










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.20 no.3 Córdoba Sept./Dec. 2010
TRABAJOS ORIGINALES
Influencia de la vegetación en la variación espacial de la abundancia de microorganismos en el desierto del Monte, San Juan, Argentina
Ángela D Vega Ávila1,*, María E Toro1, Mario Baigori2, Luciana Fernández1 & Fabio Vázquez1
1 Instituto de Biotecnología. Facultad de Ingeniería. Universidad Nacional de San Juan. Av. Libertador San Martín 1109 (Oeste), San Juan.
2 Planta Piloto de Procesos Industriales Microbiológicos Avenida Belgrano y Pasaje Caseros. Tucumán
* Instituto de Biotecnología. Facultad de Ingeniería. Universidad Nacional de San Juan. Av. Libertador San Martín 1109 (Oeste), San Juan.
Email: dvega@unsj.edu.ar.
Recibido: 29 de marzo de 2010;
Fin de arbitraje: 25 de mayo de 2010;
Revisión recibida: 3 de agosto de 2010;
Aceptado: 30 de agosto de 2010
RESUMEN. En los sistemas desérticos la vegetación frecuentemente se distribuye en parches. Asociados a estos parches, existen microorganismos que son importantes para el crecimiento de las plantas, ya que favorecen la absorción de nutrientes, fijan nitrógeno e intervienen en la descomposición de la materia orgánica y liberan enzimas fundamentales en el ciclo de los nutrientes. Los objetivos de este trabajo fueron: evaluar la abundancia de los principales grupos microbianos (bacterias, levaduras y hongos filamentosos); detectar y cuantificar bacterias con actividades enzimáticas degradadoras, asociadas a suelos y canopias de los parches de Bulnesia retama, Larrea divaricata e interparches con baja cobertura vegetal a fin de establecer posibles relaciones entre la distribución de la vegetación y los microorganismos responsables de la descomposición. No detectamos diferencias significativas en la abundancia de bacterias y levaduras entre los distintos tipos de micrositios, si bien observamos una mayor abundancia de hongos filamentosos en el suelo de los parches de L. divaricata con respecto a los de suelo desnudo. La proporción de colonias de bacterias aisladas con actividades enzimáticas (xilanolítica y amilolítica) fue mayor en los parches de L. divaricata y B. retama respecto a interparches. Las bacterias aisladas de hojas de L. divaricata presentaron actividad xilanolítica, amilolítica y celulolítica. Se encontró que en los parches existe una mayor abundancia de bacterias que en las hojas de las canopias de los vegetales, como así también mayor porcentaje de colonias de bacterias que presentaron alguna de las actividades enzimáticas involucradas en la descomposición de materia orgánica. Los resultados sugieren que podría existir una relación entre la abundancia de algunos grupos funcionales de bacterias y la distribución de la vegetación.
Palabras clave: Grupos funcionales; Actividades enzimáticas; Parches de vegetación; Interparches.
ABSTRACT. Influence of vegetation in the spatial variation of microbial abundance in Monte desert, San Juan, Argentina: Due to the lack of water, vegetation in desertic ecosystems is distributed in patches. Microorganisms associated to patches are important in plant growth because favor nutrients uptake, nitrogen fixation and organic matter decomposition; also release fundamental enzymes in nutrients cycle. The objectives of this work were to evaluate the abundance of microbial groups (bacteria, yeasts and filamentous fungi) and to detect and quantify bacteria with enzymatic activities in soils and leaves of Bulnesia retama, Larrea divaricata, and interspaces with bare soil areas; to establish relations between plant distribution and the functional groups of microorganisms involved in litter plant decomposition. No significant differences were detected in the abundance of bacteria and yeasts among all different microsites, although filamentous fungi abundance was higher in soil of patches associated to L. divaricata respect to bare soil areas. The percentage of isolated bacteria colonies that showed enzymatic activities (xylanolitic and amylolitic) was higher in patches of L. divaricata and B. retama than in interspaces. Isolated bacteria from leaves of L. divaricata showed xylanolitic, amylolitic and cellulolitic activities. It was found that abundance of bacteria in patches was higher than in leaves of canopies of the studied plants; it was also found higher percentage of bacteria colonies that showed at least one of the enzymatic activities involved in organic matter decomposition. The results suggest that there could be a relationship between abundance of some bacteria functional groups and the vegetation distribution.
Keywords: Functional groups; Enzymatic activities; Vegetation patches; Interspaces.
INTRODUCCIÓN
En los sistemas desérticos, donde la escasez de agua es la limitante principal para la actividad biológica, la vegetación suele distribuirse en parches con una alta cobertura vegetal, dispersos en una matriz de suelo de baja cobertura (interparches). Estos parches a menudo están dominados por vegetación arbustiva y presentan una acumulación fuerte de nutrientes bajo la canopia de los arbustos (i.e., islas de fertilidad), mientras que el suelo en los interparches es relativamente infértil (Schlesinger & Pilmanis 1998; Aguiar & Sala 1999; Hmerlynck et al. 2002; Abril et al. 2009; González-Polo & Austin 2009). Los microorganismos asociados a las islas de fertilidad son importantes para el crecimiento de las plantas ya que favorecen la absorción de nutrientes por las plantas, producen hormonas que promueven su crecimiento, fijan nitrógeno, suprimen patógenos e intervienen en la descomposición de la materia orgánica (Farnsworth et al. 1977; Nakas & Klein 1980; Davison 1988; Denarie et. al. 1992; Aguilera et al. 1999).
