










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.20 no.3 Córdoba sep./dic. 2010
TRABAJOS ORIGINALES
Diversidad florística bajo diferentes intensidades de pastoreo por grandes herbívoros en pastizales serranos del Sistema de Ventania, Buenos Aires
Alejandro Loydi1,* & Roberto A Distel1,2
1 Centro de Recursos Naturales Renovables de la Zona Semiárida (CERZOS), CCT CONICET Bahía Blanca, Universidad Nacional del Sur, Bahía Blanca, Argentina.
2 Departamento de Agronomía, Universidad Nacional del Sur. Bahía Blanca, Argentina.
* Centro de Recursos Naturales Renovables de la Zona Semiárida (CERZOS), CCT CONICET Bahía Blanca, Universidad Nacional del Sur, Bahía Blanca, Argentina. Tel.: +54-0291-4861124, Fax: +54-0291- 4861527. Email: aloydi@criba.edu.ar.
Recibido: 14 de julio de 2010;
Fin de arbitraje: 12 de septiembre de 2010;
Revisión recibida: 16 de septiembre de 2010;
Aceptado: 13 de octubre de 2010
RESUMEN. En pastizales de regiones subhúmedas, el pastoreo moderado de grandes herbívoros promovería la diversidad florística. Nuestro objetivo fue evaluar la diversidad florística y la composición botánica en diferentes intensidades de pastoreo por grandes herbívoros en pastizales serranos del Sistema de Ventania, Buenos Aires, Argentina. El muestreo abarcó un área de 200 ha, y comprendió comunidades de flechillas ("flechillales") sometidas al pastoreo de caballos cimarrones y ganado vacuno. Se seleccionaron nueve parcelas de 1 ha sometidas a diferentes intensidades de pastoreo. Sobre la base de la estimación de la cobertura de cada una de las especies presentes se describió el patrón de diversidad florística, se comparó la cobertura y riqueza de especies, y se evaluaron los cambios en la composición florística, en relación al gradiente de intensidad de pastoreo. El disturbio por pastoreo afectó la riqueza específica de los flechillales y la máxima riqueza se registró en intensidades de pastoreo intermedias. Dicha respuesta se debió principalmente a un mayor número de especies latifoliadas y de gramíneas invernales. En términos de cobertura aérea, el pastoreo favoreció a las latifoliadas únicamente. El pastoreo modificó la composición botánica de los flechillales, apareciendo como más notable el reemplazo de gramíneas invernales (Piptochaetium hackelii, Briza subaristata, Nassella filiculmis) por una gramínea estival de palatabilidad baja (Aristida spegazzinii) a intensidades elevadas de pastoreo. En los pastizales serranos del Sistema de Ventania, el pastoreo moderado favorecería la diversidad florística, aumentando la riqueza de especies de latifoliadas y gramíneas invernales mientras que las altas intensidades favorecerían el reemplazo de las gramíneas invernales palatables por gramíneas de palatabilidad menor.
Palabras clave: Riqueza específica; Composición botánica; Disturbio; Flechillales; Parque Provincial Ernesto Tornquist.
ABSTRACT. Floristic diversity under different intensities of large herbivore grazing in mountain grasslands of the Ventania System, Buenos Aires: In sub-humid grasslands moderate grazing by large herbivores may promote increases in plant diversity. Our objective was to evaluate changes in floristic diversity and floristic composition at different grazing intensities by large herbivores in mountain grasslands of the Ventania System, Buenos Aires, Argentina. Sampling comprised an area of 200 ha, on needlegrass grasslands grazed by feral horses and cattle. Nine 1 ha plots were selected to represent a grazing intensity gradient, and we evaluated plant species cover in order to describe the plant diversity pattern, species richness and cover, and floristic composition in relation to the grazing intensity gradient. Plant diversity was greater at moderate grazing intensity mainly due to the increment of dicots and cool-season grasses richness. In terms of cover, grazing only increased dicots' aerial cover. Grazing was associated with changes in the floristic composition of needlegrass grasslands, through inducing the replacement of cool-season grasses (e.g. Piptochaetium hackelii, Briza subaristata, Nassella filiculmis) by an unpalatable warm-season grass (Aristida spegazzinii). In mountain grasslands of the Ventania System, moderate grazing intensity appears to favor plant diversity by increasing dicots and cool-season grasses richness, whereas high grazing intensity may promote the replacement of palatable cool-season grasses by unpalatable warm season grasses.
Keywords: Species richness; Botanical composition; Disturbance; Needlegrass; Parque Provincial Ernesto Tornquist.
INTRODUCCIÓN
La diversidad florística constituye una característica importante de los ecosistemas, tanto por sus implicancias para la conservación de especies como por su rol en el funcionamiento de ecosistemas (Chapin III et al. 2000; Tilman 2000; Hooper et al. 2005). Por lo general, la diversidad vegetal incrementa la eficiencia de uso de los recursos y contribuye a estabilizar el funcionamiento de los ecosistemas frente a factores de estrés o disturbio (Naeem et al. 2000; Hooper et al. 2005). Por otra parte, si bien una alta diversidad de flora nativa puede ir acompañada por un incremento de especies exóticas ("paradoja de la invasión"), una reducción en la diversidad de las especies nativas aceleraría el proceso de invasión de especies exóticas (Fridley et al. 2007).
