










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.22 no.1 Córdoba abr. 2012
ARTÍCULO ORIGINAL
Respuesta diferenciada a la sequía de plantas jóvenes de Prosopis chilensis, P. flexuosa y sus híbridos interespecíficos: implicancias para la reforestación en zonas áridas
Diego López Lauenstein1,*; María E Fernández2 & Aníbal Verga1
1. Instituto de Fisiología y Recursos Genéticos Vegetales (IFRGV), Instituto Nacional de Tecnología Agropecuarias (INTA). Córdoba. Argentina.
2. Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)-Argentina; Instituto Nacional de Tecnología Agropecuaria, EEA Bariloche.
* Instituto de Fitopatología y Fisiología Vegetal (IFFIVE), Instituto Nacional de Tecnología Agropecuarias (INTA). Camino a 60 cuadras km 5.5. Córdoba. Argentina. TE: +543514973636. Email: dlopezlauenstein@iffive.inta.gov.ar
Recibido: 24 de abril de 2011;
Fin de arbitraje: 21 de agosto de 2011;
Revisión recibida: 29 de septiembre de 2011;
Aceptado: 6 de noviembre de 2011
RESUMEN. En la región del Chaco Árido existen procesos graves de degradación de los ecosistemas. La reforestación con materiales adaptados a las principales limitantes climáticas (sequía) es una técnica que puede contribuir a revertir dichos procesos degradativos. En relación a sus requerimientos hídricos, Prosopis flexuosa se desarrolla en llanos, ambientes más secos, mientras que Prosopis chilensis crece al pie de las sierras, a orillas de cursos de agua temporarios. Varios estudios genéticos han demostrado la existencia de híbridos interespecíficos en las áreas de contacto de las dos especies. Los objetivos de este trabajo fueron (1) determinar la respuesta diferencial en crecimiento y supervivencia de plantines de P. flexuosa, P. chilensis y sus híbridos interespecíficos ante condiciones controladas de estrés hídrico, y (2) determinar la influencia del tratamiento de rusticación en vivero sobre estos procesos posteriormente a la implantación en el campo. En los ensayos bajo condiciones controladas, P. flexuosa fue la especie que menos creció ante buenas condiciones de disponibilidad hídrica, pero fue la menos afectada ante condiciones de estrés. Por el contrario, P. chilensis fue la especie más susceptible al estrés hídrico. Los individuacs híbridos tuvieron un crecimiento similar al de P. chilensis, tanto ante buenas condiciones de disponibilidad hídrica como con déficit, pero mayor supervivencia que esta especie ante estrés severo, semejante a P. flexuosa. Bajo condiciones a campo, tanto P. chilensis como los híbridos se observó un efecto positivo de la rusticación, sin embargo en P. flexuosa solamente se pudo observar ese efecto en el sitio de plantación con mayor restricción hídrica. Estos resultados abren interrogantes acerca de los mecanismos diferenciales que permitirían a los híbridos alcanzar altas tasas de crecimiento, sin ir en detrimento de la resistencia al estrés. Esta combinación de caracteres resalta la necesidad de focalizar estudios ecofisiológicos (mecanismos de tolerancia a estrés hídricos) y genéticos (estabilidad y segregación de caracteres en la descendencia) en este taxón.
Palabras clave: Algarrobos; Hibridación; Adaptación; Estrés hídrico; Rusticación; Chaco árido.
ABSTRACT. Differences in drought responses of seedlings of Prosopis chilensis, P. flexuosa and interspecific hybrids: implications for reforestation in arid zones: In the arid Chaco region are severe processes of degradation of ecosystems. The reforestation with adapted species to the main climate limitation (drought) is a technique that can reverse such degradation processes. According to their water requirements, Prosopis flexuosa grows in plain drier environments, while Prosopis chilensis grows next to the mountains, on the banks of temporary rives. Genetic studies have demonstrated the existence of interspecific hybrids in contact areas between these species. The objectives of this study were (1) to determine the differential response in growth and survival of seedlings of P. flexuosa, P. chilensis and interspecific hybrids under controlled water stress conditions, and (2) to determine the influence of an imposed treatment of drought hardening in the nursery on the performance of plants in the field. In controlled water stress conditions, P. flexuosa showed lest growth under control treatment (no stress), however under stress treatment was the least affected by water stress. By contrast, P. chilensis was the most susceptible to drought stress, response that could be enhanced by hardening treatment before planting. The hybrid was similar to P. chilensis (under stress treatment and control treatment), but showed grate survival rate, similar to P. flexuosa. Both P. chilensis and hybrids showed a positive effect of hardening, but in P. flexuosa only this effect was observed at the driest site. The combination of these two characters in general are given as a mutual commitment, and focus highlights the need for ecophysiological studies (mechanisms) and genetic (stability and segregation of characters in the offspring) in this taxon.
Keywords: Algarrobos; Hybridation; Adaptation; Drought stress; Hardening; Arid Chaco.