En general, las condiciones en los ambientes áridos y semiáridos son desfavorables para el crecimiento microbiano masivo. Se ha sugerido que las comunidades microbianas de este tipo de ecosistemas están dominadas por microorganismos inactivos, incluyendo formas esporulantes (Mamilov & Dilly 2002). La variación de la biomasa microbiana es un indicador medible de la calidad y del potencial de uso de un suelo debido a que es una de las pocas fracciones de materia orgánica, biológicamente significativa, sensible a las consecuencias del manejo humano y a la polución (Schloter et al. 2003). Los grupos de microorganismos más importantes son las bacterias, las levaduras y los hongos filamentosos (Mamilov & Dilly 2002; Schloter et al. 2003). En general, su distribución espacial está relacionada con variaciones en la cantidad y calidad de la materia orgánica, con la disponibilidad de nutrientes y con la humedad del suelo (Vishnevestky & Steinbergert 1997; Aguilera et al. 1999).
La descomposición de la materia orgánica del suelo, además de estar regulada por el clima y la biota es muy afectada por la calidad del material a degradar. Los detritos vegetales están constituidos por celulosa y hemicelulosas, que constituyen los elementos orgánicos más abundantes además del almidón, que es la mayor reserva de carbohidratos de las plantas superiores (Frioni 1999). En ecosistemas áridos patagónicos se ha observado que las hojas de arbustos y hierbas perennes poseen mayor contenido de compuestos carbonados que son descompuestos lentamente (Carrera et al. 2009).
Parte de esta descomposición es realizada a través de enzimas liberadas por los microorganismos. Estos microorganismos cumplen un papel fundamental en el ciclo de los nutrientes (Weaver et al. 1994; Alef et al.1995; Dick et al. 1996; Aon et al. 2001; Badiane et al. 2001). Su actividad también es regulada por factores abióticos (i.e., pH, temperatura, contenido de agua y contenido de oxígeno) y bióticos (i.e., composición del residuo vegetal, relación C/N, concentración de compuestos secundarios, etc.) (Deng &Tabatabai 1995; Carrera et al. 2009). Estudios previos muestran que en suelos de ambientes áridos se encontraron bacterias que secretan enzimas hidrolasas (Dekker & Richards 1976; Torres & Abril 2005). Sin embargo, no se han hallado trabajos que evalúen la abundancia de microorganismos y la actividad enzimática de las bacterias asociadas a las hojas de canopias o a suelos bajo los arbustos de la familia Zigofiláceas en la provincia fitogeográfica del Monte argentino. Un antecedente sobre la diversidad de levaduras en canopias y suelos asociados con Bulnesia retama y Larrea divaricata muestra que la mayor parte de los géneros, y aun especies de levaduras aisladas de suelo, se encontraron representadas en la parte aérea de los arbustos. Esto sugiere una relación entre la microflora de la parte aérea de la planta, los suelos asociados e incluso con el suelo de los interparches (Toro et al. 2005). Los objetivos de este trabajo fueron evaluar la abundancia de los principales grupos microbianos (bacterias, levaduras y hongos filamentosos) y detectar y cuantificar la presencia de bacterias con actividades enzimáticas degradadoras en parches de suelo dominados por Bulnesia retama y Larrea divaricata, en la canopia de individuos de estas especies y en los interparches. A través de este estudio intentamos establecer posibles relaciones entre la distribución de la vegetación y los microorganismos responsables de la descomposición de los residuos vegetales.
MATERIALES Y MÉTODOS
Sitio y especies estudiadas
El sitio de estudio se ubica en los Médanos Grandes localizados sobre la vertiente occidental de la sierra de Pie de Palo, alejada del piedemonte, en el departamento de Caucete, a 45 km de la Ciudad de San Juan, Argentina. El clima es seco y templado, con inviernos frescos. La temperatura media de enero y julio para 10 años de registros es de 25.1 y 7.7 ºC, respectivamente (Cabrera & Willink 1980). La precipitación media anual es de 103 mm, con un régimen estacional muy diferenciado. Las lluvias, de tipo torrencial, se producen durante el verano entre los meses de enero a marzo (De Fina 1992; Bisigato et al. 2009). Los Médanos Grandes son una extensa llanura de origen eólico e hídrico cuaternario, con suelos arenosos, profundos y muy permeables, interrumpidos por grandes dunas longitudinales. El agente erosivo predominante en el lugar es el viento, lo cual da origen al patrón de vegetación leopardo (Strahler 1992; Abraham et al. 2009). El área de estudio forma parte del Desierto del Monte Central (Rundel et al. 2007; Bisigato 2009; Carrera et al. 2009). La vegetación está dominada por especies de la familia Zigofilácea como ocurre en otros desiertos alrededor del mundo (Aguiar & Sala 1999).
Una de las especies dominantes es Bulnesia retama, conocida de manera local como "retamo". San Juan es el área ecológica más importante para la especie ya que ocupa 80% del territorio de la provincia y llega hasta 2800 m.s.n.m. Esta especie es un arbusto perenne, áfilo de entre 3 y 5 m de altura, con flores hermafroditas de color amarillo y fruto esquizocárpico (Palacios & Hunziker 1984). Larrea divaricata, conocida como "jarilla", es otra de las especies dominantes en la región. Es un arbusto perenne de entre 1 y 3 m de altura; en sus hojas y tallos produce grandes cantidades de resina fenólica cuyo componente mayor (80%) es el ácido nordihidroguayarético (Obermeyer et al. 1995; Stege et al. 2005). Sus flores son amarillas y florece desde octubre hasta fines de noviembre (Vilela et al. 2009). Tiene gran importancia biogeográfica porque cubre regiones áridas y semiáridas extensas del sur de América (Cortes & Hunziker 1997).