En ecosistemas de pastizales naturales el disturbio provocado por el pastoreo de grandes herbívoros constituye uno de los principales determinantes de la diversidad florística (Milchunas et al. 1988). El pastoreo afecta la diversidad vegetal a través de modificaciones del balance entre la colonización y extinción localizadas de especies (Olff & Ritchie 1998). La colonización depende de la disponibilidad y diversidad de propágulos en el banco de semillas del suelo, y de la disponibilidad de sitios seguros para el establecimiento de plántulas. En tanto la exclusión competitiva y la sensibilidad al pastoreo determinan la extinción localizada de especies a bajas y altas intensidades de pastoreo, respectivamente Así, en pastizales de regiones sub-húmedas el pastoreo moderado, en intensidad tal que reduzca la competencia de las especies dominantes sin comprometer la persistencia de las especies menos resistentes a las defoliaciones, incrementaría la diversidad florística. Estos cambios en diversidad afectan no sólo el número de especies presentes y su abundancia (Pykälä 2005a; Stockton et al. 2005), sino también la composición específica de las comunidades (Wang et al. 2002; Altesor et al. 2006); esto favorece ciertos tipos funcionales de plantas por sobre otros (Lavorel et al. 1999; Altesor et al. 2006; Díaz et al. 2007).
En pastizales naturales de la Pampa Deprimida el pastoreo incrementa la diversidad vegetal a través de la promoción de especies nativas latifoliadas y especies exóticas (Sala et al. 1986; Chaneton et al. 2002; Rusch & Oesterheld 1997), mientras tiende a homogeneizar la composición florística y funcional a escala de paisaje (Chaneton et al. 2002). de manera similar, en los pastizales pampeanos serranos del Sistema de Ventania el pastoreo de grandes herbívoros modifica la estructura de las comunidades (Scorolli 1999; Zalba & Cozzani 2004), incrementando la abundancia de gramíneas de baja palatabilidad, arbustos y especies exóticas (Frangi & Bottino 1995; Long & Grassini 1997). Sin embargo, se desconoce el patrón de diversidad vegetal y las modificaciones en la composición florística a lo largo de un gradiente de intensidad de pastoreo, en estos pastizales que ocupan grandes extensiones de la región. Por lo general, los trabajos sobre el tema se limitan a comparar áreas clausuradas al pastoreo yáreas vecinas sujetas al pastoreo típico de la zona. Por el contrario, en el presente estudio se intentó establecer una relación general entre el pastoreo y la diversidad florística utilizando un gradiente natural construido a partir de nueve sitios con diferente historia reciente de uso pastoril. El objetivo del presente trabajo fue evaluar la diversidad y composición florística a diferentes intensidades de pastoreo por grandes herbívoros, caballos cimarrones y ganado vacuno.
MÉTODOS
Área de estudio
El área de estudio comprendió ambientes pedemontanos (500 m.s.n.m.) en las Sierras Australes del Sistema de Ventania, Buenos Aires, Argentina. El clima de la región es templado, subhúmedo. La temperatura media anual del aire es 14 °C y el promedio anual de precipitaciones 800 mm (Burgos 1968). En el año de estudio (2007) las precipitaciones fueron de 954 mm. Los suelos dominantes se clasifican en Hapludoles y Argiudoles líticos, con un contenido de materia orgánica elevado (=7%) en los horizontes superficiales (Cappannini et al. 1971; Frangi et al. 1980). En lugares excluidos del pastoreo por grandes herbívoros, el pastizal forma un tapiz herbáceo denso de 50 a 60 cm de altura y está dominado por Piptochaetium hackelii (Arechav.) Parodi, Piptochaetium napostaense (Speg.) Hack., Nassella melanosperma (J. Presl) Barkworth y Briza subaristata Lam. (en adelante, flechillales) (Frangi & Bottino 1995). La nomenclatura siguió a Zuloaga & Morrone (2007).
El trabajo se realizó en la Reserva Integral del Parque Provincial Ernesto Tornquist (Parque Tornquist en adelante) (38º 03' S y 61º 59' W) y en una estancia aledaña (Ea. Palo Alto). Ambos lugares han estado históricamente pastoreado por grandes herbívoros, con predominio de caballos cimarrones en el Parque Tornquist y de ganado vacuno en la Estancia Palo Alto.
Diseño del muestreo y análisis estadístico
Las mediciones se realizaron al final de la primavera de 2006 y principios del verano de 2007, en un área de 200 ha (180 ha en el Parque Tornquist y 20 ha en la Estancia Palo Alto). A partir de indicadores de la intensidad de pastoreo se seleccionaron nueve parcelas de muestreo de 1 ha (siete en el Parque Tornquist y dos en la Ea. Palo Alto), que representaban un gradiente de intensidad de pastoreo. Para asegurar la independencia de muestreo y aumentar el grado de control en las comparaciones a realizar entre las diferentes intensidades, las parcelas estuvieron separadas entre sí por no menos de 250 m y se ubicaron en sitios con nivel y orientación de pendiente similar (~5% y orientación Norte, respectivamente).