INTRODUCCIÓN
La región del Chaco Árido ocupa una superficie de 80000 km2. Abarca el oeste de la provincia de Córdoba, el sur de La Rioja, el sur de Catamarca y el norte de San Luis. El relieve es llano, de clima cálido y seco, con precipitaciones estivales que van desde 500 mm/año en el este a 300 mm/año en el extremo oeste, en el límite con la región fitogeográfica del Monte (Karlin et al. 1992). Es la porción más seca y menos productiva del Gran Chaco (Morello et al. 1985). Esta región está siendo afectada por una intensiva utilización de los recursos naturales. La explotación forestal y la actividad ganadera han dado origen a procesos de degradación que se manifiestan en el reemplazo del bosque por matorrales o "fachinal", con escasas posibilidades de uso productivo (Bocco et al. 2007). La recuperación del bosque natural está limitada por el déficit hídrico, que en la región llega a 700 mm/año (Karlin et al. 1992). Por otro lado, se estima para la región chaqueña, como consecuencia del cambio climático global, incrementos de la temperatura de entre 1.7 y 3.9 °C, y un aumento en las precipitaciones que acentúa su carácter monzónico, acompañado además por mayor pluviometría en los meses de diciembre, enero y febrero, alternados con largos períodos de sequía (IPCC 2007). Es por esto que se hace necesaria la intervención del hombre para asegurar la estabilidad del ecosistema y su sustentabilidad, tanto ecológica como económica y social.
Los árboles multipropósito constituyen una herramienta buena para la recuperación de la capacidad productiva de los ecosistemas áridos (Verga 2000a), de manera que la repoblación con dichas especies genera condiciones favorables para la recuperación natural de todo el ecosistema. En este sentido, los algarrobos, diversas especies del género Prosopis, son un conjunto de árboles de gran importancia en la región de estudio. Son plantas pioneras con un alto potencial biológico para la diseminación y colonización. Como leguminosas tienen la capacidad de fijar nitrógeno atmosférico en simbiosis con bacterias del género Rhizobium (Acosta et al. 1994), desarrollan raíces profundas que disminuyen la competencia por agua con especies herbáceas, mejoran el balance hídrico del sistema, aportan nutrientes de las capas subsuperficiales y, en algunos casos, se independizan de las precipitaciones (Demaio et al. 2002). Producen leña y carbón de buena calidad, madera de características físicas y mecánicas extraordinarias (Perpiñan & Pietrarelli 1992), frutos de contenido proteico y energético elevado (óptimo tanto para su uso como forrajera como para el consumo humano) y otros productos secundarios (Karlin 1983). Asimismo, los algarrobos pueden constituirse en una alternativa productiva como parte de sistemas silvopastoriles (Bregaglio et al. 2001). En la región del Chaco Árido, y en relación a sus requerimientos hídricos, P. flexuosa se desarrolla naturalmente en llanos, ambientes más secos, mientras que P. chilensis crece al pie de las sierras, a orillas de cursos de agua temporarios (Cabido et al. 1993). Ambas especies entran en contacto en el pedemonte de las sierras, donde se observa una gran proliferación de híbridos naturales interespecíficos (Verga 1995).
Una de las limitantes principales para la reforestación en ambientes caracterizados por la escasez de lluvia y temperaturas elevadas es la capacidad de supervivencia y crecimiento inicial de los plantines durante el período inmediato posterior a la plantación (Gupta 1995). El desarrollo de estrategias adaptativas durante las etapas tempranas de la ontogenia de las plántulas (primer ciclo de crecimiento) parecen determinantes para su supervivencia en ambientes con estrés hídrico (Pratalongo et al. 2003). Se ha demostrado que plantas sometidas a un período de estrés moderado (rusticación) pueden desarrollar resistencia y soportar períodos de estrés prolongados; esto mejora su supervivencia y crecimiento ante condiciones limitantes (Guarnaschelli et al. 2006). Para desarrollar resistencia las plantas deben ser capaces de modificar sus estructuras (e.g., a través del aumento de la proporción de biomasa de raíces, modificación de la estructura del xilema, etc.) o bien mediante cambios a nivel fisiológico y/o bioquímico (Vilagrosa et al. 2003; Grossnickle 2005). Cabe señalar, sin embargo, que muchas veces esas modificaciones no mejoran el desempeño de las plantas en crecimiento y supervivencia después del transplante (Villar Salvador et al. 2004).
Los objetivos de este trabajo fueron (1) determinar la respuesta diferencial en crecimiento y supervivencia de plantines de P. flexuosa, P. chilensis y sus híbridos interespecíficos ante condiciones controladas de estrés hídrico, y (2) determinar la influencia del tratamiento de rusticación en vivero sobre estos procesos posteriormente a la implantación en el campo.
MATERIALES Y MÉTODOS
Los estudios se realizaron con semillas de individuos puros de P. chilensis y P. flexuosa y de híbridos interespecíficos. Para la selección de las semillas, se cosecharon árboles semilleros provenientes de poblaciones naturales de la región del Chaco árido. Se tomaron muestras de hojas y frutos y se realizó la clasificación de los árboles semilleros mediante taxonomía numérica de caracteres morfológicos cuantitativos y cualitativos, según la metodología propuesta por Verga & Gregorius (2007). Como segunda comprobación para la integración del material de estudio se realizaron análisis genéticos a través de la descendencia mediante el marcador isoenzimático ADH (Verga 1995). De esta manera quedaron conformados los "pooles" de semillas pertenecientes a cada taxón (P. chilensis, P. flexuosa e híbridos interespecíficos).