Entre las hierbas anuales presentes en el área de estudio se pueden citar Sclerophylax arnottii Miers, Ibicella parodii Abbiatti, Portulaca grandiflora Hook, Portulaca echinosperma Hauman, Calicera spinulosa Gillies ex Miers, Morrenia odorata Lindl, Chenopodium papulosum Moq. y pastos como Aristida mendocina Phil., Pappophorum philippianum Parodi, Pappophorum caespistosum R. Fries y la exótica Schismus barbatus (L.) Thell. S. barbatus se encuentra distribuida de manera uniforme por debajo y entre los parches, a través del médano e intermédano principalmente después de las lluvias (Ribas et al. 2009).
Diseño del muestreo y análisis de suelo, plantas y microorganismos
En agosto de 2005 se tomaron muestras de suelo en seis parches de Bulnesia retama a 1 m del tallo principal, y en seis de parches de Larrea divaricata a una distancia de entre 0.3 y 0.4 m del tallo principal e interparches. De cada micrositio se extrajeron 3 submuestras de los primeros 5 cm de profundidad (casi 200 cm3) con un testigo esterilizable "in situ". En las canopias de ambos arbustos se tomaron seis muestras de hojas al azar con elementos esterilizables y se colocaron en bolsas estériles para su posterior análisis en laboratorio.
La determinación de abundancia de los distintos grupos microbianos se realizó a partir de 1 g de muestra de cada tipo de parche y arbusto (las muestras de suelo fueron tamizadas previamente y las de hojas fueron seccionadas en trozos pequeños). Las muestras fueron colocadas en frascos de 250 ml de capacidad con 100 ml de solución fisiológica estéril (8.5 g/L de NaCl) y se agitaron a 250 rpm durante 1 h a 25 °C (Kurtzman 2000). Del sobrenadante de cada muestra se realizaron por triplicado diluciones apropiadas para un conteo adecuado de las UFC (Unidades Formadoras de Colonias) y se sembró 50 µL en placas de Petri con medios de cultivo para el desarrollo de bacterias, levaduras, y hongos filamentosos, respectivamente. Las placas se incubaron durante 72 h a una temperatura de 28 °C. Se discriminó cada grupo microbiano (hongos filamentosos, levaduras, bacterias) teniendo en cuenta la morfología de colonia. Dicho agrupamiento se confirmó mediante observación con microscopio de campo claro. Las colonias con iguales características, correspondientes a los diferentes tipos de microorganismos, se cuantificaron en unidades formadoras de colonias por gramo de suelo o gramo de hoja.
El medio de cultivo utilizado para el desarrollo de bacterias fue caldo nutritivo: extracto de carne, de 3.0 a 5.0 g/L; NaCl, 5 g/L; Na2HPO4, 1 g/L; agar, 20 g/L, pH=7. Para levaduras se utilizó YEPD: extracto de levadura, 10 g/L; peptona, 20 g/L y glucosa, 20 g/L; agar 20 g/L. PH=4.5. Para hongos filamentosos se usó PDA, pH (Agar Papa Glucosada): glucosa, 5 g/L; agar, 20 g/L y extracto de papa, 100 ml/L; pH=5. Para la detección de la actividad de amilasas se utilizó como sustrato 10 g/L de almidón soluble, 10 g/L de extracto de levadura, 0.6 g/L de MgCl2, 0.1 g/L de CaCl2, con agua destilada. Se le agregó agar 20 g/L (Vázquez et al. 2004). Para la detección de actividad de xilanasas se utilizó como sustrato 1.5 g/L de xilano (Birchwood xylan, Sigma Chemical Co.) con agua destilada. Se le agregó agar 20 g/L, 6.7 g/L de Yeast Nitrogen Base (YNB) sin aminoácidos, 2 g/L de D-L-asparagina y 5.0 g/L de KH2PO4 (Lee et al. 1986). Para la detección de actividad de celulasas se utilizó como sustrato 4 g/L de carboximetil celulosa (CMC), 6.7 g/L de Yeast Nitrogen Base (YNB), con agua destilada. Se le agregó agar 20 g/L (Vázquez et al. 2004).
Se discriminaron y aislaron grupos bacterianos de acuerdo con la morfología de colonia (color, forma, tipo de superficie, brillo, bordes) y se detectó las actividades enzimáticas (xilanolíticas, amilolíticas y celulolíticas), empleando los medios descriptos, mediante la formación de halos de degradación. Este procedimiento se realizó por triplicado, se usó un control positivo y se incubaron las bacterias por 48 h a 25 °C. Para el caso de la detección de actividades celulolítica y xilanolítica, transcurrido el periodo de incubación, las placas con desarrollo de colonias fueron lavadas con agua destilada y luego fueron teñidas con solución Rojo Congo 0.2%. Se agitó durante 10 minutos a temperatura ambiente, se descartó el colorante excedente y se lavaron las placas 2 veces durante 5 minutos con NaCl 0.5 M. La presencia de actividad enzimática se confirmó mediante la formación de halos de degradación alrededor de las colonias sobre los medios teñidos con la solución de Rojo Congo (Strauss et al. 2001). Para la detección de actividad amilolítica, cumplida la etapa de incubación las placas con desarrollo de colonias se revelaron mediante la incorporación de Lugol y se dejó actuar entre 5 y 10 minutos.
