










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral v.20 n.1 Córdoba ene./abr. 2010
COMUNICACIÓNES BREVES
Estimación de la biomasa aérea de seis leguminosas leñosas del Chaco Árido (Argentina)
María del Rosario Iglesias* & Alicia Haydée Barchuk
Universidad Nacional de Córdoba, Facultad de Ciencias Agropecuarias, Cátedra de Ecología Agrícola, Córdoba, Argentina
*Universidad Nacional de Córdoba, Facultad de Ciencias Agropecuarias, Cátedra de Ecología Agrícola. Av. Valparaíso s/n, Córdoba, Argentina. Email: charo@agro.unc.edu.ar
Recibido: 3 de marzo de 2009;
Fin de arbitraje: 31 de julio de 2009;
Revisión recibida: 22 de octubre de 2009;
Aceptado: 7 de febrero de 2010
RESUMEN. La posibilidad de contar con estimaciones confiables de la biomasa aérea de la vegetación leñosa resulta imprescindible para el manejo productivo, la conservación o la restauración de los ecosistemas de bosque. Sin embargo, no siempre se dispone de técnicas relativamente sencillas y no destructivas. Este trabajo presenta modelos de regresión para estimar la biomasa aérea total de seis especies arbustivas y arbóreas de la familia Fabaceae, nativas del Chaco Árido (Prosopis flexuosa, Geoffroea decorticans, Cercidium praecox, Acacia furcatispina, Mimoziganthus carinatus y Prosopis torquata). Los modelos incluyeron distintas combinaciones del diámetro a la base (DAB) y la altura de los individuos. Todos ellos predijeron aceptablemente la biomasa por especie, si bien el de regresión lineal que utiliza el DAB2 como variable independiente presentó el mejor ajuste. Las especies fueron agrupadas según sus rasgos comunes en: monopódicas de madera dura, multitallares de madera extremadamente pesada, monopódica de madera excesivamente pesada y monopódica y multitallar de madera extremadamente pesada y para cada uno de los grupos se generó una ecuación.
Palabras clave: Modelos alométricos; Peso seco total; Densidad básica de leño; Caducifolias; Bosques xerofíticos.
ABSTRACT. Estimating aerial biomass of six woody Leguminosae of the Arid Chaco (Argentina): Reliable estimates of biomass for the vegetation of the Arid Chaco forest is critical for its management, conservation or restoration. However, simple and non destructive techniques are not largely available. This study provides regression models aimed at estimating the aerial biomass of native shrub and tree species from the Arid Chaco, belonging to the Fabaceae family (Prosopis flexuosa, Geoffroea decorticans, Cercidium praecox, Acacia furcatispina, Mimoziganthus carinatus and Prosopis torquata). An additional objective was to explore the feasibility of using a single equation for several species. The wood density differed significantly between species. Linear and nonlinear regression models of aerial biomass were adjusted for species, with different combinations of base diameter (DAB) and height as independent variables. The models properly predicted the biomass by species, the most appropriate being the linear regression model using DAB2 as an independent variable. To establish differences in the behavior of species and, more precisely, in the ratio between total dry weight and DAB, the slope homogeneity was analyzed using a regression model with auxiliary variables. Taking into account the results obtained and the basic wood density and morphological growth structure, four groups of species have been postulated, and it was feasible to apply a single equation per group. The proposed groups are: (1) monopodial hardwood (Geoffroea decorticans and Cercidium praecox), (2) multi-stem shrubs of extremely heavy wood (Acacia furcatispina and Mimoziganthus carinatus), (3) monopodial of very heavy wood (Prosopis flexuosa) and (4) monopodial and multi-stem of extremely heavy wood (Prosopis torquata).
Keywords: Allometric models; Aerial biomass; Basic wood density; Deciduous; Xerophytic forests.
INTRODUCCIÓN
La estimación de la biomasa de especies leñosas resulta indispensable para conocer la productividad primaria neta (Návar-Cháidez & González-Elizondo 2009), la circulación y la magnitud de los reservorios de nutrientes (Abrams et al. 1990; Ordóñez & Masera 2001; Zabek & Prescott 2006), caracterizar la estructura de la vegetación de bosques nativos y forestaciones (Cuevas et al. 1991; Ketterings et al. 2001), estimar la carga de combustible forestal para modelos de prevención de fuego (Melo et al. 2006), comparar características de especies (Felker et al. 1982; Paton et al. 2002; Segura & Kanninen 2005) y comunidades similares (Gaillard de Benítez et al. 2000).
