










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.20 no.3 Córdoba Sept./Dec. 2010
TRABAJOS ORIGINALES
Composición y diversidad del sotobosque de ñire (Nothofagus antarctica) en función de la estructura del bosque
Pamela Quinteros1,*, Nidia Hanzen2 & Adriana Kutschker3
1 Centro de Investigación y Extensión Forestal Andino Patagónico (CIEFAP). Esquel, Chubut.
2 Instituto Nacional de Tecnología Agropecuaria (INTA), EEA-Esquel. Esquel, Chubut.
3 Universidad Nacional de la Patagonia San Juan Bosco. Esquel, Chubut.
* Centro de Investigación y Extensión Forestal Andino Patagónico (CIEFAP). Ruta 259, km 4, (9200) Esquel, Chubut. Email: pquinteros@ciefap.org.ar.
Recibido: 22 de marzo de 2010;
Fin de arbitraje: 21 de junio de 2010;
Revisión recibida: 6 de julio de 2010;
Aceptado: 27 de julio de 2010
RESUMEN. Este trabajo describe la composición específica y la diversidad del sotobosque de Nothofagus antarctica (ñire) en función de la estructura del bosque. El estudio se realizó en el noroeste de Chubut, donde se clasificó al bosque en 10 tipos forestales según la altura y la cobertura. Realizamos censos de la vegetación que nos permitieron investigar los diferentes tipos forestales en términos de su riqueza específica, diversidad, dominancia y cobertura del sotobosque (total y según origen y hábito). Se registraron 105 especies (73% nativas y 27% exóticas) distribuidas en 43 familias. En todas las estructuras analizadas se registró una elevada cobertura de especies herbáceas. En los tipos forestales de mayor altura se registró la mayor cobertura total, que bajo dosel denso estuvo compuesta por especies nativas propias del bosque, en tanto que en bosques abiertos se registró una mayor cobertura de especies exóticas, típicas de ambientes alterados. En los bosques bajos, donde coexisten especies de ambiente altoandino, estepa, alterado y bosque, se registró mayor diversidad y riqueza de especies.
Palabras clave: Cobertura; Composición florística; Ñirantal; Patagonia; Tipos forestales.
ABSTRACT. Composition and diversity of the understory of ñire (Nothofagus antarctica) forests in relation to forest structure: We describe the species composition and diversity of the understory of Nothofagus antarctica (ñire) forest depending on the forest structure. The study was conducted in the northwest of Chubut, where the forest was classified into 10 forest types, taking into account the forest height and the canopy cover. On the basis of vegetation inventory, we studied the differences among forest types in terms of species richness, diversity, dominance and understory cover (total and by origin and life form). We recorded 105 species (73% native and 27% exotic) distributed in 43 families. In all the structures analyzed high coverage of herbaceous species was recorded. In the forest type with the highest tree-height the highest coverage were recorded, composed of native species typical of the forest under dense canopies, and of exotic species, typical of disturbed environments, in open forests. In the forest type with the lowest tree-height, where species of the forest, of disturbed environments, and of the steppe coexist, there was greater diversity and species richness.
Keywords: Cover; Floristic composition; Ñirantal; Patagonia; Forest types.
INTRODUCCIÓN
Los bosques de ñire representan el 33% del total de bosques en la Provincia de Chubut (Bava et al. 2008) y en muchas zonas de Patagonia constituyen las comunidades forestales con mayor intensidad de explotación (Montaña 1982). El uso ganadero tradicional en estos sistemas es extensivo y continuo (Manacorda et al. 1995; Hansen et al. 2004; Reque et al. 2007) y la concentración más elevada del stock bovino (>70% de la provincia) se registra en las áreas cordillerana, precordillerana y en el ecotono con la estepa (Guitart 2004). A su vez, la tortuosidad de los fustes y el mal estado sanitario de los individuos restringen el uso silvícola a la extracción más o menos intensa de leña (Manacorda et al. 1995). Las cortas de intensidad elevada son cada vez menos comunes debido a la mayor producción forrajera de estos sistemas con cobertura arbórea (Peri et al. 2005) y a la vigencia de normas que las prohiben (DGByP 2004).
La vegetación de los estratos inferiores del bosque (sotobosque) constituye la base de las redes alimentarias y el refugio de gran parte de la micro y mesofauna, y sirve de protección al suelo contra la erosión (Ellum 2009). Las características del sotobosque están muy asociadas a la cobertura del dosel que regula diversos procesos físicos como la intensidad de la radiación solar, la intercepción de las precipitaciones y la exposición al viento (Holmgren et al. 1997; Valladares et al. 2004); las prácticas silviculturales que generan apertura del dosel modifican estos procesos, influyendo sobre la cobertura y la estructura del sotobosque (Klinka et al. 1996; Thomas et al. 1999).
