










Serviços Personalizados
Journal

Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkEcología austral
versão On-line ISSN 1667-782X
Ecol. austral vol.22 no.1 Córdoba abr. 2012
ARTÍCULO ORIGINAL
Caracterización regional de la estructura y de la productividad de la vegetación de la Puna mediante el uso de imágenes MODISCaracterización regional de la estructura y de la productividad de la vegetación de la Puna mediante el uso de imágenes MODIS
Pablo Baldassini1,2,*; José N Volante4; Laura M Califano5 & José M Paruelo1,2,3
1. Laboratorio de Análisis Regional y Teledetección
2. Facultad de Agronomía. Universidad de Buenos Aires. IFEVA. CONICET
3. Departamento de Métodos Cuantitativos y Sistemas de Información. Facultad de Agronomía. UBA
4. INTA EEA Salta;
5. INTA OIT Humahuaca.
* Facultad de Agronomía (UBA). Laboratorio de Análisis Regional y Teledetección. IFEVA. CONICET. Av. San Martín 4453 (C1417DSE) Ciudad Autónoma de Buenos Aires. Argentina. Email: pbaldass@agro.uba.ar
Recibido: 29 de agosto de 2011;
Fin de arbitraje: 20 de octubre de 2011;
Revisión recibida: 22 de noviembre de 2011;
Aceptado: 29 de noviembre de 2011
RESUMEN. La Puna es una región semiárida ubicada por encima de los 3300 m.s.n.m. en el noroeste argentino. Si bien existen descripciones florísticas parciales de la vegetación en el área, no existen trabajos que describan su fisonomía y su funcionamiento, aspectos claves para el manejo de la vegetación de la región. El objetivo de este trabajo fue cartografiar los diferentes tipos fisonómicos presentes y caracterizar su funcionamiento a partir de la estimación de la productividad primaria neta aérea (PPNA) en un área de 69210 km2 en las provincias de Salta y Jujuy. La aproximación metodológica combinó la utilización de datos provistos por sensores remotos, modelos biofísicos para estimar la productividad y censos fisonómico-florísticos. De las clases vegetadas, las estepas arbustivas (Estepa arbustiva densa y Estepa arbustiva rala) fueron las que ocuparon mayor proporción del territorio analizado, cubriendo entre ambas el 45% aproximadamente, mientras que las Estepas graminosas densas fueron las menos representadas, cubriendo poco más del 1%. Entre las clases no vegetadas el Suelo desnudo fue la clase más extendida, cubriendo aproximadamente 30% del área, seguida por la clase Salar, que ocupó 8.15%. El tipo fisonómico más productivo fue la Estepa graminosa densa (PPNA = 3900 kg MS.ha-1.año-1), mientras que la Estepa arbustiva rala presentó la menor productividad (PPNA = 472 kg MS.ha-1.año-1). Los restantes tipos fisonómicos no superaron en promedio los 1000 kg MS.ha-1.año-1.
Palabras clave: Índices espectrales; Sensores remotos; Recursos forrajeros; Funcionamiento ecosistémico.
ABSTRACT. Regional characterization of the structure and productivity of the vegetation of the Puna using MODIS images: The Puna is a semi-arid region located over 3300 m.a.s.l. in the northwestern portion of Argentina. Previous regional studies in Puna focused on the description of the floristic heterogeneity of relatively small portions of the territory. No integrative descriptions of the phisiognomy and functioning of the Puna ecosystems, two keys aspects for development of management actions, have been produced. In this article we generated a map of physonogmic types and we characterized above-ground net primary production (ANPP) of the different classes defined over an area of 69210 km2. The methodology to characterize vegetation structure (physiognomic units) combines remotely sensed data from MODIS sensor with field observations. Further, we applied biophysical models and satellite information to estimate ANPP. Among the classes with more than 5% of plant cover, shrub steppes (both sparse and dense) occuppied most of the area (45%). Dense grass steppes covered less than 1%. Bare soil covered most that 30% of the area, followed by Salt flats (8.15%). The most productive physiognomic type was the dense grass steppe (ANPP = 3900 kg DM.ha-1.year-1), while the sparse shrub steppe unit presented the lowest values (ANPP = 472 kg DM.ha-1.year-1). The remaining physiogmomic types presented ANPP values lower than 1000 kg DM.ha-1.year-1.
Keywords: Ecoregions; Spectral indixes; Remote sensing; Forage resources; Ecosystem functioning.
INTRODUCCIÓN
El desajuste en el tiempo y en el espacio entre la oferta y la demanda de forraje por parte del ganado, es uno de los factores determinantes en el proceso de deterioro de sistemas áridos y semiáridos (Paruelo & Aguiar 2003). La ausencia de una adecuada caracterización de la oferta forrajera en términos de calidad y cantidad es una seria limitación en el diseño de sistemas de manejo pastoril en zonas áridas (Golluscio et al. 1998, 2010). El pastoreo por llamas, ovejas y cabras es la actividad económica principal de la región de la Puna (Ruthsatz & Movia 1975; Cajal 1998a; Fernández & Busso 1999), una altiplanicie extensa ubicada en el centro-oeste del continente americano que ocupa parte de Argentina, Bolivia, Chile y Perú (Cabrera 1957). La escasa regulación de esta actividad ha ocasionado un impacto adverso en los ecosistemas naturales, como la pérdida de cobertura vegetal, la desaparición de especies valiosas y la erosión del suelo (Braun Wilke 1991; Fernández & Busso 1999; Reboratti 2006). A su vez, se ha observado un impacto negativo del sobrepastoreo sobre la diversidad de artrópodos y roedores, como así también sobre la nidificación de especies de aves a orillas de las principales lagunas (Cajal 1998a; Cajal & Bonaventura 1998). En este contexto, la caracterización de la heterogeneidad de los recursos forrajeros y la cuantificación y monitoreo de la productividad primaria, constituyen un paso insoslayable en el diseño de sistemas de manejo pastoril que permitan minimizar los impactos negativos del pastoreo sobre la estructura y el funcionamiento ecosistémico y sostener o incrementar la producción ganadera (Paruelo & Golluscio 1994; Oesterheld et al. 2005; Grigera et al. 2007a).
Los estudios previos en la región de la Puna se han focalizado en la descripción de la heterogeneidad florística de diferentes áreas de la región. Estos trabajos se limitaron a la descripción de la flora a nivel de especies (Cabrera 1957, 1976, 1977, 1978, 1983; Kiesling et al. 1983; Nadir & Chafatinos 1990; Novara 2003) o, cuando consideraron niveles de organización superiores, se restringieron sólo a ciertas porciones de la región (Ruthsatz & Movia 1975; Castañera & Gonzalez 1991; Bonaventura et al. 1998; Tálamo et al. 2010). Por su parte, el conocimiento en términos del potencial forrajero de la Puna argentina es escaso y la mayor parte de los estudios se han realizado principalmente en otros países andinos.