Se determinó el pH del suelo en extracto acuoso (1:5 p/v) y el contenido de humedad gravimétrica a partir de muestras de 3 g de suelo que fueron secadas a una temperatura de entre 105 y 110 °C hasta peso constante.
Análisis estadísticos
La abundancia de bacterias se analizó por medio de ANOVA y las medias se compararon por medio de pruebas de Tukey. El resto de las variables no presentó una distribución normal por lo cual fue analizado con pruebas no-paramétricas de Kruskal-Wallis (InfoStat 2.0). En todos los casos se informa la media y el error estándar (Di Rienzo et al. 2002).
RESULTADOS
No se detectaron diferencias significativas en la abundancia de bacterias y levaduras entre los distintos micrositios, mientras que la abundancia de hongos filamentosos fue mayor (P=0.03) en el suelo de los parches de L. divaricata que en los interparches, y nula en el parche de B. retama. La abundancia de bacterias fue significativamente mayor que la de los otros grupos en los tres micrositios (Figura 1).
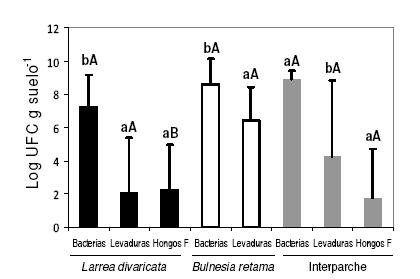
Figura 1. Abundancia total de bacterias, levaduras y hongos filamentosos aislados de los suelos de parches (L. divaricata, B. retama) e interparches. Las barras de error representan 1 error estándar. En letras minúsculas se indican diferencias significativas entre grupos de microorganismos para un mismo micrositio (P=0.006, P=0.03, P=0.007, respectivamente). En letras mayúsculas se indican diferencias significativas entre micrositios para un mismo grupo de microorganismos (P=0.03, P=0.05).
Figure 1. Total abundance of bacteria, yeasts and filamentous fungi isolated from soil of patches (L. divaricata and B. retama) and interspaces. Error bars show 1 standard error from the media. Significant differences between microorganisms groups for each microsite is indicated in letters (P=0.006, P=0.03, P=0.007). Significant differences between microsites for each group of microorganisms is indicated in capital letters (P=0.03, P==0.05).
La abundancia de bacterias en las hojas de B. retama fue significativamente mayor que en las hojas de L. divaricata. Los análisis de hojas de L. divaricata mostraron que la abundancia de los hongos filamentosos y bacterias fue significativamente mayor que la de levaduras. En las hojas de B. retama la abundancia de bacterias fue significativamente mayor que la de hongos filamentosos, donde no se aislaron levaduras (Figura 2).

Figura 2. Abundancia total de bacterias, levaduras y hongos filamentosos aislados de muestras de hojas de L. divaricata y B. retama. Las barras de error denotan el error estándar de la media. En letras minúsculas se indican diferencias significativas entre grupos de microorganismos para un mismo micrositio (P=0.006 y P=0.0008, repectivamente). En letras mayúsculas se indican diferencias significativas entre micrositios para un mismo grupo de microorganismo (P=0.002, P<0.05).
Figure 2. Total abundance of bacteria, yeasts and filamentous fungi isolated from soil of patches (L. divaricata and B. retama) and interspaces. Error bars show the standard error from the media. Significant differences between microorganisms groups for each microsite is indicated in letters (P=0.006 and P=0.0008). Significant differences between microsites for each group of microorganisms is indicated in capital letters (P=0.002, P<0.05).
En los parches de L. divaricata las bacterias asociadas al suelo fueron significativamente más abundantes que las asociadas a sus hojas (P=0.003), mientras que las levaduras y los hongos filamentosos no presentaron diferencias (P=0.8 y P=0.06, respectivamente) (Figuras 1 y 2). En los parches de B. retama la abundancia de bacterias y levaduras en el suelo fue mayor que la de las hojas (P=0.01 y P=0.0001, respectivamente), mientras que los hongos filamentosos no mostraron diferencias de abundancia entre suelo y hojas (P=0.5) (Figuras 1 y 2).
La proporción de colonias de bacterias aisladas con actividad enzimática fue mayor en los parches de L. divaricata y B. retama que en los interparches (Figura 3). En los parches de L. divaricata, 12.8% de colonias de bacterias presentó actividad xilanolítica y 2.2% amilolítica; sin embargo, en los parches de B. retama 7.5% de las colonias de bacterias presentó actividad amilolítica, 4.6% actividad xilanolítica y 0.4% ambas actividades enzimáticas. En los interparches, 1.1% de las colonias de bacterias aisladas presentó actividad xilanolítica y 4.9% tanto actividad xilanolítica como amilolítica (Figura 3). Las hojas de L. divaricata presentaron bacterias con las tres actividades enzimáticas estudiadas, mientras que no encontramos bacterias con actividad enzimática en las hojas de B. retama (Figura 3).

Figura 3. Bacterias aisladas que expresaron actividades enzimáticas en suelos de parches, interparches y hojas de Larrea divaricata y Bulnesia retama.