Los modelos predictivos de biomasa fueron ampliamente desarrollados para especies de ecosistemas mésicos y plantaciones forestales (Segura & Kanninen 2005; Fonseca et al. 2009), pero son más escasos para bosques xerofíticos (Gayoso et al. 2002; León de la Luz et al. 2005), en particular para aquellas de regiones como el Chaco Árido (Gaillard de Benítez et al. 2000) y la provincia fitogeográfica del Monte (Hierro et al. 2000), donde dominan los árboles de porte pequeño y especies arbustivas con estructuras multitallares (Gurvich et al. 2005).
Por lo general, los modelos son generados sobre la base de una especie en particular. Sin embargo, es frecuente observar que las especies de una misma formación vegetal presentan similitudes en su morfología y en la asignación de biomasa aérea. Esto permitiría generar un único modelo predictivo para estimar la biomasa de un conjunto de especies a partir de variables alométricas comunes (Acosta-Mireles et al. 2002). Los modelos de regresión para predecir la biomasa aérea por especie utilizan variables alométricas medidas en los individuos a campo y cuya evaluación puede ser o no destructiva (Segura 1997; Segura & Andrade 2008). El diámetro del fuste a la altura del pecho (1.3 m) es la variable alométrica independiente más adecuada por su fácil medición y alta correlación con la biomasa (Chambouleyron & Brown 1992; Ter-Mikaelian & Korzukhin 1997; Specht & West 2003; Fonseca et al. 2009), si bien sólo resulta conveniente para individuos con fuste alto. Otras combinaciones tales como el producto del diámetro a la altura de pecho y la altura (Husch 2001) o simplemente el diámetro a la base, resultan apropiadas para individuos jóvenes o de tamaño pequeño (Loguercio & Dossé 2001; Pérez-Cordero & Kanninen 2002), aunque su uso no está muy difundido en la actualidad.
La región del Chaco Árido (Argentina) presenta una de las mayores superficies con vegetación leñosa del país (Gasparri & Manghi 2004). En esta región, las leguminosas suelen poseer fustes cortos y multitallares, y representan la mayor proporción de la biomasa, en especial en aquellos ambientes con cierto grado de perturbación (Cabido et al. 1992; Barchuk et al. 2006). Estas especies resultan relevantes para la conservación de carbono (Asner et al. 2003), la producción de forraje (Guillen Trujillo et al. 2007) y la fijación de nitrógeno en el suelo (Ayala 2005). Este trabajo provee modelos individuales para estimar la biomasa aérea de seis especies leñosas leguminosas representativas de la región del Chaco Árido, y modelos que permiten agruparlas de acuerdo a su morfología y densidad de leño.
MATERIALES Y MÉTODOS
La calibración de los modelos se realizó en Las Toscas, al norte de la provincia de Córdoba, Argentina (29º30´ S, 64º11´ O), en un sector del Chaco Árido de la provincia fitogeográfica Chaqueña (Cabrera 1976). El clima es árido, con precipitación media anual de 500 mm y temperatura media anual de 18 ºC. La vegetación del área de estudio es un bosque xerofítico secundario de Aspidosperma quebracho-blanco y Prosopis flexuosa, caracterizado por una proporción alta de especies arbustivas de la familia Fabáceas.