Dentro del género Nothofagus los bosques de ñire presentan la mayor diversidad florística (Speziale & Ezcurra 2008). Estudios realizados por Conticello et al. (1996) en bosques de ñire y de otras especies en Patagonia, sin uso ganadero evidente, mostraron mayores valores de cobertura, riqueza y diversidad en bosques abiertos (Armesto et al. 1995; Conticello et al. 1996). En el sotobosque de ñirantales bajo el impacto del ganado doméstico y con dosel moderadamente abierto se registraron elevadas coberturas de especies herbáceas y exóticas [muchas de valor forrajero alto (Manacorda et al. 1995)], lo cual contribuye a un incremento de su riqueza específica (Bran et al. 1998; Gallo et al. 2004; Sarasola et al. 2008). En bosques altos de ñire, con condiciones óptimas de fertilidad y humedad del suelo y con presencia de ganado, el sotobosque presenta una elevada cobertura de especies herbáceas de alta calidad forrajera (Hansen et al. 2004). Debido a la relevancia de las funciones ecosistémicas del sotobosque, el objetivo de este trabajo fue analizar la relación entre la estructura del bosque y la composición específica y la diversidad del sotobosque de bosques de Nothofagus antarctica del noroeste de la Provincia de Chubut.
MATERIALES Y MÉTODOS
Sistema y área de estudio
Los bosques de N. antarctica (G. Forster) Oerst., "ñire", se desarrollan en la Patagonia Argentina entre los 36º30' y los 56º S (Veblen et al. 1996), y abarcan una superficie de 751643 ha (SAyDS 2005). Esta especie se distribuye desde las altas cumbres hasta el nivel del mar, y se introduce hacia la estepa como ribereña (Roig 1999). Es una especie decidua que puede presentar distintos morfotipos (Ramírez et al. 1985). En sitios óptimos, con suficiente humedad y bien drenados, suelos fértiles y variaciones de temperaturas anuales y diarias moderadas adquiere porte arbóreo (i.e., >15 m). Por el contrario, en suelos con variaciones hídricas amplias a lo largo del año, pobres en fertilidad y pedregosos, en zonas altoandinas y en el ecotono bosque-estepa, el ñire presenta menor altura (Ramírez et al. 1985; Veblen et al. 1996; Donoso 2006). Estos ñirantales bajos se encuentran poco representados dentro del sistema de áreas protegidas, a pesar de su alto valor para la conservación (Rusch et al. 2004) por presentar una distribución fragmentada (Donoso et al. 2006).
El área de estudio se encuentra entre los departamentos Futaleufú y Languiñeo, en el noroeste de la Provincia de Chubut, aproximadamente entre los 43º09' y 43º35' S y los 71º15' y 71º45' W. Este área se ubica fitogeográficamente dentro del dominio Subantártico, provincia Subantártica y el Distrito del Bosque Caducifolio, caracterizado por la presencia de especies de follaje caedizo como lenga y ñire entre las dominantes (Cabrera 1976). El clima de la región es templado-frío húmedo cordillerano (Burgos 1985), con una temperatura media anual de 12.2 ºC, con valores medios de 17.4 ºC y 3.8 ºC para los meses de enero y julio, respectivamente. No se registra período libre de heladas. Las precipitaciones mayores, en forma de lluvia o nieve, se producen en otoño e invierno gracias al aporte de los vientos húmedos del Pacífico, y decrecen hacia el Este en un gradiente fuerte.
Muestreo
La selección de los sitios de muestreo se basó sobre una estratificación de bosques de ñire del NO de Chubut realizada por Bran & Fertig (inédito), elaborada según la altura y la cobertura. A partir de este mapa base se identificaron nueve estructuras de bosque y luego de un reconocimiento a campo se muestrearon 10 rodales correspondientes a cada estructura. Estas distintas estructuras de bosque se denominaron Tipos Forestales e incluyeron tres clases de altura: 1) bajos: menores de 4 m, 2) medios: de 4.1 a 8 m, y 3) altos: mayores de 8.1 m de altura; y tres clases de cobertura: baja (0-40%), intermedia (41-70%) y densa (71-100%). En la estimación de la cobertura se incluyeron todos los ejemplares de ñire y de otras especies de porte arbóreo presentes en el sitio. Los tipos forestales resultantes fueron: bajos (abierto, intermedio y denso), medios (abierto, intermedio y denso), altos (abierto, intermedio y denso), se incluyeron además sectores sin cobertura arbórea (claros), rodeados de bosque de ñire. Se realizaron 10 censos de vegetación por cada tipo forestal (n=10), lo cual totalizó 100 censos distribuidos en el área de estudio. Se utilizó una unidad muestral circular de 5 m de radio, donde se relevaron las especies presentes en estrato herbáceo y arbustivo, y sus porcentajes de cobertura, mediante una estimación visual directa acordada por dos observadores.
Los nombres científicos de las especies vegetales se ratificaron con el Catálogo de Plantas Vasculares del Cono Sur (Zuloaga et al. 2008). Las plantas no identificadas hasta el nivel de especie se designaron con su nombre genérico (e.g., Trisetum sp., Carex sp., Stipa sp. y Eleocharis sp.). Así mismo, para el análisis de los datos las especies Poa ligularis, Poa pratensis, Senecio filaginoides, Senecio neaei, Bromus setifolius, Bromus stamineus, fueron designadas con el nombre genérico, debido a la gran similitud interespecífica que presentan.