Los sensores remotos y las técnicas de procesamiento digital de imágenes facilitan la caracterización estructural de la vegetación ya que permiten obtener mapas mediante diferentes métodos de clasificación (Congalton et al. 2002; Chuvieco 2002). Las descripciones espaciales de aspectos estructurales de la vegetación, tales como la heterogeneidad fisonómico-florística, tienen importancia tanto desde el punto de la conservación como del aprovechamiento pastoril de los sistemas áridos (Paruelo et al. 2004). Desde el punto de vista de la producción ganadera, la caracterización fisonómico florística permite definir diferencias regionales en la calidad de los recursos forrajeros, ya que la proporción de tipos funcionales de plantas (TFP) (Paruelo et al. 2004) leñosos es un control del índice de cosecha de forraje (Golluscio et al. 1998). La teledetección también permite estudiar atributos funcionales de la vegetación, tales como la dinámica estacional de la productividad primaria neta aérea (PPNA) o la evapotranspiración (Petorelli et al. 2005; Paruelo 2008). Las caracterizaciones funcionales de los ecosistemas basadas sobre datos espectrales han sido objeto de estudio en distintos sistemas de Argentina (Paruelo et al. 2000; Posse & Cingolani 2000; Garbulsky & Paruelo 2004; Paruelo et al. 2004; Guerschman & Paruelo 2005; Nosetto et al. 2005; Zoffoli et al. 2008). La PPNA se define como la tasa de acumulación de biomasa vegetal por unidad de superficie y tiempo en órganos aéreos (Sala & Austin 2000), representa la principal fuente de energía para los herbívoros y es un determinante clave de la carga animal (Oesterheld et al. 1998; Golluscio et al. 2009). El modelo ecofisiológico propuesto por Monteith (1972) permite cuantificar la producción de forraje a partir de la radiación fotosintéticamente activa incidente (RFA), la fracción de esa radiación que es interceptada por los tejidos verdes (fRFA), y la eficiencia en el uso de la radiación (EUR):
PPNA = fRFA * RFA (MJ.m-2.t-1) * EUR (g MS/MJ)
La fRFA puede ser estimada a partir de distintos índices de vegetación, como el IVN (Índice de Vegetación Normalizado) o el IVM (Índice de Vegetación Mejorado) (Paruelo 2008). Este último es más apto para estudios de paisaje con canopeos abiertos como los sistemas semiárido (Huete et al. 2002). Incluye la reflectancia en la porción del azul del espectro electromagnético (459-479 nm) y coeficientes de corrección que permiten minimizar el efecto de ciertos contaminantes de la atmósfera, y desacoplar la señal del sustrato y la vegetación, pemitiendo incrementar la señal de ésta. Se expresa como:
IVM = 2.5 * (IR-R) / (IR+6 * R-7.5 * A+1)
Donde A, R e IR corresponden a las reflectancias corregidas atmosféricamente en la porción del azul (459-479 nm), del rojo (620-670 nm) y del infrarrojo cercano (841-876 nm) del espectro electromagnético, respectivamente.
La RFA es una porción del espectro electromagnético (400 a 700 nm) que está directamente relacionada con la producción fotosintética de las plantas y representa ~48% de la radiación global incidente (McCree 1972; Blackburn & Proctor 1983). La EUR difiere entre especies y comunidades vegetales y varían ante modificaciones en las condiciones ambientales, como la disponibilidad de agua, la temperatura o la nutrición (Potter et al. 1993; Sinclair & Muchow 1999; Nouvellon et al. 2000; Piñeiro et al. 2006). Sin embargo, en diversos estudios se la considera constante en el tiempo y variable sólo según el tipo de cobertura, ya que la cantidad de radiación absorbida por la vegetación (RFAA) presenta una variación considerablemente mayor y es estimable con mayor precisión (Grigera et al. 2007a).
El objetivo general de este trabajo fue caracterizar la heterogeneidad estructural y funcional de los ecosistemas puneños de las provincias de Salta y Jujuy, con particular énfasis en aquellos aspectos que definen la receptividad forrajera. Especificamente se buscó: 1) definir y cartografiar los tipos fisonómicos de vegetación de la Puna en base a la dinámica estacional de los índices espectrales provistos por imágenes MODIS y a relevamientos a campo, y 2) describir la variabilidad espacial y temporal de la PPNA estimada mediante la utilización del modelo de Monteith (1972) y de información provista por imágenes MODIS.
MATERIALES Y MÉTODOS
Área de estudio
La zona de estudio se encuentra ubicada al noroeste de Argentina; se extiende por la mitad occidental de la provincia de Jujuy y por el oeste de Salta y abarca un área de ~69210 km2. Desde el punto de vista fitogeográfico, comprende dos provincias del Dominio Andino: la Provincia Puneña, ubicada entre 3300 y 4400 m.s.n.m., y la Provincia Altoandina, que se encuentra por encima de los 4400 m (Cabrera 1976). El clima de la región es seco y frío, con una gran variabilidad espacial y temporal de la precipitación. En la Provincia Altoandina, la temperatura media anual es de 3.1 °C, mientras que las precipitaciones ocurren en forma de nieve o granizo en cualquier estación (Cabrera 1957). En la Provincia Puneña, la temperatura media es inferior a los 10 °C y las lluvias son casi exclusivamente estivales y presentan una gran variabilidad temporal y espacial (Cabrera 1957; Cajal 1998b). En el noreste, las precipitaciones anuales son cercanas a los 350 mm, mientras que en el sudoeste son inferiores a 50 mm/año.
Bases de datos
Para la caracterización estructural y funcional de la vegetación se utilizaron datos de IVM del producto MOD13Q1 provenientes del sensor MODIS-Terra. Estas imágenes poseen una resolución espacial de 231 m y temporal de 16 días, lo que resulta adecuado para el monitoreo estacional e interanual de la vegetación terrestre (Justice et al. 1998; Huete et al. 2002). Se utilizaron las escenas h11v11 y h12v11 del período comprendido entre los años 2000 y 2009. Los datos de radiación incidente fueron obtenidos de una base de datos pública. Se contó con datos diarios entre los años 1994 y 2004 y se utilizó el promedio de los datos disponibles para los años comprendidos entre el 2005 y el 2009. Por otro lado, se usó un Modelo Digital de Elevación (DEM, por sus siglas en inglés) con una resolución espacial de 90 m. A partir de este modelo se obtuvieron datos de pendiente, a partir de los cuales se discriminaron ciertas coberturas no vegetadas con similar comportamiento espectral.