Ensayo en invernáculo
Se trabajó en un invernáculo con suplementación de luz artificial hasta alcanzar una intensidad de 500 µmol.m-2.s-1. Si bien la intensidad de luz fue inferior a la que normalmente se alcanzan a campo, las plantas no mostraron síntomas de deficiencia de luz tales como alargamiento de entrenudos, amarillamiento de hojas, etc. Se utilizaron macetas de 3 L y un sustrato compuesto por dos partes de mantillo y una parte de arena. Se controló la humedad del sustrato mediante pesadas cada 48 h. Durante los primeros dos meses posteriores a la siembra las plantas crecieron sin restricciones hídricas. Durante los tres meses siguientes fueron separadas en tres grupos de acuerdo a los tres tratamientos de riego (control, estrés hídrico moderado y estrés hídrico severo). Se utilizaron 30 plantas por taxón.
La magnitud del estrés hídrico se estimó sobre la base de una curva de potencial hídrico medido en pre-alba en relación con la humedad del suelo, conteniendo puntos para las dos especies puras y para los híbridos interespecíficos debido a que no se verificaron diferencias entre los taxones (Figura 1). Por encima de 10% de humedad gravimétrica, los potenciales hídricos de las plantas se mantuvieron constantes en torno a -0.5 MPa. Por debajo de esa humedad, el potencial de pre-alba disminuyó drásticamente y pasó de -1 MPa a -7 MPa, con una variación de solo 5% de humedad del suelo. Por lo tanto, el tratamiento de estrés hídrico moderado el potencial pre-alba fue aproximadamente -3 MPa y el tratamiento de estrés severo fue aproximadamente -5 MPa. El potencial hídrico fue medido con una cámara de presión Bio-control modelo Π1 (Bio-control, Buenos Aires, Argentina). Así, en el tratamiento control el contenido gravimétrico de agua en suelo fluctuó entre 15% y 25%, en el de estrés moderado entre 6 y 15%, y en el de estrés severo entre 5 y 10%.

Figura 1. Relación entre el contenido gravimétrico de agua en el de suelo (%) y el potencial hídrico en prealba (MPa) medido en P. chilensis, P. flexuosa e híbridos interespecíficos. Los individuos de los tres taxa presentaron una respuesta común por lo que se indican con un único símbolo.
Figure 1. Relationship between gravimetric soil water content (%) and pre-dawn water potential measured in P. chilensis, P. flexuosa and their interspecific hybrids. Individuals of all three taxa presented a similar response, therefore, only one symbol is used.
A partir del segundo mes de iniciado los tratamientos se midieron semanalmente el diámetro en la base del tallo, la altura y el número de hojas de cada plantín. El ensayo tuvo una duración de 5 meses (3 meses de tratamiento de estrés hídrico) y al final del ensayo las plantas fueron cosechadas y separadas en hojas, tallos y raíces. Se tomaron los pesos frescos y se llevaron a estufa a 70 ºC por 72 h para calcular el peso seco de las distintas fracciones y la relación peso seco de raíz/peso seco total. También se midió el área foliar de cada planta mediante el software Hojas (Verga 2000b).
Con cada una de las variables analizadas se calculó para cada taxón, el índice de plasticidad fenotípica RDPI (del inglés, Relative Distance Plasticity Index) propuesto por Valladares et al. (2006). El índice RDPI fue calculado como el valor absoluto de la diferencia entre dos valores para cada especie y combinación de tratamientos, dividido por la suma de estos dos valores.
RDPI = ∑(dij → j'j')/(xi'j'+ xij)/n
donde dij →j'j es la distancia fenotípica entre dos individuos del mismo taxón creciendo en distintos tratamientos de estrés hídrico. Este índice varía desde 0 (no plasticidad) y 1 (máxima plasticidad), y permite obtener variación entorno a la media y por lo tanto está sujeta a comparaciones estadísticas (Valladares et al. 2006).
Ensayos a campo
Se llevaron a cabo dos ensayos a campo, en sitios ubicados en los extremos de precipitación de la región del Chaco árido argentino: El Totoral, provincia de La Rioja (31º44'59'' S, 66º30'34'' O) y Chancaní, provincia de Córdoba (31º26'25'' S, 65º26'59'' O), Argentina. La distribución anual de las precipitaciones y las temperaturas medias mensuales para los dos sitios de plantación muestran que en Chancaní hay mayor precipitación y que en El Totoral el período seco es más prolongado (Hijmans et al. 2005) (Figura 2). La precipitación media anual para Chancaní es 533 mm, mientras que para El Totoral es de 396 mm. También se observan diferencias en la temperatura media, alrededor de 2 °C superior en El Totoral para los meses de octubre a marzo. La mayor temperatura sumada a la menor precipitación de El Totoral implica una diferencia aún mayor en el déficit hídrico entre sitios.
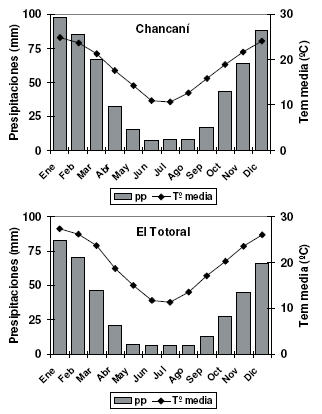
Figura 2. Climograma de los dos sitios de ensayo a campo, Chancaní (pcia. de Córdoba) y El Totoral (pcia. de la Rioja), Argentina.
Figure 2. Climogram of the two sites where field experiments were installed, Chancaní (Córdoba province) and El Totoral (La Rioja province), Argentina.
La principal diferencia edáfica entre sitios se observó en la textura del suelo: el suelo de El Totoral es arenoso franco y el de Chancaní es franco (Tabla 1), acentuando aun más las diferencias en disponibilidad hídrica entre sitios. También en Chancaní el contenido de materia orgánica es mayor (Tabla 1). Los valores de conductividad eléctrica (dS/m), indican que no existen limitantes en cuanto a salinidad en ninguno de los dos sitios.