Se seleccionaron seis especies sobre la base de su importancia y representatividad en la región: Prosopis flexuosa, Geoffroea decorticans, Cercidium praecox, Acacia furcatispina, Mimozyganthus carinatus y Prosopis torquata. Para cada especie se realizó un muestreo de entre 19 y 25 individuos, comprendidos en un rango amplio de altura y diámetro a la base. Para cada individuo se registró su diámetro a la base (DAB) a los 30 cm del suelo y su altura. La estimación de biomasa fue destructiva. El peso húmedo total de los individuos cortados desde la base (incluyendo ramas y hojas) fue determinado a campo por medio de una balanza tipo báscula (precisión 0.25 kg). En el caso de las especies multitallares, se consideró cada módulo de la estructura multitallar como un individuo. De cada individuo se extrajeron cinco rodajas de aproximadamente 5 cm de espesor (tres del fuste y base de la copa y dos de la copa). Las rodajas fueron pesadas a campo con una balanza de precisión 0.1 g (peso húmedo), y luego fueron secadas en estufa (60 ºC) hasta peso constante (Brown et al. 1995). A partir del contenido de humedad de estas submuestras se estimó el peso seco total de cada individuo. Se calculó la densidad básica del leño como la relación entre el peso seco de la rodaja y su volumen saturado. Se consideraron 14 rodajas por especie, obtenidas a partir de un submuestreo estratificado de 121 rodajas, abarcando los diferentes rangos diamétricos existentes entre los individuos apeados. Una vez obtenido el peso seco de cada una, se las colocó en agua hasta que alcanzaran valores de humedad saturada constante. Luego, se determinó el volumen saturado de cada rodaja por el método de inmersión y desplazamiento de agua (Méndez González et al. 2005).
La densidad de leño de las seis especies se comparó a través de un análisis de la varianza de una vía, seguida "a posteriori" por una prueba de Fisher para comparaciones múltiples. Se exploraron los modelos predictivos de la biomasa a partir de modelos de regresión lineales y modelos no lineales, donde se utilizó el peso seco como variable dependiente y DAB, altura total, DAB2 y el producto del DAB2por la altura, como variables independientes. Se utilizaron modelos de regresión simple [Y= ß0 + ß? (X)], y la función potencial (Y= ß0 Xß1), linealizada mediante transformación logarítmica de ambas variables, dependiente e independiente, cuya expresión matemática es: ln Y= ln ß0 + ß1 ln (X). Este modelo, también llamado logarítmico (Paton et al. 2004), presenta los términos ln (Y), ln ß0 y ln (X) que corresponden a los logaritmos naturales de Y, ß0 y X. En este caso, las variables independientes utilizadas fueron DAB y altura total. Los valores de ß0 y ß1, corresponden a la ordenada al origen y pendiente, respectivamente. En todos los casos se verificaron los supuestos de distribución normal del error experimental, homogeneidad de varianza e independencia de los errores (Sokal 1994).
Con el fin de encontrar un único modelo que permitiera estimar la biomasa de más de una especie se compararon las pendientes (ß1) de los modelos de regresión lineal simple de las seis especies, con el DAB como variable independiente, mediante un modelo de regresión con variables auxiliares (InfoStat 2008). Se generaron grupos de especies a partir de: (1) los contrastes realizados en los ANAVA con variables auxiliares (Montgomery 1991), (2) prueba "a posteriori" de Fisher para las diferentes densidades de leño y (3) la estructura morfológica de crecimiento de las especies. Por último, para cada grupo se probó el ajuste mediante un modelo de regresión lineal simple con el DAB como variable independiente. Todos los análisis estadísticos se realizaron utilizando el programa InfoStat (2007).
RESULTADOS
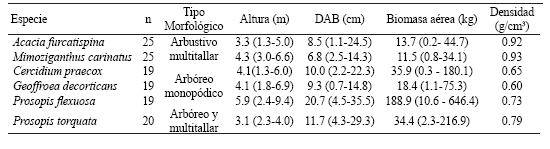
La variación entre los tamaños medios y las estructuras morfológicas de las seis especies estudiadas permitió diferenciar árboles monopódicos de pequeño a mediano porte (Geoffroea decorticans y Cercidium praecox), arbustos multitallares (Acacia furcatispina y Mimoziganthus carinatus), árboles de gran porte con valores elevados de biomasa aérea (Prosopis flexuosa) y Prosopis torquata que presenta tanto fustes monopódicos como multitallares (Tabla 1). El análisis de la densidad de leño mostró tres grupos diferenciados de densidad creciente (P<0.05): (1) C. praecox y G. decorticans, (2) P. flexuosa y P. torquata, y (3) A. furcatispina y M. carinatus (Tabla 1). Estos grupos se formaron siguiendo la clasificación de maderas propuesta por Markwar y Meck (Valencia & López 1999) como (1) pesadas, (2) excesivamente pesadas, y (3) extremadamente pesadas.