Figure 3. Isolated bacteria expressing enzymatic activities in soil patches, interspaces and leaves of Larrea divaricata and Bulnesia retama.
El pH y el contenido de humedad del suelo no difirieron entre los distintos tipos de parche (P=0.14 y P=0.43, respectivamente, P=0.05, Tabla 1).
DISCUSIÓN
Los resultados de este estudio no mostraron una influencia de la presencia y el tipo de vegetación sobre la abundancia de bacterias y levaduras del suelo circundante (Figura 1). Esto puede deberse a la baja estabilidad que presenta el sistema por tratarse de un suelo cuya estructura no está formada por horizontes. También puede influir la época de toma de muestras (bajas precipitaciones) donde el sustrato está seco y desagregado. Esta condición facilita su dispersión por el viento, que es el principal factor modelador del paisaje, y puede conducir a la remoción de la parte superficial del suelo y a una relativa homogeneización del sustrato (De Fina 1992; Strahler 1992; Abraham et al. 2009; Abril et al. 2009). Sólo los hongos filamentosos asociados al parche L. divaricata fueron más abundantes respecto de los cuantificados en el parche B. retama e interparches. No se cuantificaron hongos filamentosos cultivables en el parche de B. retama por medio de la metodología utilizada. Este hecho puede tener relación con la abundancia de bacterias en las hojas de L. divaricata ya que esta fue significativamente menor respecto que la cuantificada en las de B. retama (Figura 2). La presencia relativamente baja de bacterias en la canopia de L. divaricata podría permitir un mayor desarrollo de microorganismos capaces de competir por el mismo tipo de sustrato como es el caso de los hongos filamentosos. Esta situación podría verse reflejada en el parche asociado con el vegetal. La mayor abundancia de bacterias cuantificada en la canopia de B. retama respecto de L. divaricata (Figura 2), puede tener explicación por la presencia de un compuesto denominado ácido nordihidroguayarético (NDGA) que es bactericida y es posible de encontrar en la superficie de las hojas y tallo de L. divaricata (Obermeyer et al. 1995; Stege et al. 2005).
Los resultados del presente estudio en cuanto a la distribución de los grupos microbianos evaluados en parches de suelo e interparches concuerdan con un trabajo llevado a cabo en el Parque Nacional Fray Jorge (ecosistema árido de Chile) donde se encontró que no había diferencias en la abundancia de microorganismos de suelo bajo y fuera de la canopia del arbusto de Adesmia bedwelli (Aguilera et al. 1999). De la misma forma, el trabajo realizado en la reserva de la biósfera de Ñacuñan, que forma parte del desierto del Monte Central (como nuestra área de estudio), en el que se evaluó la heterogeneidad espacial y temporal de las islas de fertilidad bajo y fuera de la canopia de Prosopis flexuosa y L. divaricata, se encontró que la abundancia de bacterias no mostró diferencias bajo la canopia y fuera de cada parche (Abril et al. 2009). Nuestro estudio corrobora lo informado en trabajos previos donde la abundancia de grupos microbianos no está asociada a los parches conocidos como islas de fertilidad.
En este estudio se encontró que los grupos funcionales de bacterias que producen enzimas asociadas a la degradación de material vegetal fueron más abundantes en los parches vegetados que en los interparches (Figura 3). Esto sugiere que podría existir una relación entre la abundancia de algunos grupos funcionales de bacterias con la distribución de la vegetación en el ambiente bajo estudio. Estos resultados coinciden con los de Pucheta et al. (2006), quienes encontraron una mayor velocidad de descomposición de los residuos en los parches de L. divaricata y B. retama que en los interparches. Asimismo, estos resultados también están de acuerdo con otros estudios en los que se encontró una actividad microbiana elevada (respiración y enzimas) en parches vegetados vs. parches sin vegetación (Bolton et al. 1993; Smith et al. 1994; Sponseller & Fisher 2008; Abril et al. 2009).
En nuestro trabajo, las bacterias registraron valores significativamente más altos tanto en los suelos (parches e interparches) como en las canopias bajo estudio (Figuras 1 y 2). Si bien existen investigaciones previas que muestran un incremento de la biomasa fúngica en relación con la bacteriana durante la descomposición de detritos vegetales en suelos, también se registran antecedentes que evidencian que la relación hongos/bacteria puede invertirse en función del incremento de la biomasa de bacterias durante el proceso (Diedhiou et al. 2009). El transcurso de la descomposición implica cambios en la composición química de la materia orgánica en proceso, lo cual afecta en forma directa la sucesión de la comunidad microbiana del suelo (Horwath & Elliott 1996). No obstante, en estudios anteriores realizados en suelos del Chaco también se encontró una abundancia mayor de bacterias vs. hongos filamentosos en asociación con arbustos. Esto se debería a la disponibilidad de compuestos fácilmente degradables, que puede asociarse a la fertilidad del suelo (Abril & Bucher 1999; Abril et al. 2005). Un estudio realizado en los ecosistemas áridos de los Estados Unidos también mostró una mayor abundancia de bacterias que de hongos filamentosos en el suelo debajo del arbusto Artemisia tridentata, en coincidencia con una mayor tasa de descomposición de la materia orgánica (Saetre & Sartk 2005).