Los indicadores de la intensidad de pastoreo se midieron en sub-parcelas de 1 m2 (n=30) distribuidas al azar dentro de cada parcela de muestreo, e incluyeron el porcentaje de suelo desnudo, altura y diámetro basal de matas individuales de flechillas (Piptochaetium spp. y Nassella spp.), altura media ponderada de la vegetación tomada a intervalos de 5 cm y cobertura y frecuencia de aparición de estiércol, que son parámetros frecuentemente afectados por la intensidad de pastoreo (Cingolani et al. 2003; Teague et al. 2004). Con los datos obtenidos, y mediante análisis de componentes principales, se obtuvo una variable que representó la intensidad de pastoreo en cada parcela. Esta nueva variable, representada por el primer componente principal del análisis, explicó 89% de la varianza total (Figura 1). La altura de matas de flechillas, la altura media ponderada de la vegetación y el diámetro basal de las matas de flechillas se correlacionaron de forma negativa con el primer componente principal (r=-0.983, P<0.01; r=-0.979, P<0.01; r=-0.903, P<0.01, respectivamente); mientras que las variables asociadas a mayores intensidades de pastoreo, como la cobertura y frecuencia de aparición del estiércol y el porcentaje de suelo desnudo se correlacionaron de forma positiva con dicho componente (r=0.921, P<0.01; r=0.954, P<0.01; r=0.927, P<0.01, respectivamente). Las parcelas seleccionadas en la Estancia Palo Alto se ubicaron en el extremo positivo del gradiente de intensidad de pastoreo (Figura 1), junto a tres de las parcelas seleccionadas en el Parque Tornquist. Las restantes parcelas seleccionadas en el Parque Tornquist quedaron ubicadas en el extremo negativo del gradiente.

Figura 1. Gradientes de intensidad de pastoreo construidos a partir del análisis de componentes principales. Las flechas señalan las dos áreas de muestreo en la Estancia Palo Alto.
Figure 1. Grazing intensity gradient built from principal components analysis. Arrows point the two sampling areas at Palo Alto Ranch.
En cada parcela de muestreo se dispusieron de manera aleatoria subparcelas de 1 m2 (n=15), diferentes a las utilizadas para medir los indicadores de la intensidad de pastoreo, y se estimó de manera visual la cobertura aérea de todas las especies presentes. Se utilizó una modificación de la escala de Braun-Blanquet (Sutherland 1996), con incrementos de 10% para coberturas por encima de 5%. Cabe aclarar que, si bien uno de los parámetros utilizados para definir la intensidad de pastoreo fue el diámetro basal de matas individuales de flechillas (Piptochaetium spp. y Nassella spp.), dicha variable no necesariamente se correlaciona con la cobertura aérea a nivel poblacional. Por lo general, el pastoreo disminuye el diámetro basal de las matas e incrementa su densidad (Sala et al. 1986), sin afectar o afectando en menor medida su cobertura aérea. Los datos obtenidos se analizaron mediante análisis de regresión simple lineal o polinomial (Zar 1999). Se realizaron regresiones entre riqueza específica y el gradiente de intensidad de pastoreo. De la misma manera se comparó la riqueza y cobertura de grupos de especies, tales como arbustivas (arbustos o sub-arbustos con crecimiento secundario), graminoides (Ciperáceas, Lilifloras y Juncos), latifoliadas (dicotiledóneas herbáceas), pastos invernales, pastos estivales y suculentas (Cactus y afines). Por último, la cobertura de las gramíneas más comunes en el pastizal (aparición en al menos 2/3 de las áreas muestreadas) se evaluó a lo largo del gradiente de intensidad de pastoreo. Previo a la realización de todos los análisis, los datos de riqueza fueron transformados a raíz cuadrada y los datos de cobertura a arcoseno de la raíz cuadrada para cumplir con los supuestos de normalidad y homocedasticidad (Zar 1999). Por último, mediante un análisis de componentes principales se evaluaron las diferencias en la composición florística de las distintas áreas. Los datos fueron transformados previamente a arco-seno de la raíz cuadrada, y el análisis se llevó a cabo utilizando las matrices de covarianza (Legendre & Legendre 1998). Se realizó un análisis de correlación entre la disposición de las áreas en el gradientes de intensidad de pastoreo y la disposición de estas mismas áreas en el análisis de componentes principales proveniente de los datos de composición.
RESULTADOS
La riqueza específica se ajustó a un modelo cuadrático, tanto para las especies nativas como para el total de las especies (nativas + exóticas) (F(1,6)=9.60, r2=0.653, P<0.05, y F(1,6)=15.59, r2=0.731, P<0.01, respectivamente) (Figura 2).

Figura 2. Relación entre la riqueza de especies en 15 m2 y el gradiente de intensidad de pastoreo. Se muestran los datos transformados a raíz cuadrada. Los símbolos vacíos representan al total de especies, mientras que los símbolos rellenos sólo a las especies nativas.
Figure 2. Relationship between species richness in 15 m2 and the grazing intensity gradient. Square root transformed data are shown. Empty symbols represent total species richness and filled symbol represent native species richness.
La riqueza de latifoliadas (Figura 3c) aumentó a intensidades intermedias de pastoreo (F(2,6)=5.16, P<0.05), en tanto la riqueza de pastos invernales disminuyó significativamente al aumentar la intensidad (F(1,7)=10.55, P< 0.05) (Figura 3d). La riqueza de los demás grupos florísticos no varió a lo largo del gradiente de intensidad de pastoreo (Figura 3). Por su parte, la cobertura de especies de graminoides (Figura 4b) aumentó al aumentar la intensidad de pastoreo (F(1,7)= 21.25, P< 0.01). La cobertura de latifoliadas (Figura 4c) siguió el mismo patrón que la riqueza, resultando en mayores coberturas a intensidades intermedias de pastoreo (F(2,6)=15.12, P<0.01). Por su parte, en las gramíneas invernales (Figura 4d) se observó una disminución significativa de su cobertura al aumentar la intensidad (F(1,7)=8.18, P< 0.05). La cobertura de los demás grupos vegetales no varió a lo largo del gradiente de pastoreo (Figura 4).