Caracterización estructural de la vegetación
Identificación de tipos fisonómicos de vegetación. Las categorías fisonómicas utilizadas para caracterizar la vegetación se definieron siguiendo criterios jerárquicos preestablecidos relativos a la forma de vida (leñosas -árboles y arbustos-, herbáceas, musgos y líquenes), a la estructura vertical y al grado de cobertura, en base al Sistema LCCS (Land Cover Classification System) propuesto por FAO (Di Gregorio & Jansen 2000) y a las categorías establecidas por Paruelo et al. (2004). Se identificaron cinco clases vegetadas y cuatro no vegetadas:
1. Estepa arbustiva densa (Ead): áreas dominadas por arbustos, con una cobertura abierta (entre 15 y 65%) o cerrada (superior a 65%). Puede observarse la presencia de un segundo estrato compuesto por herbáceas en cobertura dispersa (inferior a 15%).
2. Estepa graminosa arbustiva (Ega): áreas dominadas por especies graminiformes, con una cobertura abierta (entre 15 y 65%) o cerrada (superior a 65%). Se observa la presencia de arbustos con cobertura dispersa (inferior a 15%).
3. Estepa arbustiva rala (Ear): áreas de baja cobertura dominadas por arbustos, con una cobertura abierta (entre 15 y 65%) o dispersa (inferior a 15%). Puede observarse la presencia de un segundo estrato compuesto por herbáceas en cobertura dispersa (inferior a 15%).
4. Estepa graminosa densa (Egd): áreas dominadas por especies graminiformes, con coberturas cerradas (superior a 65%) en donde no comparten el espacio con otras formas de vida.
5. Estepa graminosa rala (Egr): áreas dominadas por especies graminiformes, con coberturas abiertas (entre 15% y 65%), en donde no comparten el espacio con otras formas de vida.
6. Áreas no vegetadas: salares (Sl), suelo desnudo (Sd), cuerpos de agua (Ag) y nieve (Ni) permanentes. En esta categoría se incluyen todos aquellos tipos de cubiertas que posean una cobertura vegetal inferior a 5%.
Recolección de datos de la estructura de la vegetación. La información sobre la estructura de la vegetación fue recolectada entre los meses de enero y agosto de 2008 en tres campañas. Las áreas relevadas se acotaron a zonas homogéneas y representativas de la vegetación bajo estudio y se limitaron a regiones próximas a caminos vehiculares. Se realizaron 380 censos a campo a escala de stand, en los cuales se relevó la cobertura total de herbáceas y de arbustos. En base a esta información cada uno de los censos fue asignado a una de las clases vegetadas. Todos los censos fueron georreferenciados utilizando sistemas de posicionamiento global (GPS) y se categorizaron según los tipos fisonómicos definidos a priori. A su vez, se identificaron nuevas áreas correspondientes a las clases Sl, Sd y Ag mediante la fotointerpretación de imágenes disponibles en Google Earth®. Estas zonas no se computaron dentro del número inicial de censos realizados.
Mapa fisonómico de vegetación. Clasificación y evaluación. Se realizó una clasificación supervisada empleando el algoritmo de Máxima Probabilidad como regla de decisión, sobre una serie temporal de 23 imágenes de IVM de las escenas h11v11 y h12v11 correspondientes a 2008. Las áreas de entrenamiento comprendieron cerca de la mitad de los puntos de GPS tomados a campo (189 observaciones) y las áreas delimitadas en Google Earth®. Se utilizaron los píxeles correspondiente a cada punto GPS como así también los píxeles vecinos que se ubicaban dentro de un área buffer de 300 m. A la clasificación obtenida se le aplicó un filtro mayoritario, reemplazando el valor de cada píxel por el valor modal registrado sobre una ventana de 3x3 píxeles. La evaluación de la clasificación se realizó utilizando el conjunto de observaciones de campo que no fueron usados en el proceso de clasificación (191). Estos puntos también se categorizaron según los tipos fisonómicos definidos a priori y se compararon con el resultado de la clasificación realizada. La exactitud de la clasificación se evaluó utilizando diferentes índices derivados de la matriz de contingencia, como la fiabilidad global, la fiabilidad del productor y la fiabilidad del usuario.
Caracterización funcional de la vegetación. Estimación de la PPNA. La PPNA se calculó utilizando el modelo de Monteith (1972). La fRFA se estimó como una función lineal del IVM (Grigera & Oesterheld 2006) para lo cual se parametrizó la relación asignando una absorción nula (fRFA = 0) al valor de IVM correspondiente a píxeles sin cobertura vegetal (suelo desnudo o peladares) y se fijó un valor máximo (fRFA=0.95) al percentil 98 de los valores de IVM de píxeles con alta cobertura vegetal, siendo los valores superiores igualados a 0.95 (Paruelo 2008). Estos píxeles se ubicaron en la zona de las Yungas, contigua geográficamente a la Puna. Se aplicó un filtro de calidad a las imágenes de IVM con el fin de descartar aquellos píxeles que contaran con la presencia de nubes, sombras, y/o aerosoles en la atmósfera. Para ello se utilizó la banda QA del producto MOD13Q1, reemplazando los valores de IVM de los píxeles de mala calidad por el valor promedio de las imágenes correspondientes a las fechas inmediatamente anterior y posterior. El valor de RFA acumulada durante los 16 días correspondientes a cada imagen MODIS, se calculó a partir de la radiación incidente diaria, considerada el 48% de la misma (McCree 1972; Blackburn & Proctor 1983). Debido a que en la región de la Puna no se cuenta con estimaciones de EUR, se utilizaron valores registrados para tipos de vegetación similares en Patagonia, dada la similitud entre ambos ecosistemas (Cabrera 1976; Fernández & Busso 1999). Irisarri et al. (2011) determinaron un valor de EUR de 0.7 g MS/MJ para los mallines de la Patagonia, el cual se utilizó para calcular la PPNA de los pastizales cerrados de la Puna. Para los arbustales y pastizales abiertos de la Puna, se utilizó una EUR de 0.3 g MS/MJ, obtenido a partir datos de IVN de imágenes Landsat del mes de diciembre y de datos de cortes de biomasa realizados a campo (Paruelo et al. 2004).. Estas operaciones se realizaron en ambiente ENVI 4.0 e IDL 6.0 (ITT).