Tabla 1. Caracterización de seis especies de Fabáceas caducifolias del Chaco Árido. Número de muestras (n); tipo morfológico; valores medios, máximos y mínimos de altura, diámetro a la base (DAB), biomasa aérea seca y valores medios de densidad de leño.
Table 1. Characterization of six woody, deciduous Fabaceae of the Arid Chaco. Number of samples (n); morphological type (multi-stem shrub, single-stem tree, multi-stem tree); average, maximum and minimum height; basal diameter (DAB); dry aerial biomass, and mean wood density.
Los modelos individuales para cada especie presentados tuvieron coeficientes de determinación mayores a 0.7 (Tabla 2). En general, el ajuste de los de regresión lineal simple y el logarítmico resultaron altamente significativos (P<0.0001) y con valores bajos de error estándar de la estimación. En los modelos de regresión simple con variables independientes DAB ó DAB2, los coeficientes determinación (R2) fueron superiores a 0.6. Cuando la variable independiente fue el producto de DAB2 y altura, los R2 alcanzaron valores superiores a 0.8. Por último, los modelos logarítmicos también presentaron un ajuste altamente significativo (Tabla 2). Cuando se utilizó la altura como variable independiente, los modelos presentaron un ajuste altamente significativo pero los coeficientes de determinación resultaron mucho menores (Tabla 2).
Tabla 2. Modelos de regresiones lineales simples y logarítmicas para la estimación del peso seco total individual con valores de coeficientes de determinación mayores a 0.7. Coeficiente de determinación (R2) y error estándar de la estimación de la ordenada al origen (EE) de las seis especies caducifolias Fabáceas del Chaco Árido. Todos los modelos presentaron fueron significativos (P<0.0001).
Table 2. Fitted functions for dry individual weight as a function of basal diameter (DAB), coefficient of determination (R2) and standard error of y-intercept (EE) of six woody deciduous Fabaceae from the Arid Chaco, with R2 greater than 0.7. All models were significant (P<0.0001).

El análisis de la homogeneidad de pendientes mostró que P. flexuosa, de porte superior a las demás, se diferenció del resto (P<0.0001, R2=0.76). Un segundo análisis, que excluyó a P. flexuosa, presentó un alto coeficiente de determinación (R2=0.83) y mostró diferencias significativas entre las especies restantes (P<0.01) (Tabla 3, derecha). P. torquata se diferenció de C. praecox, G. decorticans, A. furcatispina y M. carinatus. El análisis de las pendientes de los modelos que tenían el DAB como variable independiente mostró que G. decorticans y C. praecox no se diferenciaron significativamente entre sí, al igual que A. furcatispina y M. carinatus (Tabla 4).
Tabla 3. Modelos de regresión lineal con variables auxiliares y probabilidad (P) de seis especies de Fabáceas (izquierda) y de cinco especies (derecha). Peso seco estimado (PS) y probabilidad de que las diferencias encontradas entre las pendientes hayan sido por azar (valor P).
Table 3. Simple linear regression with dummy variables for six (left) and five (right) woody, deciduous Fabaceae from the Arid Chaco. Estimated dry weight (PS) and probability (P)

Tabla 4. Tabla de contrastes entre especies (excluyendo Prosopis flexuosa) a partir de ANAVA con variables auxiliares; nivel de significancia=0.01.
Table 4. Contrasts between five species (excluding Cercidium flexuosa) from ANOVA with dummy variables (P< 0.01).

Los resultados obtenidos permitieron proponer los siguientes grupos de especies: (G1) monopódicas de madera pesada (G. decorticans y C. praecox); (G2) multitallares arbustivas de madera extremadamente pesada (A. furcatispina y M. carinatus); (G3) monopódica de madera excesivamente pesada (P. flexuosa) y (G4) monopódica y multitallar de madera extremadamente pesada (P. torquata). Para los grupos G1 y G2 (G1 con 38 individuos y G2 con 50 individuos) se obtuvieron modelos de regresión lineal con el DAB como variable independiente. En ambos casos, los modelos ajustaron de manera altamente significativa, con coeficientes de determinación superiores a 0.82 (Tabla 5). Cuando se formularon los modelos lineales simples para los cuatro grupos establecidos se observó que el valor de la ordenada al origen (b0) del grupo G2 (arbustivas multitallares M. carinatus y A. furcatispina) fue mayor que en el resto de los grupos. En cambio, los valores estimados de las pendientes (b1) fueron superiores en las monopódicas, principalmente P. flexuosa, donde los individuos son de mayor tamaño y peso. Aplicando dicho modelo de regresión, se observó que a diámetros basales pequeños las especies multitallares (G2) presentaron valores mayores de peso seco total, en comparación al resto (Figura 1). Al aumentar el DAB se revierte esta situación ya que a diámetros mayores las especies leñosas monopódicas (G1 y G3) y P. torquata (G4), presentaron mayor biomasa. Por último, dentro del género Prosopis, ambas especies presentaron distintos patrones (Figura 1): a diámetros basales pequeños, P. torquata tuvo valores mayores de peso seco total que P. flexuosa, que tiene a una densidad de leño mayor.