Las características físico-químicas del suelo también pueden contribuir al desarrollo bacteriano y determinar su sobreabundancia respecto de otros grupos microbianos. Se ha verificado que la mayoría de los géneros de bacterias se desarrollan mejor a pH neutro y ligeramente alcalino (Bitton et al. 1974; Frioni 1999), como el que fue medido tanto en parches como interparches en el área de este estudio (Tabla 1). El tamaño de las fracciones que forman parte de la textura del suelo también puede contribuir al desarrollo, la distribución y la abundancia de bacterias, tal como lo sugiere un estudio que reveló una mayor abundancia de bacterias del género Holophaga y Acidobacterium en partículas de suelo pequeñas como la arena, que es una de las fracciones más abundantes en nuestra área de estudio (Torsiv & Ovreas 1998).
La menor cantidad de biomasa fúngica (i.e., levaduras, hongos filamentosos) respecto de la bacteriana cuantificada en las hojas de los arbustos estudiados podría deberse a que si bien L. divaricata y B. retama poseen exudados que posibilitarían el desarrollo de hongos (Martínez & Dalmasso 1992; Martínez 1993), la combinación de contenidos elevados de azúcar con taninos y otras sustancias tóxicas en los exudados puede disminuir la variedad y la cantidad de especies asociadas a esos microambientes. Esto sería así debido a la condición de osmotolerancia que deben poseer los microorganismos para proliferar en la superficie de los vegetales (Spencer et al. 1995, 1996).
Los resultados obtenidos en el presente trabajo muestran que en los parches existe una mayor abundancia de bacterias que en las hojas de las canopias de los vegetales, como así también mayor porcentaje de colonias de bacterias que presentaron alguna de las actividades enzimáticas estudiadas, involucradas en la descomposición de materia orgánica. El fraccionamiento físico, mecánico y biológico del material vegetal (que aumenta la superficie de contacto) posibilitaría el desarrollo de bacterias con actividades hidrolíticas asociadas a la degradación (Pavel et al. 2004; Pucheta et al. 2006; Carrera et al. 2009). Este estudio constituye la primera contribución al conocimiento de la abundancia de grupos microbianos, como así también de bacterias con actividades enzimáticas en suelos asociados y hojas de canopias de estos dos conspicuos géneros de la familia Zigofilácea e interparches.
BIBLIOGRAFÍA
1. ABRAHAM, E; HF DEL VALLE; F ROIG; L TORRES; JO ARES; ET AL. 2009. Overview of the geography of the Monte Desert biome (Argentina). J. Arid Environ., 73:144-153. [ Links ]
2. ABRIL, A & EH BUCHER. 1999. The effects of overgrazing on soil microbial community and fertility in the Chaco dry savannas of Argentina. Appl. Soil Ecol., 12:159-167. [ Links ]
3. ABRIL, A; P BARTTFELD & EH BUCHER. 2005. The effects of fire and overgrazing disturbs on soil carbon balance in the dry Chaco forest. Forest Ecol. & Manag., 206:399-405. [ Links ]
4. ABRIL, A; P VILLAGRA & L NOE. 2009. Spatiotemporal heterogeneity of soil fertility in the Central Monte desert (Argentina). J. Arid Environ., 73:901-906. [ Links ]
5. AGUIAR, MR & OE SALA. 1999. Patch structure, dynamics and implications for the functioning of arid ecosystems. Tree, 4:273-277. [ Links ]
6. AGUILERA, EL; JR GUTIÉRREZ & PL MERSERVE. 1999. Variation in soil micro-organisms and nutrients underneath and outside the canopy of Adesmia bedwellii (Papilionaceae) shrubs in arid coastal Chile following drought and above average rainfall. J. Arid Environ., 42:61-70. [ Links ]
7. ALEF, K; P NANNIPIERE & C TRAZAR-CEPADA. 1995. Phosphatase activity. Pp. 335-344 en: Alef, K & P Nannipieri (eds.). Methods in Appl. Soil Microbiol. & Biochem. Academic Press, London, UK. [ Links ]
8. AON, MA; MN CABELLO; DE SARENA; AC COLANERI; MG FRANCO; ET AL. 2001. Spatio-temporal patterns of soil microbial and enzymatic activities in an agricultural soil. Appl. Soil Ecol., 18:239-254. [ Links ]
9. BADIANE, NNY; JL CHOTTE; E PATE; D MASSE & C ROULAND. 2001. Use of soil enzyme activities to monitor soil quality in natural and improved fallows in semi-arid tropical regions. Appl. Soil Ecol., 18:229-238. [ Links ]
10. BISIGATO, AJ; PE VILLAGRA; JO ARES & BE ROSSI. 2009. Vegetation heterogeneity in Monte Desert ecosystems: A multi-scale approach linking patterns and processes J. Arid Environ., 73:182-191. [ Links ]
11. BITTON, G; N LAHAV & Y HENIS. 1974. Movement and retention of Klebsiella aerogenes in soil columns. Plant & Soil, 40:373-380. [ Links ]