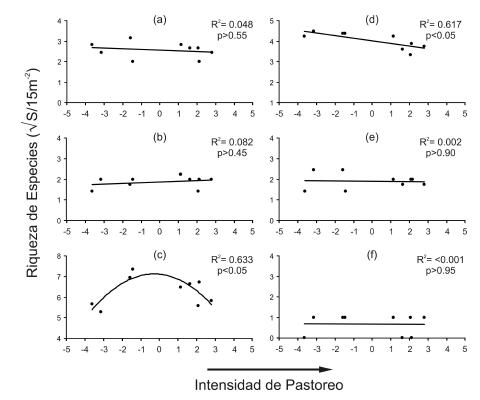
Figura 3. Relación entre la riqueza de especies de los diferentes grupos vegetales a lo largo del gradiente de intensidad de pastoreo. Se muestran los datos transformados a raíz cuadrada. Se detalla el coeficiente de determinación (R2) y la probabilidad de rechazo par a la curva de regresión. (a) Arbustivas, (b) graminoides, (c) latifoliadas, (d) pastos invernales, (e) pastos estivales, (f) suculentas.
Figure 3. Relationship between species richness of different vegetation groups and the grazing intensity gradient. Square root transformed data are shown. Coefficient of determination (R2) and probability error for regression curves are shown. (a) Shrubs, (b) graminoids, (c) forbs, (d) cool-season grasses, (e) warm-season grasses, (f) succulents.

Figura 4. Relación entre el porcentaje de cobertura de los diferentes grupos vegetales a lo largo del gradiente de intensidad de pastoreo. Se muestran los datos transformados a arco-seno raíz cuadrada. Se detalla el coeficiente de determinación (R2) y la probabilidad de rechazo par a la curva de regresión. (a) Arbustivas, (b) graminoides, (c) latifoliadas, (d) pastos invernales, (e) pastos estivales, (f) suculentas.
Figure 4. Relationship between percentage cover of different vegetation groups and the grazing intensity gradient. Arc-Sin square root transformed data are shown. Coefficient of determination (R2) and probability error for regression curves are shown. (a) Shrubs, (b) graminoids, (c) forbs, (d) cool-season grasses, (e) warm-season grasses, (f) succulents.
Al analizar los cambios en la cobertura de las gramíneas más comunes en el pastizal (Figura 5) se observó que el pastoreo aumentó la abundancia de Aristida spegazzinii (F(1,7)=32.34, P<0.01) en desmedro de la abundancia de Calotheca brizoides (F(1,7)=10.6, P<0.05), de Piptochaetium hackelli (F(2,6)=36.12, P<0.01) y de Nassella trichotoma (F(1,7)=5.67, P<0.05). El resto de las especies no mostró cambios estadísticamente significativos en su cobertura en las diferentes intensidades de pastoreo.

Figura 5. Porcentaje de cobertura de las gramíneas dominantes a lo largo del gradiente de intensidad de pastoreo. (a) Aristida spegazzinii, (b) Briza subaristata, (c) Calotheca brizoides, (d) Dantonia cirrata, (e) Piptochaetium hackelii, (f) Piptochaetium medium, (g) Piptochaetium montevidense, (h) Piptochaetium stipoides. (i) Nassella trichotoma. Se muestran los datos transformados a arco-seno raíz cuadrada. Se detalla el coeficiente de determinación (R2) y la probabilidad de rechazo par a la curva de regresión.
Figure 5. Relationship between percentages cover of the dominant grasses under and the grazing intensity gradient. (a) Aristida spegazzinii, (b) Briza subaristata, (c) Calotheca brizoides, (d) Dantonia cirrata, (e) Piptochaetium hackelii, (f) Piptochaetium medium, (g) Piptochaetium montevidense, (h) Piptochaetium stipoides. (i) Nassella trichotoma. Arc-Sin square root transformed data are shown. Coefficient of determination (R2) and probability error for regression curves are shown.
El análisis de componentes principales de la composición de especies presentó un primer componente que explicó 48.8% de la varianza total, mientras que el segundo componente explicó 21.2% de la varianza. La disposición de las áreas a lo largo del primer componente principal se correlacionó de manera positiva con el gradiente de pastoreo (r=0.928, P<0.01), mostrando que la composición específica de los flechillales varió con la intensidad de pastoreo (Figura 5, Tabla 1). A intensidades bajas dominaron Pavonia cymbalaria, Pfaffia gnaphaloides, Piptochaetium hackelii, Nassella melanosperma y Nassella filiculmis; a intensidades intermedias Calotheca brizoides, Eryngium paniculatum, Evolvulus sericeus, Hypochaeris radicata, Margyricarpus pinnatus, Piptochaetium stipoides y Plantago myosuros; y a intensidades altas Adesmia incana, Aristida spegazzinii, Berroa gnaphalioides, Chaptalia piloselloides, Chevreulia sarmentosa, Eryngium nudicaule, Plantago patagonica y Scleranthus annus. Algunas especies (e.g., Briza subaristata, Melica argyrea y Nassella trichotoma) mantuvieron su abundancia a intensidades baja e intermedia, mientras que otras (e.g.,Piptochaetium montevidense) no fueron afectadas por la intensidad de pastoreo.