Tabla 5. Funciones ajustadas de la forma de PS= ß0+ ß1 * DAB, donde PS es peso seco total individual y DAB es el diámetro basal, para los cuatro grupos de especies: G1 (Geoffroea decorticans y Cercidium praecox) y G2 (Mimoziganthus carinatus y Acacia furcatispina), G3: Prosopis flexuosa y G4: Prosopis torquata. También se muestran los valores de R2.
Table 5. Fitted functions of the form PS= ß0+ ß1 * DAB, where PS is individual dry weight and DAB is basal diameter for four groups of woody deciduous Fabaceae from the Arid Chaco. P values for all functions <0.0001. R2 values are shown.

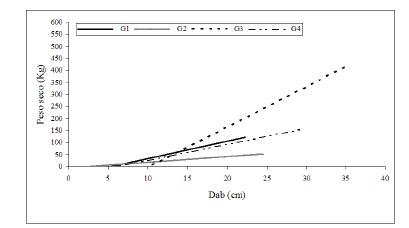
Figura 1. Relación entre biomasa total y diámetro basal (DAB) para los cuatro grupos de especies analizadas: G1: Geoffroea decorticans y Cercidium praecox; G2: Mimoziganthus carinatus y Acacia furcatispina; G3: Prosopis flexuosa; G4: Prosopis torquita. Para valores de los parámetros de las funciones ajustadas ver Tabla 5.
Figure 1. Relationship between aboveground biomass and basal diameter (DAB) for the four groups of species. G1: Geoffroea decorticans and Cercidium praecox; G2: Mimoziganthus carinatus and Acacia furcatispina; G3: Prosopis flexuosa; G4: Prosopis torquata. For parameters of the fitted function see Table 5.
DISCUSIÓN Y CONCLUSIONES
Este trabajo provee funciones simples para predecir la biomasa aérea de seis especies leguminosas caducifolias de los bosques del Chaco Árido. Además, se evidenció que a partir de las similitudes entre las especies, es factible utilizar un mismo modelo para un grupo con características comunes y así simplificar la estimación de biomasa leñosa aérea de distintas especies, dentro de un rango de tamaños similar al explorado.
Muchos estudios sobre modelos predictivos de estimación de biomasa por especie indican la superioridad de la función potencial (Verwijst 1991; Ketterings et al. 2001; Acosta-Mireles et al. 2002; Sampaio & Silva 2005). En nuestro trabajo, tanto dicha función linealizada (modelo logarítmico) con el DAB como variable independiente, como el modelo regresión lineal simple que utiliza el producto entre el DAB2 y la altura, fueron los que mostraron mejor ajuste a nivel especie (coeficiente de determinación superior a 0.8). De esta manera, para estimar la biomasa de Prosopis flexuosa y Geoffroea decorticans se recomienda este último modelo, ya que no es necesario transformar las variables a logaritmo. La altura de los individuos no resultó un aporte significativo para la estimación de la biomasa del resto de las especies, lo que evitaría la necesidad de obtener este dato que frecuentemente es de difícil registro, principalmente en los bosques cerrados (Segura & Kanninen 2005).