12. BOLTON, H; JL SMITH & SO LINK. 1993. Soil microbial biomass and activity of a disturbed and undisturbed shrub-steppe ecosystem. Soil Biol. & Biochem., 25:545-552. [ Links ]
13. CARRERA, AL; MJ MAZZARINO; MB BERTILLER; HF VALLE & EM CARRETERO. 2009. Plant impacts on nitrogen and carbon cycling in the Monte Phytogeographical Province, Argentina. J. Arid Environ., 73:192-201. [ Links ]
14. CABRERA, AL & A WILLINK. 1980. Regiones biogeográficas en América Latina. En: Eva Chesneau (eds.). Biogeografía de América Latina. Secretaría de la Organización de los Estados Americanos. Programa de Desarrollo Científico y Tecnológico, Capítulo 8. Serie de Biología. Monografía N° 13:29-107. [ Links ]
15. CORTÉS, M & J HUNZIKER. 1997. Isozymes in Larrea divaricata and Larrea tridentata (Zygophyllaceae): A study of two amphitropical vicariants and autopolyploidy. Genetica, 101:115-124. [ Links ]
16. DAVISON, J. (1988). Plant benefical bacteria. Biotechnol., 6:282-286. [ Links ]
17. DE FINA, AL. 1992. Aptitud Agroclimática de la República Argentina. Academia Nacional de Agronomía y Veterinaria Buenos Aires. Pp. 402. [ Links ]
18. DENARIE, J; F DEBELLE & C ROSENBERG. 1992. Signaling and host range variation in inoculation. Ann. Rev. Microbiol., 46:487-531. [ Links ]
19. DENG, SP & MA TABATABAI. 1995. Cellulase activity of soils: Effect of trace elements. Soil Biol. & Biochem., 27:977-979. [ Links ]
20. DEKKER, RFH & GN RICHARDS. 1976. Hemicellulases, their ocurrence, purification, properties and mode of action. Advances in Carbohydrates Chemistry & Biochemistry, 32:277-352. [ Links ]
21. DIEDHIOU, S; EL DOSSA; AN BADIANE; I DIEDHIOU; M SÈNE; ET AL. 2009. Decomposition and spatial microbial heterogeneity associated with native shrubs in soils of agroecosystems in semi-arid Senegal. Pedobiologia, 52:273-286. [ Links ]
22. DI RIENZO, J; M BALZARINI; F CASANOVES; L GONZÁLEZ; M TABLADA; ET AL. 2002. InfoStat Estudiantil versión 2.0. Universidad Nacional de Córdoba. Estadística y Diseño. F.C.A. [ Links ]
23. DICK, RP; D BREAKWILL & R TURCO. 1996. Soil enzyme activities and biodiversity measurements as integrating biological indicators. Pp. 247-272 en: Doran, JW & AJ Jones (eds.). Handbook of Methods for Assessment of Soil Quality. Soil Sci. Soc. Am., Madison. [ Links ]
24. EDITORIAL COMMITTEE. 1996. Soil physical and chemical analysis and description of soil profiles. Standards Press of China, Beijing, China (in Chinese). [ Links ]
25. FARNSWORTH, RB; EM ROMMEY & A WALLACE. 1977. Nitrogen fixation by microfloral- higher plant associations in arid to semiarid environments. Pp. 17-19 en: West, NE & JJ Skujins (eds.). Nitrogen in Desert Ecosystems. Stroudsburg, PA. Dowden, Hutchinson & Ross Inc. USA. [ Links ]
26. FRIONI, L.1999. Crecimiento microfifbiano y su control. Efecto de factores ambientales. Capítulo 4:65-85. Microbiología básica, ambiental y Agrícola. Editorial de la Fundación Universidad Nacional de Rio Cuarto. Argentina. ISBN 950-885110. [ Links ]
27. GONZÁLEZ-POLO, M & A AUSTIN. 2009. Spatial heterogeneity provides organic matter refuges for soil microbial activity in the Patagonian steppe, Argentina. Soil Biol. & Biochem., 41:1348-1351. [ Links ]
28. HMERLYNCK, E; J MCAULIFFE; E MCDONALD & S SMITH. 2002. Ecological responses of two Mojave desert shrubs to soil horizon development and soil water dynamics. Ecology, 83(3):768-779. [ Links ]
29. HORWATH, WR & LF ELLIOTT. 1996. Ryegrass straw component decomposition during mesophilic and thermophilic incubations. Biol.Fertil.Soils, 21:227-232. [ Links ]
30. KURTZMAN, CP. 2000. Three new ascomycetous yeasts from insect-associated to arboreal habitats. Canadian J. Microbiol., 46:50-58. [ Links ]
31. LEE, H; P BIELY; RK LATTA; MFS BARBOSA & H SCHNEIDER. 1986. Utilization of xylan by yeasts and its conversion to ethanol by Pichia stipitis strains. Appl. Environ. Microbiol., 52:320-324. [ Links ]
32. MAMILOV, ASH & OM DILLY. 2002. Soil microbial ecophysiology as affected by short-term variations in environmental conditions. Soil Biol. & Biochem., 34:1283-1290. [ Links ]
33. MARTÍNEZ CARRETERO, E. 1993. Regional development and desertification control through ecologicalk farming: three examples in Argentina. Desertification Bull., 23:35-38. [ Links ]
34. MARTÍNEZ CARRETERO, E & AD DALMASSO. 1992. Litter yield in shrubs of Larrea in the Andean piedmont of Mendoza, Argentina. Vegetation, 101:21-33. [ Links ]