Figura 6. Disposición de las áreas muestreadas en el espacio de las dos primeras componentes principales. Las letras representan la correlación entre las variables (especies) y los componentes principales. Ai: Adesmia incana, As:Aristida spegazzinii, Bg: Berroa gnaphalioides, Bs: Briza subaristata, Cb: Calotheca brizoides, Ca: Carex spp., Cp: Chaptalia piloselloides, Ch: Chevreulia sarmentosa, Ds: Dichondra sericea, En: Eryngium nudicaule, Ep: Eryngium paniculatum, Es: Evolvulus sericeus, Hr: Hypochaeris radicata, Mp: Margyricarpus pinnatus, Ma: Melica argyrea, Nf: Nassella filiculmis, Nm: Nassella melanosperma, Nt: Nassella trichotoma, Pb: Paronychia brasiliana, Pc: Pavonia cimbalaria, Pg: Pfaffia gnaphaloides, Ph: Piptochaetium hackelii, Ps:Piptochaetium stipoides, Pm: Plantago myosuros, Pp: Plantago patagonica, Sa: Scleranthus annus, Tl: Trifurcia lahue. CP: Componente Principal.
Figure 6. Ordination of sampled areas through principal components analysis. Letters represent correlations between variables (species) and principal components. Species codes as indicated in the legend in Spanish.
DISCUSIÓN
La mayor diversidad florística del pastizal serrano se observó a intensidades de pastoreo intermedias, y, por lo tanto, resultó consistente con la "hipótesis del disturbio intermedio" (Connell 1978) y con modelos de diversidad florística en función del potencial productivo y la historia evolutiva de los pastizales (e.g., Milchunas et al. 1988). Para pastizales productivos de regiones subhúmedas, tal es el caso del pastizal serrano en estudio (Frangi et al. 1980), estos modelos predicen una diversidad de especies máxima a intensidades de pastoreo intermedias, más allá de la historia evolutiva. Aumentos en la diversidad florística con el pastoreo se han observado en pastizales de otras regiones sub-húmedas de la Argentina (Sala et al. 1986; Pucheta et al. 1998; Nai-Bregaglio et al. 2002) y del mundo (Milchunas et al. 1988; Noy-Meir et al. 1989; Rambo & Faeth 1999; Rook et al. 2004). No obstante, con frecuencia los trabajos sobre el tema se limitaron a comparar áreas clausuradas al pastoreo con áreas aledañas sujetas al pastoreo típico de la región, y sin repeticiones reales del "tratamiento" pastoreo. Por el contrario, en el presente estudio se intentó establecer una relación general entre pastoreo y diversidad florística utilizando un gradiente natural construido a partir de nueve sitios con diferente historia reciente de uso pastoril. Por otra parte, un patrón unimodal de diversidad similar se observó en especies de aves que habitan el pastizal serrano del Sistema Ventania (Zalba & Cozzani 2004).
La variación en la diversidad vegetal a diferentes intensidades de pastoreo estuvo determinada en gran medida por las variaciones en las latifoliadas y las gramíneas la exclusión competitiva de las gramíneas invernales dominantes reduciría la riqueza y cobertura de las latifoliadas, mientras que a intensidades altas la severidad de las defoliaciones reduciría la riqueza y cobertura de las gramíneas invernales. Además, a intensidades de pastoreo altas el pisoteo de los animales a través de la compactación del suelo y el daño mecánico reduciría la disponibilidad de sitios seguros para el establecimiento de plántulas (Harper 1977).
Estos cambios estructurales en la vegetación llevaron asociados cambios en la composición de especies. A intensidades de pastoreo altas se observó un aumento en la abundancia de latifoliadas anuales de crecimiento rastrero o en roseta, similar a lo descripto por otros autores (Lavorel et al. 1999; Dupré & Diekmann 2001; Peco et al. 2005; Pykälä 2005b). En general, el pastoreo induce el reemplazo parcial de especies perennes por especies anuales (Todd & Hoffman 1999; Grime 2001; Osem et al. 2002), lo que puede facilitar, entre otras, la invasión de especies exóticas (Milchunas et al. 1988; Richardson et al. 2000; Chaneton et al. 2002; Vavra et al. 2007). Sin embargo, en el presente trabajo no hubo diferencia entre el patrón de diversidad florística descripto para especies nativas y el correspondiente al total de especies (nativas y exóticas), lo cual sugiere que la intensidad de pastoreo no afecta la abundancia de especies exóticas. En un estudio previo, en el mismo pastizal, se observó que el efecto del pastoreo sobre la abundancia de las especies exóticas fue mayor con pastoreo de caballos que con pastoreo de vacunos (Loydi et al. 2010). Esto ayudaría a explicar la independencia entre abundancia de especies exóticas y la intensidad de pastoreo documentada en el presente estudio, ya que las áreas pastoreadas por vacas (n=2) se dispusieron en la zona de mayor intensidad de pastoreo. El bajo impacto del pastoreo vacuno sobre la abundancia de especies exóticas en el pastizal serrano del Sistema Ventania no se condice con los resultados obtenidos en pastizales de otras zonas de la región pampeana, donde se observó un incremento de especies exóticas con el pastoreo vacuno (Chaneton et al. 2002). Diferencias entre pastizales en factores tales como composición botánica, identidad de las especies invasoras, suelo y clima contribuirían a explicar el comportamiento diferencial de las especies exóticas en relación al pastoreo.
También es común que el pastoreo provoque el reemplazo de gramíneas perennes de alta palatabilidad por gramíneas perennes de baja palatabilidad (Westoby et al. 1989; West 1993; Distel & Bóo 1996; Cingolani et al. 2005; Pykälä 2005b; Pazos et al. 2007). En el presente estudio, habiendo seguido un gradiente creciente de íintensidad de pastoreo, se observó el reemplazo de gramíneas palatables (principalmente del género Piptochaetium) por Aristida spegazzinii, gramínea de baja palatabilidad y tamaño de mata reducido (Rúgolo de Agrasar et al. 2005). No obstante, algunas especies del género Piptochaetium (e.g., P. montevidense o P. stipoides) no mostraron cambios significativos de cobertura a lo largo del gradiente de pastoreo. Esto podría atribuirse a la formación de céspedes de pequeñas matas con hojas casi horizontales (A. Loydi, observación personal), que les permitiría evadir el pastoreo de forma similar a lo que fuera señalado para otras especies de pastos (Hickey 1961; Fahnestock & Detling 2000).