Tabla 1. Características edáficas en las parcelas de ensayo, Chancaní (pcia. de Córdoba) y El Totoral (pcia. de La Rioja), correspondientes a los primeros 15 cm de suelo.
Table 1. Soil characteristic of field trials, Chancaní (Córdoba province) and El Totoral (La Rioja province), data obtained for the first 15 cm of soil depth.

Los plantines que se utilizaron para los dos ensayos a campo se produjeron en el vivero de la Estación Forestal Villa Dolores del INTA (Instituto Nacional de Tecnología Agropecuaria de Argentina, en la provincia de Córdoba). Se utilizaron envases plásticos sin fondo de 20 cm de alto por 8 cm de diámetro. Se utilizó un sustrato compuesto por dos partes de tierra y una parte de arena. A partir del primer mes de edad los plantines fueron separados en tres grupos y sometidos a tres regímenes de riego: tratamiento 1 (cinco riegos/semana), tratamiento 2 (tres riegos/semana) y tratamiento 3 (dos riegos/semana). Se utilizó el mismo volumen de agua en cada riego variando la frecuencia de los mismos entre tratamientos. Dichos regímenes fueron continuados por 5 meses hasta el momento de la plantación a campo. Los ensayos a campo fueron instalados en marzo de 2006 de acuerdo a lo propuesto por Catalán et al. (1994) como la mejor época de plantación. El lote fue desmontado previamente y se hicieron hoyos de 40 cm de profundidad para la implantación. El marco de plantación fue de 2 m x 2 m. El diseño de los ensayos a campo fue en parcelas totalmente aleatorizadas, con 6 plantas de cada taxón/parcela y 4 repeticiones. Al momento de plantación se tomó la primera medida de diámetro a la base del tallo y luego se tomaron medidas periódicas de diámetro a la base del tallo y de supervivencia durante dos años. A partir de las medidas de diámetro a la base del tallo se calculó la tasa de crecimiento relativo como el crecimiento en diámetro a la base en relación con el diámetro inicial (Dab final-Dab inicial)/Dab inicial.
Análisis estadístico
Para el análisis estadístico de los datos se utilizó el software Infostat (2009). Se realizaron análisis de la varianza (ANOVA) seguidos de pruebas de Duncan para comparar las medias, con un nivel de significación de 5% (P≤0.05). Para el ensayo bajo condiciones semicontroladas, se aplicó un diseño factorial para la determinación de los tratamientos, donde el genotipo tuvo tres niveles: P. chilensis, P. flexuosa e híbridos interespecíficos. El factor humedad también tuvo tres niveles: riego control, estrés hídrico moderado y estrés hídrico severo. Para cada ensayo a campo también se aplico un diseño factorial con los mismos dos factores y sus respectivos niveles.
RESULTADOS
Ensayo de invernáculo
El análisis de los parámetros de producción de biomasa en todos sus componentes (PS total, PS tallo, PS raíz y PS foliar) mostró que tanto P. chilensis como los híbridos tuvieron un crecimiento y un tamaño de planta mayor que P. flexuosa bajo buenas condiciones de disponibilidad hídrica (tratamiento control) (Figura 3). Por otro lado, los dos tratamientos de estrés hídrico afectaron el crecimiento la producción de materia seca y el número de hojas tanto en las especies puras como en los híbridos. En cuanto a las relaciones alométricas, la proporción de raíces no se modificó significativamente bajo los tratamientos de estrés en ningún taxón (P=0.07), sin embargo, en las dos especies puras se observó un aumento en la proporción de raíces bajo los tratamientos de estrés (Figura 3E).

Figura 3. Peso Seco Total (A), PS tallo (B), PS raíz (C), PS foliar (D), proporción de raíz (E) y número de hojas (F) en P. chilensis, P. flexuosa e Híbrido interespcífico. Barras blancas: tratamiento control, barras grises tratamiento estrés moderado y barras negras: tratamiento estrés severo. Media y Error Estándar. Letras distintas significan diferencias entre tratamientos (prueba de Duncan P≤.05).
Figure 3. Total dry weight(A), shoot dry weight (B), root dry weight (C), leaf dry weight (D), leaf area(E) and number of leaves in P. chilensis, P. flexuosa and interspecific hybrids. White bars: control treatment, gray bars: moderate stress and black bars: severe stress. Means and standard errors. Different letters indicate significant differences between stress treatments (P≤0.05).
La altura de los individuos no varió entre las dos especies y los híbridos pero sí entre distintos niveles de humedad; tanto las especies puras como el híbrido el tratamiento control fue superior a los dos tratamientos de estrés (Figura 4). Por el contrario, P. chilensis y los híbridos tuvieron un diámetro basal superior al de P. flexuosa, y también hubo diferencias entre el tratamiento control y los dos tratamientos de estrés hídrico (Figura 4A).

Figura 4. Diámetro a la base del tallo (Dab) (A) y Altura (B) en P. chilensis, P. flexuosa e híbridos interespecíficos. Barras blancas: tratamiento control, barras grises tratamiento estrés moderado y barras negras: tratamiento estrés severo Media y error estándar. Letras distintas indican diferencias significativas entre tratamientos (prueba de Duncan P≤.05).