Al igual que en algunos trabajos previos (Acosta-Mireles et al. 2002; Sah et al. 2004), las estimaciones basadas en especies mostraron mejor ajuste que aquéllas estimadas para grupos con características similares, sin embargo estas últimas son ampliamente utilizadas para cálculos de biomasa forestal, especialmente cuando algunas de las estimaciones específicas no están disponibles (Sah et al. 2004; Padrón & Navarro-Cerrillo 2004). En el caso particular de la familia Fabaceae existen antecedentes que plantean la utilización de un único modelo para la descripción de esta familia. Felker et al. (1982) a partir de la similitud en ocho especies juveniles de leguminosas (nueve meses de edad), entre ellas tres de Prosopis, plantea la posibilidad de considerar una misma ecuación para estimar la biomasa. Por otro lado, Padrón y Navarro-Cerrillo (2004), a falta de ecuaciones específicas en un estudio de la estimación de biomasa en bosques secos de Perú dominados por Prosopis pallida, aplican a todos los miembros de la familia Fabaceae presentes en el ecosistema (entre ellas, Cercidium praecox) la misma ecuación generada para P. pallida. Este estudio sugiere que no se debería utilizar una única ecuación de estimación de biomasa para los distintos miembros de la familia de leguminosas, ya que presentan diferencias notorias en su morfología, densidad de leño y acumulación de biomasa por unidad diamétrica. No obstante, fue posible identificar entre las seis especies, cuatro patrones distintos, definidos cada uno por una única ecuación, que simplificaría la estimación de biomasa forestal.
AGRADECIMIENTOS
Este trabajo recibió financiamiento de la Agencia Nacional de Promoción Cientifica y Técnica Argentina (FONCYT, proyecto "El secuestro de carbono y el papel ecológico de las especies leñosas en un gradiente de déficit hídrico en la zona árida - semiárida argentina", PICTR 2002-00200). La valiosa colaboración de C. Bruno, J.L. Zamar, S. Katlirevsky, D. Sercovich, F. de Rosa, S. Luque, E. Alessandria y M. Boetto.
BIBLIOGRAFÍA
ABRAMS, MM; WM JARRELL; HA SMITH & PR CLARK. 1990. Nitrogen accretion in soil and biomass production by three Prosopis species. Agroforestry Systems, 10:93-97. [ Links ]
ACOSTA-MIRELES, M; A VARGAS-HERNÁNDEZ; A VELÁSQUEZ-MARTÍNEZ & JD ECHEVERS-BARRA. 2002. Estimación de la biomasa aérea mediante el uso de relaciones alométricas en seis especies arbóreas en Oaxaca. México. Agrociencia, 36:725-736. [ Links ]
ASNER, GP; CE BORGHI & R OJEDA. 2003. Desertification in central Argentina: changes in ecosystem carbon and nitrogen from imaging spectroscopy. Ecological Applications, 13(3):629-648. [ Links ]
AYALA, LB. 2005. Estudios de algunos aspectos de la fijación simbiótica de nitrógeno por el maní (Arachis hypoagea). II. Evaluación bioquímica de la fijación y factores relacionados en la asociación maní-rhizobium spp. Agronomía Tropical, 27:427-449. [ Links ]
BARCHUK, AH; E CAMPOS; C OVIEDO & P DÍAZ. 2006. Supervivencia y crecimiento de plántulas de especies leñosas del Chaco Árido sometidas a remoción de la biomasa aérea. Ecología Austral, 16:47-61. [ Links ]
BROWN, IF; LA MARTINELLI; WW THOMASD; MZ MOREIRA; CA CID FERREIRA ET AL. 1995. Uncertainty in the biomass of Amazonian forests: An example from Rondonia, Brazil. Forest Ecology and Management, 75:175-189. [ Links ]
CABIDO, M; A ACOSTA; ML CARRANZA & S DÍAZ. 1992. La vegetación del Chaco Árido en el W de la Provincia de Córdoba; Argentina. Documents Phytosociologiques, 14:447-456. [ Links ]