Análisis de datos
Se estimó la riqueza específica (r) y los índices de diversidad de Shannon-Weaver (H) y de Simpson (D). Las especies fueron agrupadas según origen (nativas o exóticas), hábito (herbáceas, subarbustivas, arbustivas y arbóreas) y hábitat o ambiente característico (estepa, altoandino, alterado, anegado, bosque de transición y bosque húmedo) (Ezcurra & Brion 2005). Las diferencias de r, H, D, cobertura total y relativa según origen y hábito, entre tipos forestales se indagaron mediante un análisis de varianza (estadístico F, P<0.01), con una prueba "a posteriori" (Tukey, P<0.01). Se realizaron dos análisis de componentes principales, en el primero se utilizó como variable la cobertura de las especies presentes en más de 5% de los censos, y en el segundo la cobertura de todas las especies agrupadas según origen, hábito y hábitat característico, además se incluyó r, H y D. En ambos análisis de componentes principales los datos fueron previamente estandarizados y se consideró como variable clasificatoria el tipo forestal. Los análisis estadísticos se realizaron mediante el programa InfoStat versión 2009.
RESULTADOS
Especies del sotobosque
Se registraron 105 especies, distribuidas en 43 familias; las familias dominantes resultaron Asteraceae y Poaceae (ver información suplementaria en www.ecologiaaustral.com.ar). Del total de especies identificadas, las de hábito herbáceo fueron predominantes (62.4%), seguidas por las de hábito arbustivo (15.6%), subarbustivo (5.5%) y arbóreo (4.5%). En cuanto al origen, 73% pertenecieron a nativas y 27% a exóticas. El 29.9% de las especies identificadas eran características de ambientes alterados, 29.0% de bosque de transición, 17.8% de estepa y el porcentaje restante se distribuyó entre especies de bosque húmedo, altoandinas y de ambientes anegados. Las especies Schinus patagonicus, Berberis microphylla, Potentilla chiloensis, Anemone multifida, Viola maculata, Ribes cucullatum, Acaena pinnatifida, Geranium sessiliflorum, Bromus sp., Taraxacum officinale, Trifolium repens, Holcus lanatus, Poa sp. y Rumex acetosella se registraron en todos los tipos forestales.
Cobertura y Diversidad
La cobertura total del sotobosque fue menor en el bosque bajo-denso, respecto del bosque medio-abierto, de los bosques altos y de los claros (F=7.7, P<0.01) (Tabla 1). Los bosques bajo-medio, bajo-intermedio y medio-denso presentaron mayor cobertura relativa de especies nativas (F=8.4, P<0.01). Los bosques abiertos medio y alto y los claros, junto a los bosques de cobertura intermedia, medios y altos presentaron mayor cobertura relativa de especies exóticas (F=8.4, P<0.01). La cobertura relativa de especies herbáceas fue elevada en todos los tipos forestales, excepto en el bosque bajo-denso (F=7.2, P<0.01) que registró la mayor cobertura relativa de especies de hábito arbustivo (F=4.7, P<0.01). La cobertura relativa de especies subarbustivas no mostró diferencias significativas (F=2.6, P=0.01), sin embargo fue mayor en los claros y en este trabajo estuvo representada de manera especial por Acaena splendens (Tabla 1).
Tabla 1. Parámetros del sotobosque según el tipo forestal. Cobertura total, cobertura según el origen y el hábito, riqueza específica y diversidad (índice de Shannon) y dominancia (índice de Simpson). Se indican promedio y error estándar entre paréntesis. Las letras distintas indican diferencias significativas (F, P<0.01, prueba
"a posteriori" Tukey P<0.01).
Table 1. Understory parameters according to the forestal type. Total cover, and relative cover by origin and habit. Species richness, Diversity (Shannon index) and Dominance (Simpson Index). The different letters indicate significant differences (F, P<0.01, Tukey test P<0.01).
La riqueza específica no mostró diferencias significativas entre los tipos forestales (F=1.0, P>0.41), aunque presentó mayores valores en el bosque bajo-abierto y en los claros. La diversidad fue mayor en el bosque bajo-abierto respecto del bajo-denso (F=3.4, P<0.001); por el contrario, la dominancia fue superior en el bosque bajo-denso en relación a los demás, exceptuando los restantes bosques densos, con los que no presentó diferencias significativas (Tabla 1).
Composición específica
En el análisis de componentes principales, las primeras dos componentes explicaron 51% de la variabilidad (Figura 1). Este análisis permitió visualizar una asociación entre los tipos forestales de similares coberturas, indicadas como grupo I: bosques abiertos y claros, grupo II: bosques de cobertura intermedia, y grupo III: bosques densos. El grupo I incluyó a las especies Holcus lanatus, Trifolium repens, Carduus thoermeri, Hypochaeris radicata, Crepis capillaris, Poa sp., Bromus sp., Trisetum sp.,Rumex acetosella, Madia sativa, Acaena splendens, Acaena pinnatifida, Cerastium arvense, Collomia biflora, Carex sp. y Geum magellanicum. El grupo II incluyó a Geranium sessiliflorum, Galium aparine, Taraxacum officinale, Agrostis inconspicua, Senecio sp, Discaria chacaye, Elymus andinus y Ribes cucullatum, además por constituir una situación de cobertura intermedia entre bosque denso y abierto, compartió especies con ambos, como Potentilla chiloensis y Berberis microphylla con el bosque abierto y Maytenus chubutensis, Maytenus boaria, Osmorhiza chilensis, Blechnum pennamarina, Acaena ovalifolia, Veronica serpyllifolia y Viola maculata con el bosque denso. El grupo III presentó la mayor parte de las especies compartidas con el grupo II, junto a Schinus patagonicus, Quinchamalium chilense, Calceolaria lanceolata, Chusquea culeou, Viola maculata y Stellaria media. Además de la asociación entre tipos forestales de similares coberturas, los bosques bajos se reunieron formando el grupo IV; este grupo aglomeró a las especies A. inconspicua, Senecio sp, D. chacaye, R. cucullatum, E. collina, S. arenarium, Trisetum sp, A. splendens, A. pinnatifida, R. acetosella, V. maculata, A. multifida, C. culeou y M. sativa (Figura 1).