Figure 4. Shoot basal diameter (Dab) (A) and Heigh (B) in P. chilensis, P. flexuosa and interespecific hybrid. White bars: control treatment, gray bars: moderate stress and black bars: severe stress. Means and standard errors. Different letters indicate significant differences between stress treatments (P≤.05).
Las variables relacionadas a las hojas (PS foliar, AF y número de hojas) fueron las más afectadas por el estés hídrico, aunque solo se documentaron diferencias significativas para el número de hojas (P=0.003) entre los taxones (Figura 5). En segundo término, las variables de PS total, PS raíz y PS tallo tuvieron un efecto intermedio y en todas P. flexuosa fue la especie menos afectada. En el resto de las variables estudiadas, el estrés hídrico tuvo un bajo efecto, destacándose P. flexuosa como la menos afectada en el diámetro a la base y el híbrido el menos afectado en altura. Por último, no se registraron diferencias entre los taxones en la proporción de raíces (P=0.08).

Figura 5. RDPI (relative distance plasticity index) para cada una de las variables estudiadas. Letras distintas indican diferencias significativas entre taxones (prueba de Duncan P=0.05).
Figure 5. RDPI (relative distance plasticity index) for each studied trait. Different letters means statistical different between taxas (Duncan test P=0.05).
Considerando la supervivencia de las plantas en el ensayo de condiciones semi-controladas, solo bajo el tratamiento de estrés hídrico severo hubo plantas que murieron hacia el final del ensayo, pero en el caso de P. flexuosa y los híbridos el porcentaje de supervivencia fue superior al de P. chilensis (Tabla 2).
Tabla 2. Supervivencia al final del ensayo medida en P. chilensis, P. flexuosa e híbridos interespecíficos que crecieron bajo tratamientos de estrés hídrico moderado y severo, y tratamiento control (sin deficiencia de agua).
Table 2. Survival at the end of the experiment measured in P. chilensis, P. flexuosa and interespecific hybrid grown under water stress treatments (moderate and severe levels) and control treatment.

Ensayo a campo
Las plantas llevadas a campo diferían significativamente en su tamaño al momento de la implantación como producto de los tratamientos de riego diferencial (Figura 6). Asimismo, se pudo apreciar nuevamente el mayor tamaño de P. chilensis y el híbrido en comparación con P. flexuosa en el tratamiento con mayor régimen de riego.

Figura 6. Diámetro en la base del tallo en el momento de plantación en P. chilensis, P. flexuosa e híbridos interespecíficos. Régimen de riego en vivero: Tratamiento 1 = cinco riegos por semana. Tratamiento 2 = tres riegos por semana. Tratamiento 3 = dos riegos por semana. Media y error estándar. Letras distintas indican diferencias significativas entre tratamientos (prueba de Duncan P≤.005).
Figure 6. Shoot basal diameter of P. chilensis, P. flexuosa and interspecific hybrids at the time of field plantation. Irrigation frequency in the nursery: Treatment 1 = five times a week. Treatment two = three times a week. Treatment three = two times a week. Means and standard error. Different letters indicate significant differences between stress treatments within each species (P≤.005).
En el ensayo de Chancaní, el extremo más húmedo de la región del Chaco árido, se observó una mayor tasa de crecimiento relativo en diámetro a la base del tallo en aquellas plantas que provenían de los tratamientos con menor régimen de riego en el vivero, tanto en los híbridos como en P. chilensis, pero no en P. flexuosa. En el ensayo de El Totoral, es decir, el sitio con mayores restricciones hídricas, no hubo diferencias significativas en la tasa de crecimiento relativo entre los régimen de riego, pero en las especies puras las medias de los tratamientos con menor régimen fueron más altas. En los híbridos esa tendencia no fue tan marcada (Figura 7).

Figura 7. Tasa de crecimiento relativo en diámetro de P. chilensis, P. flexuosa e Híbrido interespecífico en Chancaní y en El Totoral. Régimen de riego en vivero: Tratamiento 1 = cinco riegos por semana. Tratamiento 2 = tres riegos por semana. Tratamiento 3 = dos riegos por semana. Medias y Error Estándar. Letras distintas indican diferencias significativas entre tratamiento (prueba de Duncan P≤.05).
Figure 7. Relative diameter growth rate of P. chilensis, P. flexuosa and interspecific hybrids in two field trials, Chancaní and Totoral after two year since plantation. Irrigation frequency in the nursery: Treatment 1 = five times a week. Treatment two = three times a week. Treatment three = two times a week. Means and standard error.
A diferencia de las tendencias observadas en el crecimiento, los patrones no fueron tan claros al analizar la supervivencia de los plantines provenientes de los distintos tratamientos de régimen riego en vivero (Figura 8). En las especies puras no se observaron diferencias significativas en supervivencia de los diferentes régimen de riego en ninguno de los dos sitios (P>0.1), pero en los híbridos el tratamiento con tres riegos semanales en vivero tuvo un comportamiento errático. En Chancaní la supervivencia fue significativamente menor (P=0.009) y en El Totoral significativamente mayor (P=0.03).
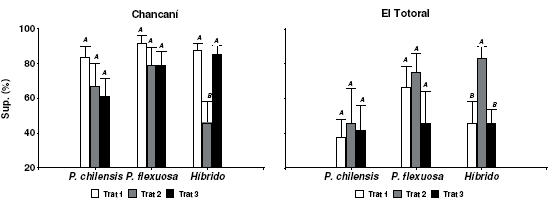
Figura 8. Porcentaje de supervivencia de P. chilensis, P. flexuosa e Híbrido interespecífico en dos ensayos a campo (Chancaní y El Totoral) al segundo año desde la plantación. Régimen de riego en vivero: Tratamiento 1 = cinco riegos por semana. Tratamiento 2 = tres riegos por semana. Tratamiento 3 = dos riegos por semana. Medias y Error Estándar. Letras distintas indican diferencias significativas entre tratamiento (prueba de Duncan P≤.05).