CABRERA, AL. 1976. Regiones Fitogeográficas Argentinas. Pp. 1-85 en: Kugler, WF (ed.), Enciclopedia Argentina de Agricultura y Jardinería. Segunda edición. Editorial Acme S.A.C.I, Buenos Aires, Argentina. [ Links ]
CHAMBOULEYRON, M & W BRAUN. 1992. Producción primaria neta de tres leguminosas del piedemonte mendocino. Multequina, 1:65-71. [ Links ]
CUEVAS, E; S BROWN & AE LUGO. 1991. Above and below ground organic matter storage and production in a tropical pine plantation and a paired broadleaf secondary forest. Plant Soil, 135:257-268. [ Links ]
FELKER, PR; JF OSBORN & GH CANNEL. 1982. Biomass estimation in a young stand of mesquite (Prosopis spp.), ironwood (Olneya tesota), palo verde (Cercidium floridum and Parkinsonia aculeata) and leucaena (Leucaena leucocephala). J. Range Manage., 35:87-89. [ Links ]
FONSECA, WG; FG ALICE & JM REY. 2009. Modelos para estimar la biomasa de especies nativas en plantaciones y bosques secundarios en la zona Caribe de Costa Rica. Bosque, 30(1):36-47. [ Links ]
GAILLARD DE BENÍTEZ, C; M PECE; M JUÁREZ DE GALÍNDEZ; A MALDONADO; VH ACOSTA ET AL. 2000. Biomasa aérea de quebracho blanco (Aspidosperma quebracho-blanco) en dos localidades del Parque Chaqueño seco. Quebracho, 9:115-127. [ Links ]
GASPARRI, I & E MANGHI. 2004. Estimación de volumen. Biomasa y contenido de carbono de las regiones forestales argentinas. Unidad de Manejo del Sistema de Evaluación Forestal. Ministerio de Salud y Ambiente. Consultado 22 mar. 2010. Disponible en www.ambiente.gov.ar/archivos/web/UMSEF/File/volumen_biomasa_carbono.pdf. [ Links ]
GAYOSO, J. 2002. Medición de la capacidad de captura de carbono en bosques de Chile y promoción en el mercado mundial. Universidad Austral de Chile. Valdivia, Chile, 53 pp. [ Links ]
GUILLEN TRUJILLO, A; A PALACIOS ESPINOSA & JL ESPINOZA VILLAVICENCIO. 2007. Ecuaciones de predicción para la producción de forrajes de Palo verde (Cercidum floridum Bent H. ex A. Gra Y) en baja California Sur, México. Interciencia, 32:712-715. [ Links ]
GURVICH, D; L ENRICO & A CINGOLANI. 2005. Linking plant functional traits with post-fire sprouting vigour in woody species of central Argentina. Austral Ecology, 30:789-796. [ Links ]
HIERRO, JL; L BRANCH; D VILLARREAL & K CLARK. 2000. Predictive equations for biomass and fuel characteristics of Argentine shubs. J. Range Manage., 53:617-621. [ Links ]
HUSCH, B. 2001. Estimación del contenido de carbono de los bosques. Simposio Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales.Valdivia, Universidad Austral de Chile, 9 pp. [ Links ]
INFOSTAT 2007. Grupo Infostat. FCA, Universidad Nacional de Córdoba. Versión 2007. Argentina. [ Links ]
INFOSTAT 2008. Manual del usuario. Versión 2008. Grupo Infostat. FCA, Universidad Nacional de Córdoba. Argentina. Primera Edición. Editorial Brujas Argentinas. [ Links ]
KETTERING, QM; R COE; M VAN NOORDWJK; Y AMBAGAU & CA PALM. 2001. Reducing uncertainty in the use of allometric biomass equations for predicting aboveground tree biomass in mixed secondary forest. Forest Ecology and Management, 146:199-209. [ Links ]
LEÓN DE LA LUZ, JL; R DOMÍNGUEZ & S DÍAZ CASTRO. 2005. Evaluación de un análisis dimensional para predecir peso fresco del leño en dos especies de mezquite, Prosopis articulata S. Watson y P. palmeri S. Watson. Acta Botánica Mexicana, 72:17-32. [ Links ]
LOGUERCIO, GA & GE DEFOSSÉ. 2001. Ecuaciones de biomasa aérea. Factores de expansión y de reducción de la lenga Nothofagus pumilio (poepp. et endl.) Krasser, en el SO del Chubut. Argentina. Simposio Internacional Medición y Monitoreo de la Captura de Carbono en Ecosistemas Forestales, 11 Pp. [ Links ]