Figura 1. Análisis de componentes principales realizado con las especies presentes en más de 5% de los censos de vegetación. Se señalan los grupos correspondientes a similares coberturas: I=bosques abiertos y claros (TF: 1, 4, 7 y 10), II=bosques intermedios (TF: 2, 5 y 8), III=bosques densos (TF: 3, 6 y 9) e igual altura, IV=bosques bajos (TF: 1, 2 y 3). Las abreviaturas de las especies se encuentran en la información suplementaria (www.ecologiaaustral.com.ar).
Figure 1. Principal components analysis conducted with the species present in more than 5% of the vegetation samples. Groups of similar coverages are reported: I=open and clear forests (TF: 1, 4, 7 and 10), II=intermediate forest (TF: 2, 5 and 8), III=dense forests (TF: 3, 6 and 9), and groups of the same height: IV=low forests (TF: 1, 2 and 3). Abbreviations of species are found in suplementary information (www.ecologiaaustral.com.ar).
En el segundo análisis de componentes principales, las primeras dos componentes principales explicaron 83.1% de la variabilidad (Figura 2) y se observaron asociaciones similares a las del análisis previo (Figura 1). Los grupos a y b de la Figura 2 aglomeraron los bosques bajos intermedios y densos por un lado, y los bosques bajos abiertos y los claros; en el primer grupo se asociaron mayores coberturas relativas de especies nativas como Senecio sp., A. multifida, V. maculata y de hábito arbustivo como D. chacaye y R. cucullatum; en tanto que el segundo grupo correspondió a los bosques con mayor riqueza específica, diversidad, y mayor cobertura relativa de especies de hábito subarbustivo, características de ambientes altoandinos y de estepa, como A. splendens, S. arenarium, E. collina. Además estos tipos forestales también presentaron mayores frecuencias de especies de ambientes de estepa y altoandinos (ver información suplementaria en www.ecologiaaustral.com.ar). El 58% de las especies de estepa presentó una frecuencia mayor en el bosque bajo-abierto y/o en los claros y 26 % en el bosque bajo-intermedio y/o bajo-denso, en tanto que 33% de las especies altoandinas registraron una frecuencia mayor en los bosques bajo-abierto y/o en los claros (ver información suplementaria en www.ecologiaaustral.com.ar).

Figura 2. Análisis de componentes principales realizado con las coberturas de todas las especies según origen (nativas y exóticas), hábito (herbáceas, subarbustivas y arbustivas) y ambiente característico [anegado, alterado, altoandino, estepa, bosques de transición (BosqueT) y húmedo (BosquesH)], los índices de Shannon (H), de Simpson (D) y la riqueza específica (r). Se señalan los grupos correspondientes a similares coberturas: A=bosque bajo denso e intermedio (TF 2 y 3), B=bosque bajo-abierto y claros (TF: 1 y 10), C=bosques medios y altos de dosel abierto o intermedio (TF: 4, 5, 7 y 8), y D=bosques medios y altos densos (TF: 6 y 9).
Figure 2. Principal components analysis conducted with the coverages of all species by origin (native and exotic), habit (herbaceous, sub-shrub and shrub), and habitat (overflooded, disturbed forest, high Andean, steppe, transitional forest and moist forest). Groups of similar coverages are reported: A=dense and intermediate low forest (TF 2 and 3), B=open and clear low forests (TF: 1 and 10), C=high or medium forest with clear or intermediate canopy coverage (TF 4, 5, 7 and 8), and D=high or medium dense forest (TF 6 and 9).
El grupo c de la Figura 2 agrupa los bosques medio-abierto, medio-intermedio, alto-abierto y alto-intermedio, y reúne las mayores coberturas relativas de especies exóticas y de hábito herbáceo, características de ambientes alterados y anegados, como C. capillaris, T. officinale, Carex sp., G. magellanicum, T. repens, Holcus lanatus y P. chiloensis. Estos tipos forestales, además, presentaron las mayores frecuencias de las especies correspondientes a los ambientes anegados y alterados. El 45 % de las especies de ambientes alterados presentó una frecuencia mayor en los tipos abiertos de altura media y/o alta, y 10% en los de cobertura intermedia, de altura media y alta, mientras que 67% de las especies de ambientes anegados presentó mayor frecuencia en los tipos abiertos de altura media y/o alta, y 11% en los de cobertura intermedia, de altura media y alta TF 5 y/o 8 (ver información suplementaria en www.ecologiaaustral.com.ar). Nuevamente, los bosques medios y altos de cobertura intermedia se presentaron colindantes tanto con los bosques abiertos como con los densos, de la misma altura.