RESULTADOS
Identificación de tipos fisonómicos
La clasificación de coberturas obtenida (Figura 1) tuvo una fiabilidad global de 85%, donde todas las categorías identificadas tuvieron una exactitud del productor superior a 75%, con excepción de la categoría Egr, cuya precisión fue 67.42% (Tabla 1). En lo que respecta a la exactitud del usuario, todas las clases, con excepción de Sd y Ear, superaron el 80% (Tabla 1). La clase Ni fue identificada a posteriori de la clasificación. Dado el similar comportamiento espectral entre Ni y Sl, la separación entre ambas categorías se realizó mediante la utilización de una imagen de pendiente, derivada de un Modelo Digital de Elevación. Aquellos píxeles que fueron clasificados inicialmente como Sl y que poseían una pendiente superior a 5% fueron reclasificados en la categoría Ni. De las clases vegetadas, las estepas arbustivas (Ead y Ear) fueron las que ocuparon mayor proporción del territorio analizado; entre ambas cubrieron ~45%. Mientras tanto, Egd fue la menos representativa al cubrir poco más del 1%. Las clases Egr y Ega cubrieron 5% y 6.75% de la zona de estudio, respectivamente. En cuanto a las clases no vegetadas, Sd fue la clase más extendida ya que cubrió aproximadamente 30% del área, seguida por la clase Sl, que ocupó 8.15%. Las clases Ag y Ni fueron las menos representativas dentro de las clases no vegetadas, cubriendo 0.17% y 2.05%, respectivamente. La proporción de clases vegetadas de baja cobertura, como Ear, y las clases no vegetadas (Sd y Sl, principalmente) incrementaron su proporción hacia el sudoeste de la zona de estudio, conforme disminuyen las precipitaciones anuales (Figura 1). Las Egd se ubicaron principalmente en la región húmeda, donde se encuentra más de 75% de las mismas, al igual que las Ega, cuya proporción asciende al 85%.
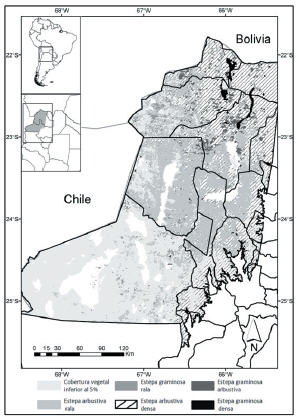
Figura 1. Clasificación de los tipos fisonómicos de vegetación identificados en el área de estudio. El mapa de unidades fisonómicas se realizó a partir de una clasificación supervisada de un compilado de imágenes MODIS de IVM del año 2008 (23 en total). En el mapa se observan las cinco clases vegetadas: y las clases que poseen una cobertura vegetal inferior al 5%.
Figure 1. Classification of the physognomic types identified in the study area. The map was generated from a supervised classification of a IVM images stack in the year 2008 (23 in total). The map shows five classes vegetated and classes with less than 5% of plant cover.
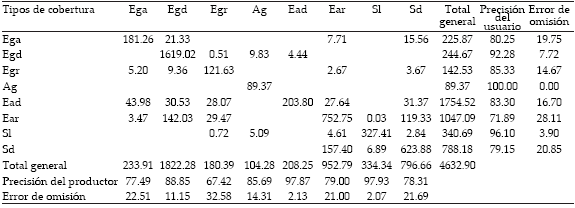
Tabla 1. Matriz de contingencia de la clasificación. Las columnas indican las clases de referencia, mientras que las filas muestran las categorías derivadas de la clasificación. La diagonal de esta matriz expresa el acuerdo entre las dos fuentes (mapa y datos de campo), mientras que los marginales suponen errores de asignación. En los extremos se detallan las estadísticas que permiten evaluar la precisión y confiabilidad de la clasificación: exactitud del productor, error de comisión, exactitud del usuario y error de comisión. En la validación se utilizaron tanto los píxeles correspondientes a cada punto de GPS (191) como así también los píxeles vecinos ubicados dentro de un área buffer de 300 m. Los valores observados en el cuerpo de la tabla corresponden al área (ha) de las distintas clases incluidas en las área buffer.
Table 1. Contingency matrix of the classification. The columns indicate the reference classes, while the rows showing the categories drawn from the classification. The diagonal of this matrix represents the agreement between the two sources (map and field data), while the marginal corresponds to allocation errors. At the the right column and the bottom row statistics to evaluate the accuracy and reliability of classification are presented: producer's accuracy, error of commission, user's accuracy and error of commission. In the validation were used the pixels for each GPS point (191) and the pixels located within a buffer area of 300 m. The values observed in the body of the table, correspond to the area (ha) of the different classes included in the buffer area.
Estimación de la PPNA
Se observó una reducción de la PPNA en el sentido este-oeste y norte-sur. Los departamentos ubicados al este de la Puna jujeña son los que registraron los valores mayores (Figura 2). El tipo fisonómico más productivo fue Egd (PPNA promedio de 3900 kg MS.ha-1.año-1), mientras que Ear presentó la menor productividad (PPNA promedio de 472 kg MS.ha-1.año-1) (Figura 3). Los restantes tipos fisonómicos no superaron, en promedio, los 1000 kg MS.ha-1.año-1; la PPNA promedio de las Ega, Egr y Ead fue de 680, 630 y 825 kg MS.ha-1.año-1, respectivamente (Figura 3). Todos los tipos fisonómicos presentaron una variabilidad espacial considerable mostrando Egd los valores mayores (Figuras 3). El valor promedio anual de PPNA máximo registrado para este tipo fisonómico fue 10464 kg MS.ha-1.año-1, mientras que el mínimo fue 1400 kg MS.ha-1.año-1 (Figura 3). La Ega tuvo un valor promedio máximo de PPNA anual de 1142 kg MS.ha-1.año-1, mientras que el mínimo fue 389 kg MS.ha-1.año-1 (Figura 3). La Egr y la Ead registraron un máximo anual de PPNA de 1074 y 1350 kg MS.ha-1.año-1, respectivamente, mientras que el mínimo anual fue de 243 y 372 kg MS.ha-1.año-1 (Figura 3). La Ear fue el único tipo fisonómico cuya PPNA anual máxima no superó los 1000 kg MS.ha-1.año-1, registrándose un valor de 750 kg MS.ha-1.año-1 (Figura 3). La PPNA anual mínima ha sido también la marca más baja entre todos los tipos fisonómicos, siendo apenas de 110 kg MS.ha-1.año-1 (Figura 3).

Figura 2. Mapa de productividad primaria neta aérea (PPNA) anual promedio en kg MS.ha-1.año-1. Los mismos derivan de la utilización de imágenes de IVM proveniente del sensor MODIS para el período 2000-2009 y de la aplicación del modelo propuesto por Monteith (1972).
