caracterizan por presentar grandes variaciones en los niveles de escorrentía en pequeñas distancias (Poesen et al. 2003), lo que también contribuiría a explicar los resultados observados en las terrazas. En coincidencia con esta idea, Burylo et al. (2007) encontraron que la capacidad de retener la escorrentía por parte de las obras de infraestructuras es esencial para la colonización de cárcavas por semillas y propágulos. Por otro lado Gasque & García-Fayos (2004) observaron que Stipa tenacissima posee un efecto positivo sobre la germinación de Pinus halepensis. Sin embargo, los autores siguieren el efecto observado podría deberse a la capacidad que posee S. tenacissima para funcionar como trampa de sedimentos debido a la protección que brinda contra la escorrentía. Torres et al. (2008) encontraron que en las Sierras Grandes de Córdoba el principal factor que explica el número de renovales de P. australis a nivel de micrositio el grado de erosión. En dicho trabajo se sugiere que en sitios con erosión de suelos, las semillas son arrastradas durante las tormentas y no se retienen en sitios sin cobertura vegetal. Por todo lo expuesto, consideramos que existe un potencial para el uso de semillas de P. australis para la forestación en terrazas realizadas dentro de cárcavas. Sin embargo, para que esta técnica supere a la plantación de ejemplares producidos en vivero hará falta más investigación para encontrar métodos de acelerar el crecimiento de las plántulas.
AGRADECIMIENTOS
A la fundación Volkswagen (Alemania), al programa de "Rufford Small Grants" (Inglaterra) y proyecto PIP #11220080101458 otorgado por CONICET por el financiamiento. Al CIRSA por proveer los datos de lluvia. A los numerosos voluntarios del proyecto Conservación y Reforestación de las Sierras de Córdoba por la colaboración en los trabajos de campo. Al club Andino Córdoba por el alojamiento y apoyo al proyecto. A la Familia Nores por el permiso para realizar los experimentos en su propiedad.
BIBLIOGRAFÍA
BACHMEIER, OA; EV BUFFA; SV HANG & AA ROLLAN (eds.). 2006. Temas de edafología. Editorial Sima. Córdoba, Argentina. 250 pp. [ Links ]
BIN, Z; Y YANG & H ZEPP. 2004. Effect of vegetation restoration on soil and water erosion and nutrient losses of a severely eroded clayey Plinthudult in southeastern China. CATENA, 57:77-90. [ Links ]
BISIGATO, A & MB BERTILLER. 1999. Seedling emergence and survival in contrasting microsites in Patagonian Monte shrubland. Journal of Vegetation Science, 10:335-342. [ Links ]
BURYLO, M; F REY & P DELCROS. 2007. Abiotic and biotic factors influencing the early stages of vegetation colonization in restored marly gullies (Southern Alps, France). Ecological Engineering, 30:231-239. [ Links ]
CABIDO, MR. 1985. Las comunidades vegetales de la Pampa de Achala. Sierras de Córdoba, Argentina. Documents Phytosociologiques, 9:431-447. [ Links ]
CABIDO, MR; G FUNES; E PUCHETA; F VENDRAMINI & S DÍAZ. 1998. A chorological analysis of the mountains form Central Argentina. Is all what we call Sierra Chaco really Chaco? Contribution to the study of the flora and vegetation of the Chaco. XII. Candollea, 53:321-331. [ Links ]
CAVERIES, LA; CL QUIROZ; MA MOLINA-MONTENEGRO; AA MUÑOZ & A PAUCHARD. 2005. Nurse effect of the native cushion plant Azorella monantha on the invasive non-native Taraxacum officinale in the high-Andes of central Chile. Perspectives in Plant Ecology, Evolution and Systematics, 7:217-226. [ Links ]