En el grupo d de la Figura 2 se reúnen los bosques densos de mayor altura, al cual se asocian el índice de dominancia de Simpson y las especies de bosques húmedo y de bosque de transición como S. patagonicus, V. maculata, M. chubutensis, M. boaria y O. chilensis. Estos tipos forestales también presentaron la mayor frecuencia de especies características de bosque húmedo y de transición. El 43% de las especies de bosque húmedo presentó mayor frecuencia en el bosque medio-denso y/o alto-denso, y el 29 % en los bosques medio y alto de cobertura intermedia, mientras que 35% de las especies de bosques de transición presentó mayor frecuencia en los medios y altos de cobertura densa y 19% en los medios y altos de cobertura intermedia (ver información suplementaria en www.ecologiaaustral.com.ar).
DISCUSIÓN
La mayoría de las especies registradas en el sotobosque de ñire fueron nativas y características de ambientes alterados y de bosques de transición (Ezcurra & Brion 2005). La cobertura fue elevada siendo variable la dominancia de especies nativas y exóticas en los distintos bosques estudiados. La cobertura total del sotobosque fue superior en los bosques altos; esto podría asociarse a mejores condiciones de sitio, humedad y fertilidad del suelo, que caracteriza a las áreas donde el ñire adquiere una estructura arbórea (Ramírez et al. 1985; Donoso et al. 2006). De igual forma, los claros y el bosque medio-abierto registraron coberturas totales elevadas, que podrían asociarse a mayor radiación solar incidente en el sotobosque que permite un mayor desarrollo de la vegetación (Klinka et al. 1996; Thomas et al. 1999). En este sentido, el bosque bajo-denso registró menor cobertura total, lo que estaría relacionado a condiciones deficientes de sitio (Ramírez et al. 1985; Hansen et al. 2004; Donoso et al. 2006; Peri 2009) así como a la escasa luz que llega a los estratos inferiores (Klinka et al. 1996; Holmgren et al. 1997; Thomas et al. 1999). Esta menor cobertura de herbáceas podría asociarse también al pastoreo y a los incendios frecuentes que forman comunidades secundarias con incremento de especies arbustivas y subarbustivas (Seibert 1979).
La cobertura de especies exóticas predominó en los claros y en los bosques medio y alto de cobertura abierta o intermedia, como mencionaran Bran et al. (1998) y Sarasola et al. (2008); esta creciente colonización de especies ruderales, podría deberse a una respuesta observada en otros bosques patagónicos bajo intenso impacto de grandes ungulados (Relva & Veblen 1998; Vázquez 2002). Sin embargo, la cobertura de especies exóticas también aumentaría en sitios con bajo grado de uso, indicando la existencia de un grupo de especies exóticas como T. officinale, T. repens, H. radicata, R. acetosella, V. serpyllifolia y H. lanatus que penetran en la comunidad apenas se produce algún grado de disturbio (Gallopin et al. 2005). Por el contrario, la cobertura de especies nativas fue superior en los bosques bajos, intermedios y densos, y en el bosque medio-denso. Las especies nativas de los bosques bajos son propias de ambiente altoandino y de estepa, mientras que en el medio-denso predominan especies nativas de bosque de transición y de bosque húmedo, indicadoras de menor alteración antrópica (San Martín et al. 1991; Gallopin et al. 2005).
La apertura del dosel se asocia a una mayor riqueza y diversidad del sotobosque (Conticello et al. 1996), como se registró en el bosque bajo abierto. Esta mayor diversidad podría deberse a una mayor alteración de estos ambientes (Rusch et al. 2005). El efecto combinado de los disturbios antrópicos y las condiciones rigurosas de sitio, determinan la confluencia de especies de estepa, de bosque de transición, de ambiente altoandino y alterado, sin dominancia ni exclusión de especies, permaneciendo todas en los ñirantales bajos (Connell 1978). Por el contrario donde la cubierta del dosel es densa, como en el bosque bajo-denso, la riqueza y diversidad disminuyen, y aumenta la dominancia de algunas especies tolerantes a la sombra (Armesto et al. 1995; Conticello et al. 1996).
Se presentaron algunos grupos de especies asociados a TF de similares coberturas o alturas. En el bosque sombrío y húmedo fueron frecuentes las especies propias de bosque de transición y bosque húmedo, como O. chilensis, B. penna-marina, A. ovalifolia y M. chubutensis, características de sectores poco alterados (San Martín et al. 1991; Gallopin et al. 2005). En los ñirantales arborescentes la apertura del dosel provoca un incremento de la cobertura de especies herbáceas y exóticas indicadoras de sitios alterados, entre las cuales se incluyen las especies de buena calidad forrajera como H. lanatus, T. repens, T. pratense, Poa sp. y Bromus sp, entre otras (Manacorda et al. 1995; Manacorda et al. 2001; Hansen et al. 2004; Olivares et al. 2008; Sarasola et al. 2008). Los bosques bajos presentan mayores coberturas relativas de especies nativas principalmente de hábito arbustivo, características de bosque de transición y estepa. El bosque bajo abierto se asocia principalmente a los claros y comparten especies como A. splendens, Poa sp, R. acetosella, C. thoermeri y H. radicata, características de estepa y de sitios degradados (Montaña 1982; Bran et al. 1998).