Figure 2. Map of average annual aboveground net primary productivity (ANPP) (kg DM.ha-1.year-1). derived from IVM data from the MODIS sensor for the period 2000-2009 and the Monteith´s model (1972).
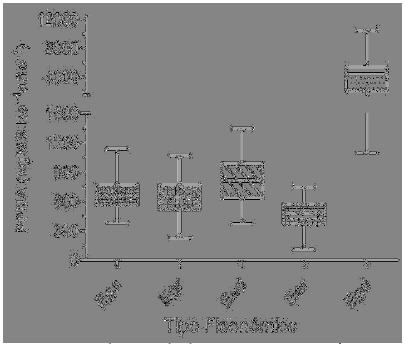
Figura 3. Productividad primaria neta aérea (PPNA) promedio anual para los diferentes tipos fisonómicos de vegetación identificados. Las cajas representan el rango intercuartílico, las barras indican los valores máximos y mínimos, y la línea central el valor promedio anual.
Figure 3. Annual average aboveground net primary productivity (ANPP) for the different vegetation types identified. The boxes represent the interquartile range , the . bars indicate maximum and minimum values and the central line corresponds to the average value.
La PPNA mostró un patrón estacional claro. Los valores mensuales máximos se registraron principalmente durante el mes de marzo para todos los tipos fisonómicos, con excepción de las Egr, cuyo valor máximo correspondió a enero (Figura 4). Los mínimos ocurrieron en el mes de junio para todos los tipos fisonómicos (Figura 4). Las Egd presentaron la mayor variabilidad relativa entre los máximos y mínimos mensuales, que en promedio fue 10% (Figura 4). Por el contrario, las Egr presentaron la mínima variabilidad relativa (4.8%), mientras que las demás unidades fisonómicas mostraron valores intermedios (Figura 4). La variabilidad interanual de la PPNA mensual (medida como el coeficiente de variación: STD/Media) fue máxima durante el período invernal para todos los tipos fisonómicos, siendo las Egd las que registraron los valores más altos (Figura 5). Esta unidad fisonómica presentó un coeficiente de variación de la PPNA entre años superior a 25% durante el primer mes de la estación de crecimiento. A diferencia de los demás tipos fisonómicos, las Ear registraron una mayor variabilidad mensual interanual durante agosto (Figura 5). Las Egr y las Egd mostraron los valores de CV mensual interanual más bajos (Figura 5).
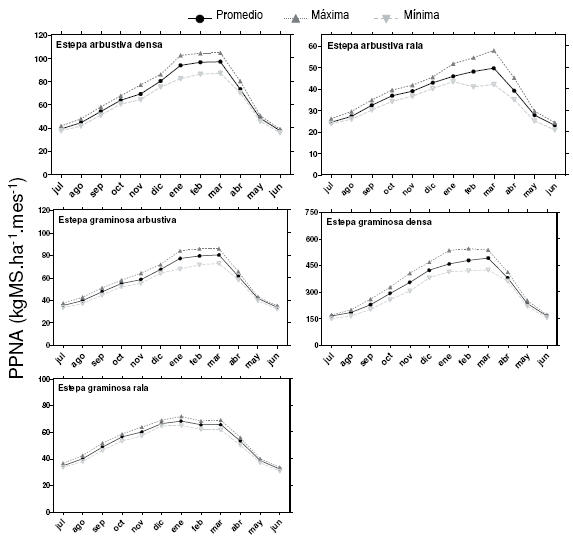
Figura 4. Productividad primaria neta aérea (PPNA) mensual para los diferentes tipos fisonómicos de vegetación identificados. Para los meses comprendidos entre los años 2000 y 2009 se detalla la productividad promedio mensual y las máximas y mínimas.
Figure 4. Aboveground net primary productivity (ANPP) monthly for the different vegetation types identified. Productivity for the period 2000-2009: the monthly average productivity, and highest and minimum.

Figura 5. Dinámica estacional del coeficiente de variación interanual (2000-2009) de la PPNA para los diferentes tipos fisonómicos identificados.
Figure 5. Seasonal dynamics of the interannual coefficient of variation (2000-2009) of the ANPP for the different physiognomic types identified.
DISCUSIÓN
Las descripciones fisiográficas de la Puna argentina que se encuentran en la literatura pertenecen a la parte norte de esta región, típicamente más húmeda que la del sur (Cabrera 1957; Ruthsatz & Movia 1975; Castañera & Gonzalez 1991; Bonaventura et al. 1998). Ruthsatz & Movia (1975) diferenciaron unidades de vegetación en la región en las estepas andinas del noreste de la provincia de Jujuy, las que cubren ~20% del área abordada en este trabajo. Otros trabajos [e.g., Castañera & Gonzalez (1991) y Bonaventura et al. (1998)] se restringieron al estudio de parte de la cuenca de Pozuelos. Este trabajo, por el contrario, abarcó una superficie sensiblemente mayor que los análisis previos. Tal como fuera descripto por autores como Cabrera (1957), Ruthsatz & Movia (1975), Cabrera (1976), Bonaventura et al. (1998) y Borgnia et al. (2006), la vegetación observada en el área de estudio presentó una estructura simple, con uno o dos estratos y con características propias de ambientes xéricos. Las estepas y las clases no vegetadas fueron las más abundantes, cubriendo los ambientes de vega (Egd) poco más del 1% del área. La clasificación de tipos fisonómicos obtenida a partir de la dinámica temporal del IVM, presentó una exactitud global similar a la observada en otras clasificaciones basadas en el comportamiento espectral diferencial de la vegetación (Guerschman et al. 2003; Cingolani et al. 2004; Paruelo et al. 2004; Borgnia et al. 2006). Si bien la distinción entre algunos tipos fisonómicos no fue tan clara, las categorías definidas presentaron una aceptable exactitud. Esto puede deberse a que las unidades fisonómicas descriptas no presentan discontinuidades, sino que por el contrario la diferenciación resulta de un cambio gradual en la cobertura o en la proporción de tipos funcionales de plantas.