Figure 8. Survival of P. chilensis, P. flexuosa and Interespecific Hybrid in two field trials, Chancaní and El Totoral after two years since plantation. Irrigation frequency in the nursery: Treatment 1 = five times a week. Treatment 2 = three times a week. Treatment 3 = two times a week. Means and standard error.
Como es de esperarse en función de la disponibilidad hídrica en cada sitio, sí se observó mayor supervivencia general en el sitio más húmedo (Chancaní) que en el más seco (El Totoral), para todas las especies y tratamientos de rusticación. La media para Chancaní fue 75% y en El Totoral de 54% (P=0.0005).
DISCUSIÓN
La disponibilidad de agua fue un factor importante en la fase inicial de crecimiento de las plantas ya que afectó a todas las variables relacionadas con la producción de biomasa, tanto en las especies puras como en los híbridos interespecíficos. Prosopis flexuosa, que crece naturalmente en lugares donde los períodos de sequía son prolongados (Cabido et al. 1993), fue la especie con menor crecimiento bajo condiciones de buena disponibilidad hídrica, pero también fue la especie que menos redujo la acumulación de materia seca, comportamiento esperable en especies con mayor resistencia al estrés (Villagra & Cavagnaro 2005). La menor reducción en el crecimiento podría deberse a las características foliares y los mecanismos celulares que permiten comportarse como una especie relativamente tolerante a la sequía, con alta supervivencia ante estrés (Cavagnaro & Passera 1993; Villagra et al. 2010). Por el contrario, P. chilensis fue la especie más afectada por el estrés hídrico, con mayores reducciones en producción de biomasa y crecimiento en diámetro, hasta el punto que bajo estrés severo o sequías prolongadas su supervivencia se vio seriamente comprometida. Los híbridos no presentaron una respuesta intermedia entre la de las especies puras: crecieron en forma similar a la de P. chilensis (alto crecimiento con alta disponibilidad de agua, y alta reducción del crecimiento ante estrés), y sobrevivieron en forma similar a P. flexuosa ante condiciones de estrés severo.
Las especies de climas áridos se caracterizan por tener hojas de pequeño tamaño, compuestas y con alto peso foliar específico (Gibson 1998). En el género Prosopis, se han observado características foliares que pueden considerarse como adaptaciones a la sequía (Vilela & Palacios 1997). Sin embargo, debido a la distribución de los nichos específicos de cada especie, estas características foliares varían de acuerdo a los requerimientos ecológicos de cada una (Villagra et al. 2010). Prosopis chilensis, que crece en zonas donde puede tener acceso a aportes extra de agua, posee foliólulos más grandes, con mayor área foliar y una relación peso foliar/área foliar menor a P. flexuosa (Verga 1995), a la que un menor tamaño de hojas le permitiría reducir la superficie transpirante y mejorar el intercambio de calor disminuyendo la temperatura foliar (Gibson 1998). Los caracteres morfológicos foliares de los híbridos son intermedios al de las especies puras (Verga 1995). Asimismo, P. flexuosa reaccionó al estrés hídrico desprendiendo gran cantidad de hojas a medida que el estrés transcurría. Al tener gran cantidad de hojas y de pequeño tamaño pudo realizar un mejor ajuste del área foliar y así, evitaría más eficientemente las fallas en el sistema hidráulico (Vilagrosa et al. 2003). Por el contrario, P. chilensis y los híbridos perdieron menos cantidad de hojas pero de mayor tamaño, por lo que la biomasa y el área foliar cayeron abruptamente. La abscisión de hojas es un efectivo mecanismo de evitación de la sequía, pero tiene asociado un alto costo en ganancia de carbono y crecimiento (Pallardy & Lowenstein 2004). Ante un estrés severo los tres grupos presentaron una caída casi total de sus hojas, pero P. chilensis presentó además una alta proporción de individuos muertos.
Las diferencias de tamaño de las plantas que fueron llevadas a los ensayos de campo, producto de los tratamientos de régimen de riego en vivero, evidencian el efecto de la rusticación. En general los tratamientos que tuvieron menor disponibilidad de agua en vivero crecieron mejor después del transplante. Esto se pudo observar principalmente en P. chilensis, la especie más susceptible al estrés, la que presentó diferencias significativas en el crecimiento entre las plantas control y las provenientes de estrés hídrico severo (no así, moderado) en ambos sitios. En el caso del híbrido, el beneficio de una rusticación severa se manifestó solo en el sitio húmedo, mientras que en P. flexuosa, no se verificó un beneficio (al menos estadísticamente significativo) de la rusticación en ninguno de los dos sitios. Cabe destacar que, a pesar de la ausencia de respuesta diferencial entre tratamientos, se observó un buen desempeño en crecimiento de los individuos de P. flexuosa a campo, similar a los de mejor desempeño de P. chilensis y los híbridos. Resultados similares en cuanto al efecto positivo de la rusticación en el crecimiento de P. chilensis y los híbridos han sido obtenidos por otros autores para otras especies. Por ejemplo, Guarnaschelli et al. (2006) encontraron un mejoramiento en el estado hídrico y en el intercambio gaseoso después del transplante en plantas de Eucalyptus globulus sometidas a un período previo de rusticación, en comparación con individuos no rusticados, que les permitió obtener mayores crecimientos.