CONCLUSIONES
El sotobosque de ñire presenta una cobertura elevada de especies, con una gran variación en la composición específica entre los tipos de bosques analizados. Sin embargo, las especies Schinus patagonicus, Berberis microphylla, Potentilla chiloensis, Anemone multifida, Viola maculata, Ribes cucullatum, Acaena pinnatifida, Geranium sessiliflorum, Bromus sp., Taraxacum officinale, Trifolium repens, Holcus lanatus, Poa sp. y Rumex acetosella, se registraron en todas las estructuras analizadas.
En los ñirantales arborescentes y densos se registra un sotobosque prístino con especies nativas típicas de bosque húmedo. En estos bosques la apertura del dosel induce un incremento de la cobertura de especies herbáceas y exóticas. En los bosques bajos se registra menor cobertura del sotobosque, con una mayor importancia relativa de especies nativas de escaso valor forrajero. Las condiciones ambientales rigurosas y la mayor apertura del dosel en los bosques bajos, conducen a una mayor diversidad de especies, si bien es importante mencionar que estos sitios diversos serían equivalentes a sectores sin cubierta arbórea que presentan una frecuencia elevada de especies indicadoras de degradación.
La cobertura del bosque influye en el sotobosque, y esto repercute sobre la composición específica y genera cambios ligados a las condiciones de sitio. Teniendo en cuenta que estos sistemas boscosos están expuestos a intensos usos antrópicos en la región, resultaría imprescindible generar herramientas que permitan un manejo ecológicamente viable de los ñirantales.
INFORMACIÓN SUPLEMENTARIA
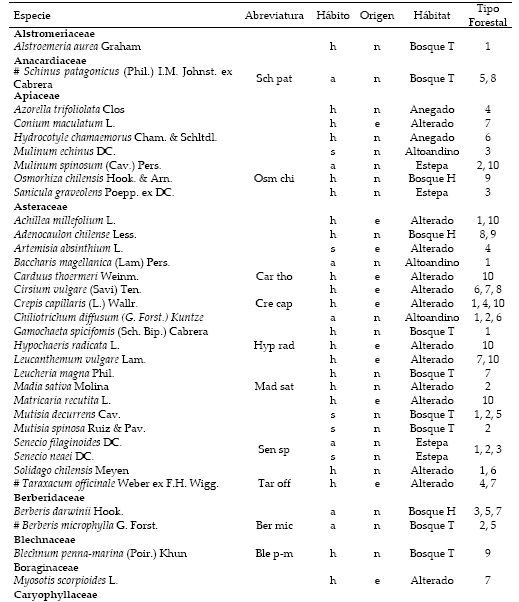
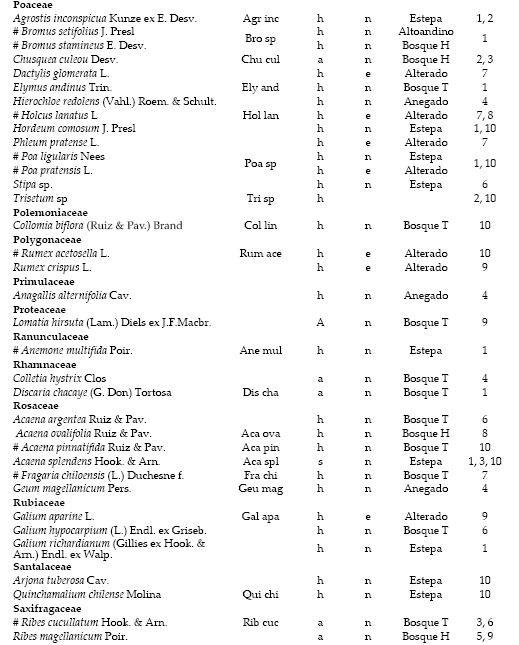
Lista de especies ordenadas por familia, incluyendo el origen (n=nativas; e=exóticas), hábito (h=herbáceas; s=subarbustivas; a=arbustivas; A=arbóreas) y hábitat [Estepa, altoandino, anegado, bosque húmedo y bosque de transición (Bosque H y T)]. Se señalan las especies registradas en todos los tipos forestales (#), la abreviatura utilizada para los análisis de componentes principales de las especies frecuentes en más de 5% de los censos y el tipo forestal donde se registró la frecuencia mayor de cada especie.
List of species ordered by family, including the origin (n=native; e=exotic), habit (h=herbaceous; s=subshrub, a=shrub; A=trees) and habitat (steppe, high Andean, overflooded, moist forest and transitional forest [H and T forest)]. The species recorded in all the forest type (#), the abbreviation used for principal components analysis common species by more than 5% of the census, and the forest type where there was greater frequency of each species are identified.