Los valores de PPNA calculados constituyen una de las pocas estimaciones realizadas para la región de la Puna. Estas estimaciones se basaron sobre el modelo de Monteith (1972) y tomaron como información de base la proveniente de sensores remotos de resolución espacial moderada. Este modelo tiene una base biofísica sólida (Running et al. 2000; Paruelo 2008) y ha sido utilizado con éxito en muchos sistemas de pastizal y arbustal como los de la Puna (e.g., Paruelo et al. 1997; Piñeiro et al. 2006). A pesar de la robustez del método, el uso de estimaciones de PPNA basadas en sensores remotos requiere de una campaña de registro de datos a campo para, especialmente, derivar estimaciones de la eficiencia en el uso de la radiación (EUR) de las distintas unidades de vegetación (Piñeiro et al. 2006). Las estimaciones obtenidas no pudieron ser comparadas con datos de campo ya que no se han encontrado trabajos sobre la PPNA en la región estudiada.
Los valores y rangos de variabilidad de la PPNA registrada fueron mayores para los ambientes de vega (Egd) y menores en las estepas. La productividad mínima y media anual registrada en las vegas fue equivalente a la observada por Buono et al. (2010) para ciertos mallines patagónicos, aunque se observan diferencias en cuanto a los máximos anuales alcanzados; las Estepas graminosas densas (Egd) obtuvieron los valores más elevados. Sin embargo, la productividad media anual difirió considerablemente con las estimaciones de PPNA realizadas por Irisarri et al. (2011) en los mallines patagónicos, las cuales mostraron valores promedio de 6600 kg MS.ha-1.año-1, un 60.5% superior al obtenido en este y otros trabajos para los ambientes de vega (Paruelo et al. 2004; Buono et al. 2010).
Las diferencias observadas en la PPNA de los distintos tipos fisonómicos puede ser explicada por la influencia de diversos factores ambientales en donde la disponibilidad de agua es el factor de mayor importancia (Noy Meir 1973). Las bajas precipitaciones registradas en la región determinan que la disponibilidad de agua sea la principal limitante de la producción de las estepas arbustivas. Por el contrario, la extensión de la estación de crecimiento debido a las restricciones térmicas actúan como el factor limitante del crecimiento de las plantas en los pastizales húmedos o vegas (Egd) dado que estos ambientes poseen fuentes de agua adicionales a las lluvias, como vertientes naturales, agua de deshielo y arroyos que permiten mantener un nivel de humedad permanente durante todo el año (Borgnia, datos no publicados). Las Egd (vegas) mostraron los mayores niveles de variabilidad, tanto espacial como temporalmente, en comparación con los demás tipos fisonómicos. La mayor variabilidad espacial de la PPNA puede deberse a que difieren en su composición y diversidad florística (Borgnia et al. 2006), en función del régimen hídrico, la altitud, el sobrepastoreo y el manejo ganadero, la fisiografía, la textura del suelo, el pH y la salinidad (Borgnia, datos no publicados). En lo que respecta a la variabilidad mensual interanual de la PPNA, la misma ha presentado los máximos valores de coeficiente de variación durante los meses de invierno para todos los tipos fisonómicos, al igual que lo reportado por Paruelo et al. (2004) para estepas y mallines patagónicos.
La caracterización de la PPNA de los distintos tipos fisonómicos presentes mediante el uso de sensores remotos permite avanzar hacia un sistema de monitoreo espacialmente explícito, convirtiéndose en una herramienta de apoyo para la toma de decisiones en el manejo de los recursos naturales. Un conocimiento detallado de la variabilidad espacial y temporal de la productividad permite desarrollar diversas aplicaciones de manejo, como el balance y presupuesto forrajero, la determinación del tiempo de ocupación del ganado o la evaluación del manejo sobre la productividad forrajera (e.g., Paruelo et al. 1999), favoreciendo un uso más eficiente de los recursos y una reducción del riesgo de sobrepastoreo (Grigera et al. 2007b).
AGRADECIMIENTOS
Agradecemos la contribución de H. Dieguez, G. García Accinelli, L. Lizarraga, C. Morales, M. Zelaya y personal de INTA Abra Pampa e INTA Cerrillos. Este trabajo fue realizado gracias al aporte del LART, FAUBA, CONICET e INTA. Agradecemos el apoyo de la UBA (Programa UBACYT 2008-2010), del Fondo para la Investigación Científica y Tecnológica (FONCYT) (PICT 2199) y del Instituto Interamericano para la Investigación del Cambio Global (IAI, CRN 2031), financiado por National Science Foundation US (Grant GEO-0452325).
BIBLIOGRAFÍA
1. BLACKBURN, WJ & JTA PROCTOR. 1983. Estimating photosinthetically active radiation from measured solar irradiance. Solar Energy 31(2):233-234. [ Links ]
2. BONAVENTURA, SM; R TECCHI & D VIGNALE. 1998. La vegetación en la Reserva de la Biósfera Laguna de Pozuelos. Pp. 43-59 en: Cajal, JL; JG Fernández &R Tecchi (eds.). Bases para la Conservación y Manejo de la Puna y Cordillera Frontal de Argentina. El rol de las Reservas de la Biósfera. FUCEMA-UNESCO. [ Links ]
3. BORGNIA, M; A MAGGI; M ARRIAGA; B AUED; BL VILÁ; ET AL. 2006. Caracterización de la vegetación en la Reserva de Biósfera Laguna Blanca (Catamarca, Argentina). Ecología Austral 16(1):29-45. [ Links ]
4. BRAUN WILKE, R. 1991. Tres recursos leñosos: Queñoa, Churqui y Tola. Pp. 43-50 en: García Fernández, J & R Tecchi (eds.). La Reserva de la Biósfera Laguna de Pozuelo: un ecosistema pastoril de los Andes Centrales. Instituto de Biología de Altura, UNJU, UNESCO-MAB, Montevideo. [ Links ]
5. BUONO, G; M OESTERHELD; V NAKAMATSU & JM PARUELO. 2010. Spatial and temporal variation of primary production of Patagonian wet meadows. Journal of Arid Environments 74:1257-1261. [ Links ]