También es común que el pastoreo provoque el reemplazo de gramíneas perennes de alta palatabilidad por gramíneas perennes de baja palatabilidad (Westoby et al. 1989; West 1993; Distel & Bóo 1996; Cingolani et al. 2005; Pykälä 2005b; Pazos et al. 2007). En el presente estudio, habiendo seguido un gradiente creciente de íintensidad de pastoreo, se observó el reemplazo de gramíneas palatables (principalmente del género Piptochaetium) por Aristida spegazzinii, gramínea de baja palatabilidad y tamaño de mata reducido (Rúgolo de Agrasar et al. 2005). No obstante, algunas especies del género Piptochaetium (e.g., P. montevidense o P. stipoides) no mostraron cambios significativos de cobertura a lo largo del gradiente de pastoreo. Esto podría atribuirse a la formación de céspedes de pequeñas matas con hojas casi horizontales (A. Loydi, observación personal), que les permitiría evadir el pastoreo de forma similar a lo que fuera señalado para otras especies de pastos (Hickey 1961; Fahnestock & Detling 2000). El mantenimiento de la diversidad vegetal, a nivel de especie o de grupo funcional, cumple un rol sustancial en el funcionamiento y la provisión de servicios ecosistémicos (Díaz & Cabido 1997, 2001; Tilman et al. 1997; Chapin III et al. 2000; Loreau et al. 2001), y en la capacidad de respuesta de los ecosistemas frente a factores de estrés o disturbio (Tilman & Downing 1994; Chapin III et al. 2000; McCann 2000). Los resultados del presente trabajo, al igual que resultados obtenidos en pastizales de otras regiones sub-húmedas de Argentina y del mundo, sugieren que el pastoreo moderado de grandes herbívoros favorecería el mantenimiento de la diversidad de especies vegetales nativas en los pastizales serranos del Sistema de Ventania. Sin embargo, el hecho que el muestreo de la vegetación no se repitió en el tiempo, y que las mediciones fueron realizadas al final de un año con precipitaciones por encima del promedio histórico, limita el alcance de las conclusiones en el presente estudio. Aún resta conocer si el patrón de diversidad descripto resulta sensible a la variabilidad interanual de las precipitaciones, en particular durante períodos de sequía, dado que la interacción entre ambos factores (pastoreo y sequía) puede afectar la composición botánica del pastizal (Heitschmidt et al. 2005). Así mismo, falta definir de manera precisa los valores absolutos de la intensidad de pastoreo en términos de la relación demanda:oferta de forraje que favorecen la diversidad florística. El control del pastoreo para promover diversidad debería ser uno de los objetivos primarios en la planificación del manejo de los pastizales serranos del Sistema de Ventania, tanto en su utilización para la producción ganadera como en su preservación en las Áreas Naturales Protegidas. En este último caso, el pastoreo controlado de grandes herbívoros exóticos podría suplir la ausencia actual de disturbios naturales (e.g., fuego, grandes herbívoros nativos) y fomentar la conservación de la biodiversidad, uno de los principales fines de la creación de las Áreas Naturales Protegidas.
AGRADECIMIENTOS
Al personal del Parque Provincial Ernesto Tornquist y de la Estancia Palo Alto por autorizarnos el ingreso a las áreas de muestreo. A dos revisores anónimos y a la editora por su cuidadosa revisión y cuyos comentarios aumentaron la calidad del manuscrito.
BIBLIOGRAFÍA
1. ALTESOR, A; G PIÑEIRO; F LEZAMA; RB JACKSON; M SARASOLA; ET AL. 2006. Ecosystem changes associated with grazing in subhumid South American grasslands. J. Veg. Sci., 17:323-332. [ Links ]
2. BURGOS, J. 1968. El clima de la provincia de Buenos Aires en relación con la vegetación natural y el suelo. Pp. 33-100 en: Cabrera, AL (ed.). Flora de la provincia de Buenos Aires. Colección Científica INTA, Buenos Aires. [ Links ]
3. CAPPANNINI, D; CO SCOPPA & J VARGAS GIL.1971. Suelos de las Sierras Australes de la Provincia de Buenos Aires. Pp. 203-234 en: Reunion Sobre Geología de las Sierras Australes Bonaerenses. Comisión Investigaciones Científicas de la Provincia de Buenos Aires. [ Links ]
4. CINGOLANI, AM; MR CABIDO; D RENISON & V SOLÍS NEFFA. 2003 Combined effects of environment and grazing on vegetation structure in Argentine granite grasslands. J. Veg. Sci., 14:223-232. [ Links ]
5. CINGOLANI, AM; I NOY-MEIR & S DÍAZ. 2005. Grazing effects on rangeland diversity: A synthesis of contemporary models. Ecol. Appl., 15:757-773. [ Links ]
6. CHANETON, EJ; SB PERELMAN; M OMACINI & RJC LEÓN. 2002. Grazing, Environmental Heterogeneity, and Alien Plant Invasions in Temperate Pampa Grasslands. Biol. Inv., 4:7-24. [ Links ]