BIBLIOGRAFÍA
1. ARMESTO, JJ; C VILLAGRÁN & MK ARROYO (eds). 1995. Ecología de los Bosques Nativos de Chile. Editorial Universitaria, Santiago, Chile. Pp. 470. [ Links ]
2. BAVA, J; A HAAG; GN AGUADO; H CLAVERIE; P LÓPEZ BERNAL; ET AL. 2008. Distribución, estado y potencialidad para la producción maderera de los bosques del Chubut. En: Libro de actas II Reunión sobre Nothofagus en la Patagonia. Esquel, Chubut. [ Links ]
3. BRAN, D; S RUSAK; J AYESA & H MORAGA. 1998. Los matorrales de Nothofagus antarctica (ñire) en el contacto Bosque-Estepa. Estructura y regeneración después de una tala. Comunicación técnica Nº 47. INTA. EEA. Bariloche. [ Links ]
4. BURGOS, J. 1985. Clima del extremo sur de Sudamérica. En: Boelcke, O; DM Moore y FA Roig (eds.), Transecta Botánica de la Patagonia Austral. Pp. 10-48. CONICET (Argentina), Royal Society (Gran Bretaña) e Instituto de la Patagonia (Chile). [ Links ]
5. CABRERA, A. 1976. Regiones Fitogeográficas Argentinas. Enciclopedia Argentina de Agricultura y Jardinería. Tomo II.2ª Edición. Ed. ACME S.A.C.I. Bs. As. [ Links ]
6. CONTICELLO, L; R GANDULLO; A BUSTAMANTE & C TARTAGLIA. 1996. Fitosociología de los bosques caducifolios del norte del Departamento Lácar y sur de Huiliches de la provincia de Neuquén (Argentina). Bosque, 17:27-43. [ Links ]
7. CONNELL, ML. 1978. Diversity in tropical rain forests and coral reefs. Science, 199:1302-1310. [ Links ]
8. EZCURRA, C & C BRION. 2005. Plantas del Nahuel Huapi. Catálogo de La Flora Vascular del Parque Nacional Nahuel Huapi. Argentina. Ed. Bavaria. San Carlos de Bariloche. Argentina. Pp. 70. [ Links ]
9. DGBYP (DIRECCIÓN GENERAL DE BOSQUES Y PARQUES). 2004. Reglamento único de aprovechamiento forestal de los bosques de la provincia del Chubut. R764/04. [ Links ]
10. DONOSO, C; L STEINKE & A PRÉMOLI. 2006. Nothofagus antarctica (G. Forster) Oerst. Ñirre, Ñire, Ñiré, Anís (Tierra del Fuego) Ñirre: de Ngërü (mapudungun): zorro. En: Donoso, C (ed.), Las especies arbóreas de los bosques templados de Chile y Argentina. Autoecología. Ed. Marisa Cuneo. Santiago, Chile. [ Links ]
11. ELLUM, DS. 2009. Floristic Diversity in Managed Forests: Demography and Physiology of Understory Plants Following Disturbance in Southern New England Forests. J. Sust. For., 28: 132-151. [ Links ]
12. GALLO, M; M LENCINAS & P PERI. 2004. Modificación de la biodiversidad por el manejo forestal: plantas, aves e insectos. Informe del Proyecto de Investigación Aplicada a los Recursos Forestales Nativos. Región Bosques Andino-Patagónicos. Pp. 25. [ Links ]
13. GALLOPIN, GC; CE MARTIN & MA MERMOZ. 2005. Impacto de la ganadería en la cuenca del río Manso Superior. Parte I: Bosque de ñire con laura. Anales de Parques Nacionales, Tomo XVII. [ Links ]
14. GUITART, E. 2004. Diagnóstico productivo orientado a establecer la capacidad del sector ganadero del NO del Chubut para involucrarse y sostener un proyecto de diferenciación por calidad del producto carne vacuna. Informe Interno. INTA. Pp. 26. [ Links ]
15. HANSEN, N; M FERTIG & L TEJERA. 2004. Sistemas silvopastoriles en Chubut. Informe del Proyecto de Investigación Aplicada a los Recursos Forestales Nativos. Región Bosques Andino-Patagónicos. Pp. 8. [ Links ]
16. HANSEN, N; M FERTIG; M ESCALONA; L TEJERA & W OPAZO. 2008. Ramoneo en regeneración de ñire y disponibilidad forrajera. En: Libro de actas II Reunión sobre Nothofagus en la Patagonia. Esquel, Chubut. [ Links ]
17. KLINKA, K; H CHEN; Q WANG & L MONTIGNY. 1996. Forest Canopies and Their Influence on Understory Vegetation in Early-Seral Stands on West Vancouverls land. North. Sci., 70:193-200. [ Links ]
18. MANACORDA, M; R SOMLO; A PELLIZA SBRILLER & P WILLEMS. 1995. Dietas de ovinos y bovinos en la región de los bosques de Ñire (Nothofagus antarctica) de Río Negro y Neuquén. RIA, 26: 137-146. [ Links ]
19. MANACORDA, M & G BONVISUTTO. 2001. Uso silvopastoral de los bosques de ñire (Nothofagus antarctica) en Río Negro, Patagonia Argentina. Rev. For. Centroa., 35:41-44. [ Links ]