6. CABRERA, AL. 1957. La vegetación de la Puna Argentina. Revista de Investigaciones Agrícolas. Buenos Aires 11(4):317-512. [ Links ]
7. CABRERA, AL. 1976. Regiones Fitogeográficas Argentinas. En Enciclopedia Argentina de Agricultura y Jardinería. Tomo II. Fasciculo1. Ed. ACME S.A.C.I. Buenos Aires. [ Links ]
8. CABRERA, AL. 1977. Flora de la Provincia de Jujuy. Parte II. Ed. INTA. Buenos Aires. [ Links ]
9. CABRERA, AL. 1978. Flora de la Provincia de Jujuy. Parte X. Ed. INTA. Buenos Aires. [ Links ]
10. CABRERA, AL. 1983. Flora de la Provincia de Jujuy. Parte VIII. Ed. INTA. Buenos Aires. [ Links ]
11. CASTAÑERA, M & A GONZÁLEZ. 1991. La vegetación de la Cuenca de Pozuelos. Pp. 43-50 en: García Fernández, J & R Tecchi (eds.). La Reserva de la Biosfera Laguna de Pozuelos: un ecosistema pastoril en los Andes Centrales. Instituto de Biología de Altura, UNJU, UNESCO-MAB, Montevideo. [ Links ]
12. CAJAL, JL. 1998a. Potencial de Conservación y factores de amenaza en la Puna y Cordillera Frontal. Pp. 289-296 en: Cajal, JL; JG Fernández & R Tecchi (eds.). Bases para la Conservación y Manejo de la Puna y Cordillera Frontal de Argentina. El rol de las Reservas de la Biósfera. FUCEMA-UNESCO. [ Links ]
13. CAJAL, JL. 1998b. Las unidades morfoestructurales, el clima, la vegetación y las poblaciones humanas en la Puna y Cordillera Frontal. Pp. 9-24 en: Cajal, JL; JG Fernández & R Tecchi (eds.). Bases para la Conservación y Manejo de la Puna y Cordillera Frontal de Argentina. El rol de las Reservas de la Biósfera. FUCEMA-UNESCO. [ Links ]
14. CAJAL, JL & SM BONAVENTURA. 1998. Densidad, biomasa y diversidad de mamíferos en la Puna y Cordillera Frontal. Pp. 191-213 en: Cajal, JL; JG Fernández & R Tecchi (eds.). Bases para la Conservación y Manejo de la Puna y Cordillera Frontal de Argentina. El rol de las Reservas de la Biósfera. FUCEMA-UNESCO. [ Links ]
15. CHUVIECO, ES. 2002. Teledetección Ambiental: la observación de la Tierra desde el espacio. Ariel Ciencia Ed. Madrid. Pp. 584. [ Links ]
16. CINGOLANI, AM; DM RENISONA; MR ZAK & MR CABIDO. 2004. Mapping vegetation in a heterogeneous mountain rangeland using landsat data: an alternative method to define and classify land-cover units. Remote Sensing Environment 92:84-97. [ Links ]
17. CONGALTON, RG; K BIRCH; R JONES & J SCHRIEVER. 2002. Evaluating remotely sensed techniques for mapping riparian vegetation. Computers and Electronics in Agriculture 37(1-3):113-126. [ Links ]
18. DI GREGORIO, A & LJM JANSEN. 2000. Land cover classification system (LCCS): Classification concepts and user manual. Food and Agriculture Organization of the United Nations (FAO), Roma. Pp. 251. [ Links ]
19. FERNÁNDEZ, OA & CA BUSSO. 1999. Arid and semi-arid rangelands: two thirds of Argentina. Pp. 41-60 en: Arnalds, O & S Archer (eds.). Case Studies of Rangeland Desertification. Agricultural Research Institute Report Nro. 200, Reykjavik (Islandia). [ Links ]
20. FRANCO, J. 2006. Energías renovables en la Puna. Un aporte realizado desde el Instituto de Energías no Convencionales. Pp. 47-51 en: Brown, A; U Martínez Ortiz; M Acerbi & J Corcuera (eds.). Situación Ambiental Argentina 2005. Fundación Vida Silvestre Argentina, Buenos Aires. [ Links ]
21. GARBULSKY, MF & JM PARUELO. 2004. Remote sensing of protected areas to derive baseline vegetation functioning characteristics. Journal of Vegetation Science 15:711-720. [ Links ]
22. GOLLUSCIO, RA; VA DEREGIBUS & JM PARUELO. 1998. Sustainability and range management in the Patagonian steppes. Ecología Austral 8:265-284. [ Links ]
23. GOLLUSCIO, RA; H BOTTARO; D RODANO; M GARBULSKY; S BOBADILLA; ET AL. 2009. Divergencias en la estimación de receptividad ganadera en el noroeste de la Patagonia: diferencias conceptuales y consecuencias prácticas. Ecología Austral 19:3-18. [ Links ]
24. GOLLUSCIO, RA; M ROMÁN; A CESA; D RODANO; H BOTTARO; M NIETO; ET AL. 2010. Aboriginal settlements of arid Patagonia: Preserving bio or sociodiversity? The case of the Mapuche pastoral Cushamen Reserve. Journal of Arid Environments 74(10):1329-1339. [ Links ]
25. GRIGERA, G & M OESTERHELD. 2006. Forage production monitoring systems parameterization of fPAR and RUE. Global vegetation Workshop. The University of Montana, Missoula MT, USA, 7 al 10 de agosto de 2006. [ Links ]
26. GRIGERA, G; M OESTERHELD & F PACIN. 2007a. Monitoring forage production for farmers' decision making. Agricultural Systems 94:637-648. [ Links ]
27. GRIGERA, G; M OESTERHELD; M DURANTE & F PACIN. 2007b. Evaluación y seguimiento de la productividad forrajera. Revista Argentina de Producción Animal 27:137-148. [ Links ]
28. GUERSCHMAN, JP; J PARUELO; C DI BELLA; M GIALLORENZI & F PACÍN. 2003. Land classification in the Argentine pampas using multitemporal landsat TM data. International Journal of Remote Sensing 17:3381-3402. [ Links ]
29. GUERSCHMAN, JP & JP PARUELO. 2005. Agricultural impacts on ecosystem functioning in temperate areas of North and South America. Global and Planetary Science 47:2 (4):170-180. [ Links ]
30. HUETE, A; K DIDAN; T MIURA; EP RODRIGUEZ; X GAO; ET AL. 2002. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote sensing of environment 83:195-213. [ Links ]
31. IRISARRI, JGN; M OESTERHELD; JM PARUELO & M TEXEIRA. 2011. Patterns and controls of aboveground net primary production in meadows of Patagonia. A remote sensing approach. Journal of Vegetation Science. En prensa. [ Links ]
32. JUSTICE, CO; E VERMOTE; JRG TOWNSHEND; R DEFRIES; DP ROY; ET AL. 1998. The Moderate Resolution Imaging Spectroradiometer (MODIS): land remote sensing for global change research. Geoscience and Remote Sensing 36(4):1228-1249. [ Links ]