CASTRO, J; R ZAMORA; JA HÓDAR & JM GÓMEZ. 2002. Use of shrubs as nurse plants: A new technique for reforestation in Mediterranean mountains. Restoration Ecology, 10:297-305. [ Links ]
CERDÀ, A. 1998. The influence of aspect and vegetation on seasonal changes in erosion under rainfall simulation on a clay soil in Spain. Can. J. Soil Sci., 78:321-330. [ Links ]
CINGOLANI, AM; M CABIDO; D RENISON & V SOLÍS NEFFA. 2003. Combined effects of environment and grazing on vegetation structure in Argentinean granitic grasslands. J. Veg. Sci., 14:223-232. [ Links ]
CINGOLANI, AM; D RENISON ; MR ZAK & M CABIDO. 2004 Mapping vegetation in a heterogeneous mountains rangeland using Landsat data: an alternative method to define and classify Land-cover units. Remote Sensing of Environment, 92:84-97. [ Links ]
CINGOLANI, AM; D RENISON ; P TECCO; D GURVICH & M CABIDO. 2008. Predicting cover types in a mountain range with long evolutionary grazing history: a GIS approach. Journal of Biogeography, 35:538-551. [ Links ]
CIRSA. Sistema para alerta tempranas de crecidas para el Río San Antonio, Córdoba. www.fi.unsj.edu.ar. [ Links ]
COSTA, C. 2008. Tratamiento pre-germinativos de Maytenus boaria. Tesis de grado. Universidad Nacional de Córdoba. [ Links ]
ENRICO, L; G FUNES & M CABIDO. 2004. Regeneration of Polylepis australis BITT. In the mountains of central Argentina. Forest Ecology and Management, 190:301-309. [ Links ]
GASQUE, M & P GARCÍA-FAYOS. 2004. Interaction between Stipa Tenacísima and Pinus halepensis: consequences for reforestation and the dynamic of the grass steppes in semiarid Mediterranean areas. Forest Ecology and Management, 189:251-261. [ Links ]
GÓMEZ-APARICIO, L; R ZAMORA; JM GÓMEZ; JA HÓDAR; J CASTRO; ET AL. 2004. Applying plant facilitation to forest reforestation: a meta analysis of the use of shrub as nurse plant. Ecological Applications, 14:1128-1138. [ Links ]
GORGAS, JA & JL TASSLE (eds.). 2003. Los suelos. Agencia Córdoba Ambiente, Córdoba, Argentina. 612 pp. [ Links ]
HAIR JR, JF; RE ANDERSON; RL TATHAM & WC BLACK. 1999. Análisis discriminante múltiple y regresión logística. [5] Pp. 249-344 en: Análisis Multivariante. 5ta ed. Prentice Hall. Madrid. [ Links ]
HUDSON, N. 1982. Erosión por cárcavas. [12] Pp. 219-239 en: Conservación de los suelos. 1 ed. Reverte S.A. España. [ Links ]
KENNEDY, A; J GILLEN; B KEETCH; C CREASER & MUTITJULU COMMUNITY. 2001. Gully erosion control at Kantju Gorge, Uluru-Kata Tjuta National park, central Australia. Ecological Management & Restoration, 2:17-27. [ Links ]
KÖRNER, C. 2002. Mountain biodiversity, its causes and function: an overview. [1] Pp. 3-20 en Körner C & Spehn EM (ed.). Mountain biodiversity. A global assessment. 1a ed. The Parthenon Publishing Group, New York, USA. [ Links ]
LAMB, D, J PARROTTA; R KEENAN & N TUCKER. 1997. Rejoining habitat remnants: restoring degraded rainforest lands. Pp. 366-385 en: Laurance, WF & RO Beirregaard. Tropical Forest Remnants: Ecology, Management and Conservation of fragmented communities. University of Chicago Press, Chicago, USA. [ Links ]
LAL, R. 2001. Soil degradation by erosion. Land Degradation and Development, 12:519-539. [ Links ]
MAESTRE, FT; J CORTINA; S BAUTISTA & J BELLOT. 2003. Does Pinus halepensis facilitate the stablishment of shrubs in Mediterranean semi-arid afforestations? Forest Ecology and Management, 176:147-160. [ Links ]
MORGAN, RPC. 1996. Erosión y conservación de suelos. Mundo prensa. Madrid. 343 pp. [ Links ]