20. MONTAÑA, C. 1982. Las comunidades de ñire (Nothofagus antarctica (Forst.) Oerst.) de la cuenca del río Manso Superior (Río Negro, Argentina). Tesis Doctoral. Univ. Nac. de Córdoba. [ Links ]
21. OLIVARES, A; H SCHMIDT; J PAVEZ & S DURAN. 2008. Efecto de la Cobertura de Ñirre (Nothofagus antarctica) sobre la Producción Herbácea en la Provincia de Última Esperanza, Región De Magallanes. Ciencia e Investigación Forestal, 14: 55-63. [ Links ]
22. PERI, P. 2005. Sistemas silvopastoriles en ñirantales. IDIA XXI Revista de información sobre investigación y desarrollo agropecuario INTA, 255-261. [ Links ]
23. PERI, P. 2009. Sistemas silvopastoriles en Patagonia: revisión del conocimiento actual. 1er. Congreso Nacional de Sistemas Silvopastoriles. Misiones. Argentina. [ Links ]
24. RAMÍREZ, C; M CORREA; H FIGUEROA & J SAN MARTÍN. 1985. Variación del hábito y hábitat de Nothofagus antarctica en el centro de Chile. Bosque, 2:55-73. [ Links ]
25. RELVA, M & T VEBLEN. 1998. Impacts of introduced large herbivores on Austrocedrus chilensis forest in northern Patagonia, Argentina. For. Eco. Manage., 108:27-40. [ Links ]
26. REQUE, J; M SARASOLA; J GYENGE & ME FERNÁNDEZ. 2007. Caracterización Silvícola de Ñirantales del Norte de la Patagonia para la Gestión Forestal Sostenible. Bosque, 6:55-73. [ Links ]
27. ROIG, FA. 1999. La vegetación de la Patagonia. En: Correa, M (dir.), Flora Patagónica. Colección Científica del INTA, 8(1):48-166. [ Links ]
28. RUSCH, V; R ROVETA; C PERALTA; B MÁRQUES; A VILA; ET AL. 2004. Criterios indicadores de manejo sustentable. Informe del Proyecto de Investigación Aplicada a los Recursos Forestales Nativos. Región Bosques Andino-Patagónicos. Pp. 63. [ Links ]
29. RUSCH, V; M SARASOLA & T SCHLICHTER. 2005. Indicadores de Biodiversidad en Bosques Nothofagus. IDIA XXI Forestales, INTA, 5(8):8-14. [ Links ]
30. SAYDS (SECRETARIA DE AMBIENTE Y DESARROLLO SUSTENTABLE). 2005. Primer inventario de bosques nativos. Proyecto Bosques Nativos y Aéreas Protegidas. Pp. 104. [ Links ]
31. SAN MARTÍN, J; A TRONCOSO; A MESA; T BRAVO & C RAMÍREZ. 1991. Estudio fitosociológico del bosque caducifolio magellánico en el límite norte de su área de distribución. Bosque, 12(2):29-41. [ Links ]
32. SARASOLA, M; D LÓPEZ; J GAITÁN & G SIFFREDI. 2008. Productividad de sistemas silvopastoriles en bosques de ñire en la cuenca del río Foyel. Libro de actas II Reunión sobre Nothofagus en la Patagonia. Esquel, Chubut. [ Links ]
33. SEIBERT, P. 1982. Carta de vegetación de la región de El Bolsón, Rio Negro y su aplicación a la planificación del uso de la tierra. Documenta Phytosociológica, 2:1-120. [ Links ]
34. SPEZIALE, K & C EZCURRA. 2008. ¿Es importante conservar el ñire? Libro de actas II Reunión sobre Nothofagus en la Patagonia. Esquel, Chubut. [ Links ]
35. THOMAS, SC; CB HALPERN; DA FALK; DA LIGUORI & KA AUSTIN. 1999. Plant Diversity in Managed Forests: Understory Responses to Thinning and Fertilization. Ecol. Appl., 9(3):864-879. [ Links ]
36. VALLADARES, F; I ARANDA & D SÁNCHEZ-GÓMEZ. 2004. La luz como factor ecológico y evolutivo para las plantas y su interacción con el agua, 335-369. En: Valladares, F, Ecología del bosque mediterráneo en un mundo cambiante. Ministerio de Medio Ambiente, EGRAF, S.A., Madrid-España. [ Links ]
37. VÁZQUEZ, DP. 2002. Multiple effects of introduced mammalian herbivores in a temperate forest. Biol. Inv., 4:175-191. [ Links ]
38. VEBLEN, T & D LORENZ. 1988. Recent vegetation changes along the forest/steppe ecotone Northern Patagonia, Annals Assoc. Amer. Geogr., 78(1):93-111. [ Links ]
39. VEBLEN, T; R HILL & J READ. 1996. The Ecology and Biogeography of Nothofagus forest. Yale University press, New Haven and London. [ Links ]
40. ZULOAGA, F; O MORRONE & M BELGRANO (EDS.). 2008. Catálogo de las Plantas Vasculares del Cono Sur. Monographs in Systematic Botany, Missouri Botanical Garden, 107:1-3348. [ Links ]