7. CONNELL, JH. 1978. Diversity in tropical rain forests and coral reefs. High diversity of trees and corals is maintained only in a nonequilibrium state. Science, 199:1302-1310. [ Links ]
8. CHAPIN III, FS; ES ZAVALETA; VT EVINER; RL NAYLOR; PM VITOUSEK; ET AL. 2000. Consequences of changing biodiversity. Nature, 405:234-242. [ Links ]
9. DÍAZ S & M CABIDO. 1997. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci., 8:463-474. [ Links ]
10. DÍAZ S & M CABIDO. 2001. Vive la différence: Plant functional diversity matters to ecosystem processes. TREE, 16:646-655. [ Links ]
11. DÍAZ, S; S LAVOREL; S MCINTYRE; V FALCZUK; F CASANOVES; ET AL. 2007. Plant trait responses to grazing - A global synthesis. Glob. Chang. Biol., 13:313-341. [ Links ]
12. DISTEL, RA & RM BÓO. 1996. Vegetation states and transitions in temperate semiarid rangelands of Argentina. Pp. 117-118 en: Proceedings of the Fifth International Rangeland Congress. Salt Lake City, Utah, USA. [ Links ]
13. DUPRÉ, C & M DIEKMANN. 2001. Differences in species richness and life-history traits between grazed and abandoned grasslands in southern Sweden. Ecography, 24:275-286. [ Links ]
14. FAHNESTOCK, JT & JK DETLING. 2000. Morphological and physiological responses of perennial grasses to long-term grazing in the Pryor Mountains, Montana. Am. Midl. Nat., 143:312-320. [ Links ]
15. FRANGI, JL & OJ BOTTINO. 1995. Comunidades vegetales de la Sierra de la Ventana, Provincia de Buenos Aires, Argentina. Rev Fac Agron, La Plata, 71:93-133 [ Links ]
16. FRANGI, JL; NE SÁNCHEZ; MG RONCO; G ROVETTA & R VICARI. 1980. Dinámica de la biomasa y productividad primaria aérea neta de un pastizal de "flechillas" de sierra de La Ventana (Bs. As., Argentina). Bol. Soc. Arg. Bot., 19:203-228. [ Links ]
17. FRIDLEY, JD; JJ STACHOWICZ; S NAEEM; DF SAX; EW SEABLOOM; MD SMITH; TJ STOHLGREN; D TILMAN; B VON HOLLE. 2007. The invasion paradox: reconciling pattern and process in species invasions. Ecology, 88:3-17. [ Links ]
18. GRIME, JP. 2001. Plant strategies, vegetation processes, and ecosystem properties. 2nd Edition. John Wiley and Sons, LTD, Chichester, UK. [ Links ]
19. HARPER, JL. 1977. Population Biology of Plants. Academic Press, Londres, UK. [ Links ]
20. HEITSCHMIDT, RK; KD KLEMENT & MR HAFERKAMP. 2005. Interactive effects of drought and grazing on Northern Great Plain rangelands. Range. Ecol. Manage., 58:11-19. [ Links ]
21. HICKEY, WC. 1961. Growth Form of Crested Wheatgrass as Affected by Site and Grazing. Ecol., 42:173-176. [ Links ]
22. HOOPER, DU; FS CHAPIN III; JJ EWEL; A HECTOR; P INCHAUSTI; ET AL. 2005. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr., 75:3-35. [ Links ]
23. LAVOREL, S; S MCINTYRE & K GRIGULIS. 1999. Plant response to disturbance in a Mediterranean grassland: How many functional groups? J. Veg. Sci., 10:661-672. [ Links ]
24. LEGENDRE, P & L LEGENDRE. 1998. Numerical ecology. Elsevier Science BV, Amsterdam. [ Links ]
25. LONG, MA & CM GRASSINI. 1997. Actualización del conocimiento florístico del Parque Provincial Ernesto Tornquist. Informe final Convenio Ministerio de Asuntos Agrarios-Universidad Nacional del Sur. Bahía Blanca, Argentina. [ Links ]
26. LOREAU, M; S NAEEM; P INCHAUSTI; J BENGTSSON; JP GRIME; ET AL. 2001. Ecology: Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science, 294:804-808. [ Links ]
27. LOYDI, A; RA DISTEL & SM ZALBA. 2010. Large Herbivore Grazing and Non-Native Plant Invasions in Montane Grasslands of Central Argentina. Nat. Areas. J., 30:148-155. [ Links ]
28. MCCANN, KS. 2000. The diversity-stability. Nature, 405:228-233. [ Links ]
29. MILCHUNAS, DG; OE SALA & WK LAUENROTH. 1988. A generalized model of the effects of grazing by large herbivores on grassland community structure. Am. Nat., 132:87-106. [ Links ]
30. NAEEM, S; JMH KNOPS; D TILMAN; KM HOWE; T KENNEDY; ET AL. 2000. Plant diversity increases resistance to invasion in the absence of covarying extrinsic factors. Oikos, 91:97-108. [ Links ]
31. NAI-BREGAGLIO, M; E PUCHETA & M CABIDO. 2002. El efecto del pastoreo sobre la diversidad florística y estructural en pastizales de montaña del centro de Argentina. Rev. Chil. Hist. Natl., 75:613-623. [ Links ]
32. NOY-MEIR, I; M GUTMAN & Y KAPLAN. 1989. Responses of Mediterranean grassland plants to grazing and protection. J. Ecol., 77:290-310. [ Links ]
33. OLFF, H & ME RITCHIE. 1998. Effects of herbivores on grassland plant diversity. TREE, 13:261-265. [ Links ]