A diferencia de los patrones en el crecimiento, no se observó un efecto claro de la rusticación en vivero sobre la supervivencia después del transplante. Esto puede deberse a que en el campo la muerte de las plantas no solo es provocada por el efecto del estrés hídrico, sino también por otros factores de estrés abióticos (e.g., térmico, lumínico) y biótico (e.g., defoliación por herbívoros). En este sentido, en los ensayos a campo se encontró una gran cantidad de plantas comidas por roedores, por lo que no se pueden obtener conclusiones claras en relación con el efecto particular de la rusticación en vivero sobre la supervivencia. En concordancia con lo mostrado por Catalán et al. (1994), no se observaron diferencias en supervivencia entre las dos especies parentales y sus híbridos interespecíficos.
Uno de los procedimientos que se utilizan para favorecer la recuperación de zonas degradadas es la implantación con especies nativas (Montagnini et al. 1999). Estos procesos son costosos y requieren un análisis pormenorizado ecológico y económico, especialmente en zonas áridas, en las que las limitantes ambientales son severas y el riesgo de fallas es muy alto. Los resultados de este trabajo permiten vislumbrar alternativas de manejo, ligadas a la rusticación en vivero, aplicables para un mayor éxito de crecimiento a campo en los taxones más susceptibles al estrés, como es P. chilensis. Asimismo, un proceso de rusticación en la especie más resistente, P. flexuosa, no redundaría en una ventaja diferencial luego del transplante. A partir de estos resultados, surge la necesidad de ajustar las técnicas de producción de plantines en vivero de estas especies, considerando también condiciones generales (mayor o menor grado de estrés) del potencial sitio de plantación.
Por otro lado, un resultado interesante de este trabajo es el comportamiento de los individuos híbridos. Como se mencionó, si bien morfológicamente son individuos "intermedios" entre ambas especies puras, su resistencia al estrés híbrido no tendría este carácter. Los híbridos presentaron claramente un alto potencial de crecimiento ante buenas condiciones hídricas, similar al de P. chilensis. Sin embargo, su supervivencia fue muy alta ante condiciones de estrés severo, al nivel de la especie pura más tolerante. Estos resultan abren interrogantes acerca de los mecanismos diferenciales que le permitirían alcanzar altas tasas de crecimiento, sin presentar un compromiso aparente con la resistencia al estrés. La combinación de ambos caracteres resalta la necesidad de focalizar estudios ecofisiológicos (mecanismos) y genéticos (estabilidad y segregación de caracteres en la descendencia) en este taxón.
BIBLIOGRAFÍA
1. ACOSTA, M; L OLIVA & A ABRIL. 1994. Colección de Rhizobios de Prosopis arbóreos en al zona semiárida de la Provincia de Córdoba (Argentina). Ciencia del Suelo 12:38-40. [ Links ]
2. BOCCO, M; R COIRINI; U KARLIN & A VON MULLER. 2007. Evaluación socioeconómica de sistemas productivos sustentables en el Chaco Árido, Argentina. Zonas Áridas 11(1):70-84. [ Links ]
3. BREGAGLIO, M; U KARLIN & R COIRINI. 2001. Efecto del desmonte selectivo sobre la regeneración de la masa forestal y la producción de pasturas, en el chaco árido de la provincia de Córdoba, Argentina. Multequina 10:17-24. [ Links ]
4. CABIDO, M; C GONZÁLEZ; A ACOSTA & S DÍAZ. 1993. Vegetation changes along a precipitation gradient in Central Argentina. J. Veg. Sci. 109: 5-14. [ Links ]
5. CATALÁN, L; C CARRANZA; L GONZÁLEZ; U KARLIN & M LEDESMA. 1994. Afforestation trials with Prosopis chilensis (Mol.) Stuntz and Prosopis flexuosa D.C. in the Dry Chaco. Argentina, For. Ecol. Manage. 70:113-119. [ Links ]
6. CAVAGNARO, J & C PASSERA. 1993. Relaciones hídricas de Prosopis flexuosa(algarrobo dulce) en el Monte, Argentina. Pp 73-78 en: IADIZA (ed.). Contribuciones Mendocinas a la Quinta Reunión Regional para América Latina y el Caribe de la Red de Forestación del CIID. Conservación y Mejoramiento de Especies del Género Prosopis. Mendoza, Argentina. [ Links ]
7. DEMAIO, P; U KARLIN & M MEDINA. 2002. Árboles nativos del centro de Argentina. Ed. L.O.L.A. Pp. 210. [ Links ]
8. GIBSON, A. 1998. Photosynthesis organs of desert plants. Bioscience 48(11):911-920. [ Links ]
9. GROSSNICKLE, S. 2005. Importance of root growth in overcoming planting stress. New for. 30:273-294. [ Links ]
10. GUARNASCHELLI, A; P PRYSTUPA & J LEMCOFF. 2006. Drought conditioning improves water status, stomatal conductance and survival fo Eucalyptus globulus subspecies bicostata seedlings. Ann For. Sci. 63:941-950. [ Links ]