35. NAKAS, JP & DA KLEIN. 1980. Mineralization capacity of bacteria and fungi from the rhizosphere-rhizoplane of a semiarid grassland. App. Environm. Microbiol., 39:113-117. [ Links ]
36. OBERMEYER, WR; SM MUSSER; JM BETZ; RE CASEY; AE POHLAND; ET AL. 1995. Chemical studies of phytoestrogens and related compounds in dietary supplements: Flax and Chaparral. Proc. Soc. Exp. Biol. Med., 208:6-12. [ Links ]
37. PALACIOS, RA & JH HUNZIKER. 1984. Revisión taxonómica del género Bulnesia (Zygophyllaceae). Darwiniana, 25:299-320. [ Links ]
38. PAVEL, R; J DOYLE & Y STEINBERGER. 2004. Seasonal patterns of cellulase concentration in desert soil. Soil Biol. & Biochem., 36:549-554. [ Links ]
39. PUCHETA, E; M LLANOS; C MEGLIOLI; M GAVIORNO; M RUIZ; ET AL. 2006. Litter decomposition in a sandy Monte desert of western Argentina: Influences of vegetation patches and summer rainfall. Austral Ecol., 31:808-816. [ Links ]
40. RIBAS-FERNÁNDEZ, Y; L QUEVEDO-ROBLEDO & E PUCHETA. 2009. Pre- and post-dispersal seed loss and soil seed dynamics of the dominant Bulnesia retama (Zygophyllaceae) shrub in a sandy Monte desert of western Argentina. J. Arid Environ., 73: 14-21. [ Links ]
41. RUNDEL, P; PE VILLAGRA; MO DILLON; SA ROIGJUÑENT& G DEBANDI. 2007. Arid and Semi-Arid Ecosystems. Pp. 158-183 en: Veblen, TT; K Young; & AE Orme (eds.). The Physical Geography of South America. Oxford University Press. [ Links ]
42. SAETRE, P & M SARTK. 2005. Microbial dynamics and carbon and nitrogen cycling following re-wetting of soils beneath two semi-arid plant species. Oecologia, 142:247-260. [ Links ]
43. SCHLESINGER, WH & AM PILMANIS. 1998. Plant-soil interactions in deserts. Biogeochem., 42:169-187. [ Links ]
44. SCHLOTER, M; O DILLY & JC MUNCH. 2003. Indicators for evaluating soil quality. Agriculture, Ecosystems & Environ., 98:255-262. [ Links ]
45. SMITH, JL; JJ HALVORSON & H BOLTON. 1994. Spatial relationships of soil microbial biomass and C and N mineralization in a semi-arid shrub-steppe ecosystem. Soil Biol. & Biochem., 26:1151-1159. [ Links ]
46. SPENCER, DM; JFT SPENCER; E FENGLER & LI DE FIGUEROA. 1995. Yeasts associated with algarrobo trees (Prosopis spp.) in northwest Argentina: a preliminary report. J. Ind.Microbiol., 14:472-474. [ Links ]
47. SPENCER, DM; JFT SPENCER; LI DE FIGUEROA; O GARRO & E FENGLER. 1996. Yeasts associated with pods and exudates of algarrobo trees (Prosopis spp.) and species of columnar cacti in northwest Argentina. Appl. Microbiol. Biotechnol., 44:736-739. [ Links ] [ Links ]
49. STEGE, PW; RC DAVICINO; AE VEGA; YA CASALIB; S CORREACI; ET AL. 2005. Antimicrobial activity of aqueous extracts of Larrea divaricata Cav (jarilla) against Helicobacter pylori. Phytomedicine, 13:724-727. [ Links ]
50. STRAHLER, AN. 1992. Geología Física. Editorial Omega. Pp. 510. [ Links ]
51. STRAUSS, MLA; NP JOLLY; MG LAMBRECHTS & P VAN RENSBURG. 2001. Screening for the production of extracellular hydrolytic enzymes by non-Sacchraromyces wine yeasts. J. Appl. Microbiol., 91:182-190. [ Links ]
52. TORRES, PA & A ABRIL. 2005. Microbial succession in litter decomposition in the semi-arid Chaco woodland. Soil Biol. & Biochem., 37:49-54. [ Links ]
53. TORO, ME; NP ORO; AD VEGA; MC NALLY; YP MATURANO; ET AL. 2005. Diversidad de levaduras en canopias y suelos asociados a Bulnesia retama y Larrea divaricata. Revista Argentina de Microbiología, 37:209-213. [ Links ]
54. TORSVIK, V; FL DAAE; RA SANDAA & L OVREAS. 1998. Review article: Novel techniques for analyzing microbial diversity in natural and perturbed environments. J. Biotechnol., 64:53-62. [ Links ]
55. VÁZQUEZ, F; MD VALLEJO HERRERA; LIC DE FIGUEROA & ME TORO. 2004. Extracellular hydrolytic enzymes produced by yeasts. Pp. 283-298 en: Spencer, JFT & ALR de Spencer (eds.). Environmental Microbiology. Methods and Protocols. Vol. 16, chapter 30. Humana Press, Totowa, New Jersey. E-ISBN 1-58829-116- 2. [ Links ]
56. VILELA, A; ML BOLKOVIC; P CARMANCHAHI; M CONY; D DE LAMO; ET AL. 2009. Past, present and potential uses of native flora and wildlife of the Monte Desert. J. Arid Environ., 73:238-243. [ Links ]
57. VISHNEVESTKY, S & Y STEINBERGERT. 1997. Bacterial and fungal dynamics and their contribution to microbial biomass in desert soil. J. Arid Environ., 37:83-90. [ Links ]
58. WEAVER, RW; JS ANGLE & PS BOTTOMLEY. 1994. Methods of Soil Analysis. Part 2. Microbiological & Biochemical Properties. N°. 5. Soil Sci. Soc. Am., Madison. USA. [ Links ]