33. KIESLING, R; A PÉREZ MOREAU; A PONTIROLI & FO ZULOAGA. 1983. Flora de la Provincia de Jujuy, República Argentina. Col. Cient. INTA. Buenos Aires. [ Links ]
34. LICHTENSTEIN, G & B VILÁ. 2003. Vicuna Use by Andean Communities: An overview. Mountain Research and Development 23(2):197-201. [ Links ]
35. MCCREE, KJ. 1972. Test of current definitions of photosynthetically active radiation against leaf photosynthesis data. Agricultural Meteorology 10(C):443-453. [ Links ]
36. MONTEITH, JL. 1972. Solar radiation and productivity in tropical ecosystems. Journal of Applied Ecology 9:747-766. [ Links ]
37. MORALES, CE. 2005. Pobreza, desertificación y degradación de tierras, Pp. 25-57 en: Morales, C & S Parada (eds.). Pobreza, desertificación y degradación de los recursos naturales. Naciones Unidas, Santiago de Chile. [ Links ]
38. NADIR, A & T CHAFATINOS. 1990. Los Suelos del NOA (Salta y Jujuy). Tomos I, II y III. Universidad Nacional de Salta. Argentina. [ Links ]
39. NOSETTO, MD; EG JOBBÁGY & JM PARUELO. 2005. Land-use change and water losses: the case of grassland afforestation across a soil textural gradient in central Argentina. Global Change Biology 11:1101-1117. [ Links ]
40. NOUVELLON, Y; D LO SEEN; S RAMBAL; A BEGUE; MS MORAN; ET AL. 2000. Time course of radiation use efficiency in a shortgrass ecosystem: consequences for remotely sensed estimation of primary production. Remote Sensing of Environment 71:43-55. [ Links ]
41. NOVARA, LJ. 2003. Catálogo de la flora de la Puna en el noroeste argentino. Aportes Bot. Salta, Ser. Misceláneas 2:1-56. [ Links ]
42. NOY-MEIR, I. 1973. Desert Ecosystems: Environment and Producers. Annual Review of Ecology and Systematics 4:25-51. [ Links ]
43. OESTERHELD, M; CM DIBELLA & H HERDILES. 1998. Relation between NOAA-AVHRR satellite data and stocking rate in grasslands. Ecological Applications 8:207-212. [ Links ]
44. OESTERHELD, M; R ARAGÓN; G GRIGERA; M OYARZÁBAL & M SEMMARTÍN. 2005. ¿Cómo deben percibir la heterogeneidad de la vegetación de los agroecosistemas? El caso de la Pampa Deprimida. Pp. 131-142 en: Oesterheld, M; MR Aguiar; CM Ghersa & JM Paruelo (eds.). La heterogeneidad de la vegetación de los agroecosistemas. Un homenaje a Rolando J.C. León. EFA, 1ra Edición, Buenos Aires. [ Links ]
45. PARUELO, JM & AR GOLLUSCIO. 1994. Range assessment using remote sensing in Northwest Patagonia (Argentina). Journal of Range Management 47:498-502. [ Links ]
46. PARUELO, JM; HE EPSTEIN; WK LAUENROTH & IC BURKE. 1997. ANPP estimates from NDVI for the Central grassland region of the United States. Ecology 78(3):953-958. [ Links ]
47. PARUELO, JM; MF GARBULSKY; JP GUERSCHMAN & M OESTERHELD. 1999. Caracterización regional de los recursos forrajeros de las zonas templadas de Argentina mediante imágenes satelitarias. Revista Argentina de Producción Animal 19:125-131. [ Links ]
48. PARUELO, JM; M OESTERHELD; CM DI BELLA; M ARZADUM; J LAFONTAINE; ET AL. 2000. Estimation of primary production of subhumid rangelands from remote sensing data. Applied Vegetation Science 3:189-195. [ Links ]
49. PARUELO, JM & MR AGUIAR. 2003. El impacto humano sobre los ecosistemas: el caso de la desertificación en Patagonia. Ciencia Hoy 13:48-59. [ Links ]
50. PARUELO, JM; RA GOLLUSCIO; JP GUERSCHMAN; A CESA; VV JOUVET; ET AL. 2004. Regional scale relationships between ecosystem structure and functioning: the case of the Patagonian steppes. Global Ecology and Biogeography 13:385-395. [ Links ]
51. PARUELO, JM. 2008. La caracterización funcional de ecosistemas mediante sensores remotos. Ecosistemas 17:4-22. [ Links ]
52. PETTORELLI, N; JO VIK; A MYSTERUD; JM GAILLARD; CJ TUCKER; ET AL. 2005. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecology & Evolution 20:503-510. [ Links ]
53. PIÑEIRO, G; M OESTERHELD & JM PARUELO. 2006. Seasonal variation in aboveground production and radiation use efficiency of temperate rangelands estimated through remote sensing. Ecosystems 9:357-373. [ Links ]
54. POSSE, G & A CINGOLANI. 2000. Environmental controls on NDVI and sheep production in the Tierra del Fuego steppe of Argentina. Applied Vegetation Science 3:253-260. [ Links ]
55. POTTER, CS; JT RANDERSON; CB FIELD; PA MATSON; PM VITOUSEK; ET AL. 1993. Terrestrial ecosystem production: a process model based on global satellite and surface data. Global Biogeochemical Cycles 7:811-841. [ Links ]
56. REBORATTI, C. 2006. Situación Ambiental en las Ecorregiones Puna y Altos Andes. Pp. 33-39 en: Brown, A; U Martínez Ortiz; M Acerbi & J Corcuera (eds.). Situación Ambiental Argentina 2005. Fundación Vida Silvestre Argentina, Buenos Aires. [ Links ]
57. RUNNING, SW; L QUEEN & M THORNTON. 2000. The Earth Observing System and forest management. Journal of Forestry 98(1):29-31. [ Links ]
58. RUTHSATZ, B & CP MOVIA. 1975. Relevamiento de las estepas andinas del noreste de la provincia de Jujuy. Fundación para la Educación la Ciencia y la Cultura. Pp. 127. [ Links ]
59. SALA, OE & AT AUSTIN. 2000. Methods of estimating aboveground net primary productivity. Pp. 31-43 en: Sala, OE; et al. (eds.). Methods in ecosystem science. Springer-Verlag New York, Inc. [ Links ]
60. SINCLAIR, TR & RC MUCHOW. 1999. Radiation use efficiency. Sparks, DL (ed.). Advances in Agronomy 65:215-263. [ Links ]
61. TÁLAMO, A; J TOLOBA; C TRUCCO & E ACUÑA. 2010. Unidades de vegetación y composición en sectores del Altiplano del noroeste de Argentina. I. Ambientes de estepas. Ecología en Bolivia 45(1):4-19. [ Links ]
62. ZOFFOLI, ML; P KANDUS; N MADANES & DH CALVO. 2008. Seasonal and interannual analysis of wetlands in South America using NOAA-AVHRR NDVI time series: the case of the Parana Delta Region. Landscape Ecology 23:833-848. [ Links ]