34. OSEM, Y; A PEREVOLOTSKY & J KIGEL. 2002. Grazing effect on diversity of annual plant communities in a semi-arid rangeland: Interactions with small-scale spatial and temporal variation in primary productivity. J. Ecol., 90:936-946. [ Links ]
35. PAZOS, GE; AJ BISIGATO & MB BERTILLER. 2007. Abundance and spatial patterning of coexisting perennial grasses in grazed shrublands of the Patagonian Monte. J. Arid. Environ., 70:316-328. [ Links ]
36. PECO, B; I DE PABLOS; J TRABA & C LEVASSOR. 2005. The effect of grazing abandonment on species composition and functional traits: The case of dehesa grasslands. Basic and Applied Ecol., 6: 175-183. [ Links ]
37. PUCHETA, E; M CABIDO; S DÍAZ & G FUNES. 1998. Floristic composition, biomass, and aboveground net plant production in grazed and protected sites in a mountain grassland of central Argentina. Acta Oecologica, 19:97-105. [ Links ]
38. PYKÄLÄ, J. 2005a. Plant species responses to cattle grazing in mesic semi-natural grassland. Agric, Ecosyst. & Environ., 108:109-117. [ Links ]
39. PYKÄLÄ, J. 2005b. Cattle grazing increases plant species richness of most species trait groups in mesic semi-natural grasslands. Plant Ecol., 175: 217-226. [ Links ]
40. RAMBO, JL & SH FAETH. 1999. Effect of vertebrate grazing on plant and insect community structure. Conserv. Biol., 13:1047-1054. [ Links ]
41. RICHARDSON, DM; N ALLSOPP; CM D'ANTONIO; SJ MILTON & M REJMANEK. 2000. Plant invasions - The role of mutualisms. Biol. Rev. Camb. Philos. Soc., 75:65-93. [ Links ]
42. ROOK, AJ; B DUMONT; J ISSELSTEIN; K OSORO; MF WALLIS DE VRIES; ET AL. 2004. Matching type of livestock to desired biodiversity outcomes in pastures - a review. Biol. Conserv., 119:137-150. [ Links ]
43. RÚGOLO DE AGRASAR, ZE; PE STEIBEL & HO TROIANI. 2005. Manual ilustrado de las gramíneas de la provincia de La Pampa. Universidad Nacional de La Pampa, Universidad Nacional de R. EdUNLPam. Pp. 374. [ Links ]
44. RUSCH, GM & M OESTERHELD. 1997. Relationship between productivity, and species and functional group diversity in grazed and non-grazed Pampas grassland. Oikos, 78:519-526. [ Links ]
45. SALA, OE; M OESTERHELD; RJC LEÓN & A SORIANO. 1986. Grazing effects upon plant community structure in subhumid grasslands of Argentina. Vegetatio, 67:27-32. [ Links ]
46. SCOROLLI, AL. 1999. Demografía y áreas de actividad de una población de caballos cimarrones en el Parque Provincial Ernesto Tornquist. Universidad Nacional del Sur, Bahía Blanca. Argentina. [ Links ]
47. STOCKTON, SA; S ALLOMBERT; AJ GASTON & JL MARTIN. 2005. A natural experiment on the effects of high deer densities on the native flora of coastal temperate rain forests. Biol. Conserv., 126:118-128. [ Links ]
48. SUTHERLAND, W. 1996. Ecological census techniques. A handbook. Cambridge University Press, Cambridge, UK. [ Links ]
49. TEAGUE, WR; SL DOWHOWER & JA WAGGONER. 2004. Drought and grazing patch dynamics under different grazing management. J. Arid Environ., 58:97-117. [ Links ]
50. TILMAN, D. 2000. Causes, consequences and ethics of biodiversity. Nature, 405:208-211. [ Links ]
51. TILMAN, D & JA DOWNING. 1994. Biodiversity and stability in grasslands. Nature, 367:363-5. [ Links ]
52. TILMAN, D; J KNOPS; D WEDIN; P REICH; M RITCHIE; ET AL. 1997. The influence of functional diversity and composition on ecosystem processes. Science, 277:1300-1302. [ Links ]
53. TODD, SW & MT HOFFMAN. 1999. A fence-line contrast reveals effects of heavy grazing on plant diversity and community composition in Namaqualand, South Africa. Plant Ecol., 142:169-178. [ Links ]
54. VAVRA, M; CG PARKS & MJ WISDOM. 2007. Biodiversity, exotic plant species, and herbivory: The good, the bad, and the ungulate. For. Ecol. and Manag., 246:66-72. [ Links ]
55. WANG, Y; M SHIYOMI; M TSUIKI; M TSUTSUMI; X YU; ET AL. 2002. Spatial heterogeneity of vegetation under different grazing intensities in the Northwest Heilongjiang Steppe of China. Agric, Ecosyst. & Environ., 90:217-229. [ Links ]
56. WEST, NE. 1993. Biodiversity of rangelands. J. Range Manag., 46:2-13. [ Links ]
57. WESTOBY, M; B WALKER & I NOY-MEIR. 1989. Opportunistic management for rangelands not at equilibrium. J. Range Manag., 42:266-274. [ Links ]
58. ZALBA, SM & NC COZZANI. 2004. The impact of feral horses on grassland bird communities in Argentina. Anim. Conserv., 7:35-44. [ Links ]
59. ZAR, JH. 1999. Biostatistical Analisis. Prentice Hall, Upper Saddle River, New Jersey. USA. [ Links ]
60. ZULOAGA, FO & O MORRONE. 2007. Catálogo de plantas vasculares del cono sur. Instituto de Botánica Darwinion. www.darwin.edu.ar/Proyectos/FloraArgentina/FA.asp. [ Links ]