MELO, LAMN; RV SOARES; JDP SIQUEIRA & FF KIRCHNER. 2006. Modelagem de combustiveis florestais no Parque Nacional do Iguacu, Paraná, Brasil. Floresta, 36(3):415-424. [ Links ]
MÉNDEZ GONZÁLEZ, J; F MORALES CARREDANO; VJ RUIZ GARCÍA & JA NÁJERA LUNA. 2002. Ajuste de modelos para estimar biomasa fustal en Pinus cooperi y P. leiophylla, de la región del Salto; Durango, México. Agrociencia, 36:725-736. [ Links ]
MOGLIA, M & G JOFRÉ. 1998. Response to clearing of woody plants in a forest of quebracho blanco (Aspidosperma quebracho-blanco) and algarrobo (Prosopis flexuosa) in the Province of San Luis (Argentina). Pitón, 63:257-265. [ Links ]
MONTGOMERY, DC. 1991. Diseño y Análisis de Experimentos. Grupo Editorial Iberoamérica. [ Links ]
NÁVAR-CHÁIDEZ, JJ & S GONZÁLEZ-ELIZONDO. 2009. Diversidad, estructura y productividad de bosques templados de Durango, México. Polibotanica, 27:71-87. [ Links ]
ORDÓÑEZ, JAB & O MASERA. 2001. Captura de carbono ante el cambio climático. Maderas y Bosques, 7(1):3-12. [ Links ]
PADRÓN, E & RM NAVARRO CERRILLO. 2003. Above-Ground Biomass in Natural Populations of Prosopis pallida HBK in the North Peru. Journal of Arid Environmental, 56:283-292. [ Links ]
PATON, D; J NÚÑEZ; D BAO & A MUÑOZ. 2002. Forage biomass of 22 shrub species from Monfragüe Natural Park (SW Spain) assessed by log-log regression models. Journal of Arid Environments, 52(2):223-231. [ Links ]
PÉREZ CORDERO, LD & M KANNINEN M. 2002. Wood specific gravity and aboveground biomass of Bombacopsis quinata plantations in Costa Rica. Forest Ecology and Management, 165:1-9. [ Links ]
SAH, JP; MS ROSS; S KOPTUR & JR ZINDER. 2004. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida Keys pine forests. Forest Ecology and Management, 203:319-329. [ Links ]
SAMPAIO, EVSB & GC SILVA. 2005. Biomass equations for Brazilian semiarid caatinga plants. Acta Bot. Brasilica, 19:935-943. [ Links ]
SEGURA, M & H ANDRADE. 2008. ¿Cómo hacerlo? ¿Cómo construir modelos alométricos de volumen, biomasa o carbono de especies leñosas perennes? Agroforestería de las Américas, 46:89-96. [ Links ]
SEGURA, M & M KANNINEN. 2005. Allometric Models for Tree Volume and Total Aboveground Biomass in a Tropical Humid Forest in Costa Rica. Biotropica, 37(1):2-8. [ Links ]
SEGURA, M. 1997. Almacenamiento y fijación de carbono en Quercus costarricensis, en un bosque de altura en la cordillera de Talamanca, Costa Rica. Tesis de Licenciatura, Universidad Nacional. Heredia, Costa Rica. Escuela de Ciencias Ambientales, 126 pp. [ Links ]
SOKAL, RE. 1994. Biometry. Ed. WH Freeman and Company, 887 pp. [ Links ]
SPECHT, A & PW WEST. 2003. Estimation of biomasa and sequesteres carbono in faro forest plantations in northen New South Wales, Australia. Biomass and Bioenergy, 25:363-379. [ Links ]
TER-MIKAELIAN, MT & MD KORZUKHIN. 1997. Biomass equation for sixty-five North American tree species. For. Ecol. Manage., 97:1-24. [ Links ]
VALENCIA, MS & FA LÓPEZ. 1999. Variación de la densidad de la madera dentro y entre árboles de Pinus rudis Endl.; en Sierra Las Alazanas, Arteaga, Coah. Foresta-AN. Nota Técnica No. 1. UAAAN. Saltillo, Coah, 17 pp. [ Links ]
VERWIJST, T. 1991. Logarithmic transformations in biomass estimation procedures: violation of the linearity assumption in regression analysis. Biomass and Bioenergy, 1:175-180. [ Links ]
ZABEK, LM & CE PRESCOTT. 2006. Biomass equations and carbon content of aboveground leafless biomass of hybrid poplar in coastal British Columbia. Forest Ecology Management, 223:291-302. [ Links ]