MORGAN, RPC & D MNGOEZULU. 2003. Threshold conditions for initiation of the valley side gullies in the middle veld of Swaziland. CATENA, 50:401-414. [ Links ]
POESEN, J; J NACHTERGAELEA; G VERSTRAETENA & C VALENTIN. 2003. Gully erosion and environmental change: importance and research needs. CATENA, 50:91-133. [ Links ]
QUINN, GP & MJ KEOUGH. 2002. Generalized linear models and logistic regresión. [13] Pp. 359-379 en: Experimental Design and data analysis for biologist. 1a ed. Cambridge University press. [ Links ]
RENISON, D & AM CINGOLANI. 1998. Experiencias en germinación y reproducción vegetativa aplicados a la reforestación con Polylepis australis (Rosaceae) en las Sierras Grandes de Córdoba, Argentina. AgriScientia, 15:47-53. [ Links ]
RENISON, D; AM CINGOLANI & D SCHINNER. 2002. Optimising restoration of Polylepis australis forest through reforestation: When, where and how to transplant seedling to the mountains? Ecotrópica, 8:219-224. [ Links ]
RENISON D; I HENSEN & AM CINGOLANI. 2004. Anthropogenic soil degradation affects seed viability in Polylepis australis mountain forests of central Argentina. Forest Ecology and Management, 196:327-333. [ Links ]
RENISON, D; AM CINGOLANI; R SUAREZ; E MENOYO & C COUTSIERS. 2005. The reforestation of degraded mountain woodlands: Effects of seed provenance and microsite characteristics on Polylepis australis seedling survival and growth in central Argentina. Restoration Ecology, 13:129-135. [ Links ]
RENISON, D; I HENSEN ; R SUAREZ & AM CINGOLANI. 2006. Cover and growth habit of Polylepis woodlands and shrublands in the mountains of central Argentina: [subtitle]human or environmental influence? Journal of Biogeography, 33:876-887.
RENISON, D; I HENSEN & R SUAREZ. 2010a. Landscape structural complexity of high-mountain Polylepis australis forests: A new aspect of restoration goals. Restoration Ecology, (online first: DOI: 10.1111/j.1526-100X.2009.00555.x). [ Links ]
RENISON, D; I HENSEN; R SUAREZ; AM CINGOLANI; P MARCORA; ET AL. 2010b. Soil conservation in Polylepis mountain forests of Central Argentina: Is livestock reducing our natural capital? Austral Ecology, (online first: doi:10.1111/j.1442-9993.2009.02055.x). [ Links ]
REY, F. 2003. Influence of vegetation distribution on sediment yield in forested marly gullies. CATENA, 50:549-562. [ Links ]
SÁNCHEZ-VELÁSQUEZ, LR; S QUINTERO-GRADILLA; F ARAGÓN-CRUZ & MAR PINEDA-LÓPEZ. 2004. Nurses for Brosimum Alicastrum reintroduction in tropical dry forest. Forest Ecology and Management, 198:401-404 [ Links ]
SUZÁN, H; G NADAN & DT PATTEN. 1996. The importance of Olneya tesota as a nurse plant in the Sonoran desert. Journal of vegetation science, 7:635-644. [ Links ]
TORRES, RC; D RENISON; I HENSEN; R SUÁREZ & L ENRICO. 2008. Polylepis australis regeneration niche in relation to seed dispersal, site characteristic and livestock density. Forest Ecology and Management, 254:255-260. [ Links ]
UNDERWOOD, AJ. 1997. Analysis of variance. [7] Pp.140-197 en: Experiments in ecology: their logical design and interpretation using analysis of variance. 1a ed. Cambridge University Press. UK. [ Links ]
WEID, A & C GALEN. 1998. Plant parental care: conspecific nurse effect in Frasera speciosa and Cirsiun scopulorun. Ecology, 79:1657-1668. [ Links ]
ZIMMERMANN, H; D RENISON; I LEYER & I HENSEN. 2009. Do we need livestock grazing to promote Polylepis australis tree recruitment in the Central Argentinean Mountains? Ecological Research, 24:1075-1081. [ Links ]