11. GUPTA, G. 1995. Rain-water management for tree planting in the Indian Desert. J Arid. Environ. 31:219-235. [ Links ]
12. HIJMANS, RJ; S CAMERON; J PARRA; P JONES & JARVIS. 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J Climatol. 25:1965-1978. [ Links ]
13. INFOSTAT. 2009. InfoStat versión 2009. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina [ Links ]
14. IPCC. 2007. Cambio climático 2007: Informe de síntesis. Pp. 52 en: Cuarto Informe de evaluación del Grupo Intergubernamental de Expertos sobre el Cambio Climático [Contribución de los Grupos de trabajo I, II y III. Equipo de redacción principal: Pachauri, RK y A Reisinger (directores de la publicación)]. IPCC, Ginebra, Suiza. [ Links ]
15. KARLIN, U. 1983. Recursos forrajeros naturales del Chaco Seco: Manejo de Leñosas. Pp. 78-96 en: ll Reunión de Intercambio Tecnológico en Zonas Áridas y Semiáridas. Córdoba. [ Links ]
16. KARLIN, U; R COIRINI; L PIETRARELLI & E PERPIÑAL. 1992. Caracterización del Chaco Árido y propuesta de recuperación del recurso forestal. Pp. 7-12 en: Karlin, UO & RO Coirini (eds.). Sistemas agroforestales para pequeños productores de zonas áridas. Proyecto GTZ. Desarrollo agroforestal en comunidades rurales del Noroeste Argentino, Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. [ Links ]
17. MONTAGNINI, F; M GUARIGUATA-URBANO; A MARISCAL-CHAVEZ; N RIBEIRO & D SHEPHERD. 1999. Reforestación con especies nativas para la recuperación de áreas degradadas: experiencias en tres regiones de Latinoamérica. Pp. 07-14 en: Seminario Centroamericano La Reforestación: Una Alternativa Social, Económica y Ambiental Ante los Desastres Naturales. I., Siguatepeque, HN, 14-16 de julio, 1999. [ Links ]
18. MORELLO, J; J PROTOMASTRO; L SANCHOLUZ & C BLANCO. 1985. Estudio macroecológico de los Llanos de La Rioja. Serie del Cincuentenario de la Administración de Parques Nacionales 5:1-53. [ Links ]
19. PALLARDY, S & N LOEWENSTEIN. 2004. The role of xylem sap abscisic acid in leaf abscission of droughted seedlings of isohydric and anisohydric temperate deciduous angiosperms. Pp. 365-372 en: Proceedings of the 14th Central Hardwoods Forest Conference GTR-NE. [ Links ]
20. PERPIÑAN, E & L PIETRARELLI. 1992. Variabilidad de la densidad y contracción volumétrica de la madera de Prosopis. Invs. Agr. Sist. Rec. For. 1(2)-223-221. [ Links ]
21. PRATALONGO, P; R QUINTANA; I MALVÁRES & M CAGNONI. 2003. Comparative analysis of variables associates with germination and seedling establishment for Prosopis nigra (Griseb.) Hieron and Acacia caven (Mol.). For. Ecol. Manag. 179:15-25. [ Links ]
22. VALLADARES, F; D SÁNCHEZ-GÓMEZ & M ZAVALA. 2006. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 94:1103-1116. [ Links ]
23. VERGA, A. 1995. Genetic study of Prosopis chilensis y Prosopis flexuosa (Mimosaceae) in the dry Chaco of Argentina. Tesis Doctoral. Göttingen Researh. Notes in Forest Genetics. Abteilung für Forstgenetik und Forstpflanzensüchtung der Universität Göttingen. Göttingen, Alemania. Pp. 96. [ Links ]
24. VERGA, A. 2000a. Algarrobos como especies para forestación: una estrategia de mejoramiento. SAGPyA Forestal, Nº 16 Septiembre de 2000 y Nº 17 octubre de 2000. [ Links ]
25. VERGA, A. 2000b. Hojas. Programa para la medición de caracteres morfológicos de hojas. INTA. IFFVE. Córdoba. Argentina. [ Links ]
26. VERGA, A & H GREGORIUS. 2007. Comparing morphological with genetic distances between populations: A new method and its application to the Prosopis chilensis - P. flexuosa complex. Silvae Genet. 56(2):45-51. [ Links ]
27. VILAGROSA, A; J BELLOT; V VALLEJO & E GIL-PELEGRÍN. 2003. Cavitation, stomatal conductance, and leaf dieback in seedlings of two co-occurring mediterranean shrubs during an intense drought. J. Exp. Bot. 54(390):2015-2024. [ Links ]
28. VILLAGRA, P & J CAVAGNARO. 2005. Water stress effects on the seedlings growth of Prosopis argentina and Prosopis alpataco. J. Arid Environ. 64(3):390-400. [ Links ]
29. VILLAGRA, P; A VILELA; C GIORDANO & J ÁLVAREZ. 2010. Ecophysiology of Prosopis species from the arid lands of Argentina: What do we know about adaptation to stressful environments? Desert Plants 3:321-340. [ Links ]
30. VILLAR-SALVADOR, P; R PLANELLES; J OLIET; J PEÑUELAS-RUBIRA; D JACOBS; ET AL. 2004. Drought tolerance and transplanting performance of holm oak (Quercus ilex) seedlings after drought hardening in the nursery. Tree Physiol. 24:1147-1155. [ Links ]
31. VILELA, A & R PALACIOS. 1997. Adaptative features in leaves of South American species of the genus Prosopis (Leguminosae; Mimosoideae). Bulletin of the International Group for Study of Mimosoideae 20:62-70. [ Links ]

