










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.23 no.2 Córdoba ago. 2013
ARTÍCULO ORIGINAL
Rendimiento hídrico en cuencas primarias bajo pastizales y plantaciones de pino de las sierras de Córdoba (Argentina)
Esteban G. Jobbágy1,2, *; Ana M. Acosta1 & Marcelo D. Nosetto1,2,3
1. Grupo de Estudios Ambientales, IMASL - Universidad Nacional de San Luis y CONICET.
2. Departamento de Agronomía - FICES, Universidad Nacional de San Luis, Argentina.
3. Cátedra de Climatología Agrícola. Facultad de Ciencias Agropecuarias. Universidad Nacional de Entre Ríos, Argentina.
* jobbagy@unsl.edu.ar
Editora Asociada: Ana Cingolani
Recibido: 17 de enero de 2013; Fin de arbitraje: 8 de marzo;
Revisión recibida: 28 de marzo; Aceptado: 27 de abril.
RESUMEN
Los cambios en la cobertura vegetal pueden tener importantes efectos sobre el ciclo hidrológico, afectando la magnitud y distribución temporal del caudal de ríos y arroyos. Estos efectos cobran una relevancia especial en áreas montañosas de regiones secas dado su papel importante en la provisión de agua. Este es el caso de las laderas orientales de las sierras de Córdoba, en donde se establecieron ~35000 ha de plantaciones de pinos en reemplazo de pastizales naturales. Exploramos cómo esta transformación ha afectado el rendimiento hídrico de pequeñas cuencas serranas. Para ello seleccionamos cuatro pares de cuencas primarias ocupadas por pastizales naturales y plantaciones de Pinus ellioti (superficie: 27 a 143 ha; elevación: 1100 a 1750 m.s.n.m.). En todos estos pares de cuencas determinamos el caudal base de arroyos por dilución de un trazador salino con frecuencia estacional entre mayo de 2004 y enero de 2007, y en dos de ellos realizamos un seguimiento continuo del caudal con sensores automáticos durante la transición entre la estación seca y húmeda de 2006-2007. En promedio, el rendimiento hídrico de las cuencas forestadas fue 48% inferior al de las cuencas de pastizal (112 vs. 204 mm/año o 24 vs. 13% de la precipitación recibida, P<0.05). Las mediciones continuas mostraron caudales base mayores y poca respuesta al tipo de vegetación en cuencas elevadas y de alta pendiente y caudales base menores y muy sensibles al establecimiento de plantaciones en cuencas de menor elevación y pendiente intermedia. La caracterización satelital de todas las cuencas a partir del índice verde normalizado (NDVI) satelital del sensor MODIS sugirió una productividad primaria y evapotranspiración mayor y más estable bajo plantaciones respecto a los pastizales, con contrastes máximos entre estos tipos de vegetación observados en invierno. En la actualidad, los impactos de las plantaciones serranas sobre la provisión de agua pueden manifestarse únicamente en cuencas primarias dado que las de mayor orden se encuentran forestadas sólo de manera parcial. A nivel regional, es importante contemplar estos efectos al proyectar cuánto, dónde y cómo se forestará, en especial en áreas que aportan agua a los principales focos de consumo energético y urbano.
Palabras clave: Forestación; Ciclo del agua; Servicios ecosistémicos; Balance hídrico
ABSTRACT
Water yield in primary watersheds under grasslands and pine plantations in the hills of Córdoba (Argentina): Vegetation changes can have a strong imprint on the hydrological cycle, affecting the magnitude and temporal distribution of stream and river flow. These effects gain special relevance in the mountain ranges of dry regions, which play a key role in water provision. This is the case of the eastern front of the Córdoba hills, where ~35.000 ha of pine plantations have replaced natural grasslands. We explored how this transformation has affected water yield in small watersheds. For this purpose we selected four pairs of neighboring watersheds occupied by natural grasslands and Pinus ellioti plantations (area: 27 to 143 ha; elevation: 1100 to 1750 m.a.s.l.). For all pairs, basal streamflow was measured through the tracer dilution method with seasonal frequency between May 2004 and January 2007, and in two of these pairs we performed a continuous monitoring using automatic level sensors during the transition between the dry and humid season of 2006-2007. On average, water yield in afforested watersheds was 48% less than in grassland watersheds (112 vs. 204 mm/year or 24 vs. 13% of precipitation inputs during the study period, P<0.05). Continuous measurements revealed higher base flows and little response to vegetation in high elevation and slope watersheds, and lower and more vegetationsensitive base flows in low elevation and intermediate slope watersheds. The remote sensing characterization of watersheds based on a green index (NDVI) from MODIS suggested higher and more stable primary productivity and evapotranspiration under plantations compared to grasslands, with maximum contrasts taking place in winter. Currently the impact of plantations in the hills on water provision can be intense only at the level of primary watersheds, since those of larger order are only partially afforested. At the regional level is important to contemplate these effects to project how much, where and how will be planted, particularly in areas that supply water to the largest foci of hydroenergy and urban demand.
Keywords: Afforestation; Water cycle; Ecosystem services; Water balance
INTRODUCCIÓN
En regiones con balances hídricos negativos (evapotranspiración potencial> precipitación), las áreas montañosas juegan un papel clave como proveedoras de agua líquida (Viviroli & Weingartner 2004). Bajo este contexto climático, los territorios llanos y sedimentarios devuelven casi la totalidad de la precipitación que reciben a la atmósfera por transpiración o evaporación directa (Scanlon et al. 2006; Jobbágy et al. 2008), mientras que en los sectores montañosos o serranos las altas pendientes y la rocosidad suelen impedir este consumo evaporativo exhaustivo, favoreciendo la alimentación de cursos de agua tanto por escurrimiento (transporte superficial) como por drenaje profundo (transporte subterráneo), aún bajo climas áridos (Jobbágy et al. 2011).
El rendimiento hídrico de las cuencas montañosas, definido como la fracción de la precipitación que abandona las mismas en forma líquida, no sólo depende del clima y de la configuración topográfica y geológica de las cuencas, sino también de la vegetación que las ocupa (Zhang et al. 2001; Brown et al. 2005; Huxman et al. 2005; Jackson et al. 2009; Nosetto et al. 2012). Desde hace casi un siglo, se han documentado respuestas del caudal de cuencas montañosas tras cambios en la vegetación, incluyendo alteraciones en el caudal total anual, en su estacionalidad, y en su partición entre "picos de caudal" o crecientes inmediatamente posteriores a las lluvias y "caudal base" asociado al aporte más lento del sistema hidrológico subterráneo (Farley et al. 2005). Las transformaciones hidrológicas de una cuenca no solo afectan la provisión de agua para consumo humano, industrial, y agrícola, sino también la regulación de crecientes e inundaciones (Jackson et al. 2001; Bradshaw et al. 2007). Desde el punto de vista de la provisión de agua, los cambios en el caudal base pueden ser más importantes que los del caudal total, siendo el primero el que define la oferta segura o estable de agua para sus distintos usos, mientras que desde el punto de vista de la regulación de crecientes, los picos de caudal son el componente más relevante.
Entre los cambios posibles de vegetación, aquellos que involucran transiciones de sistemas herbáceos a leñosos, suelen tener las improntas hídricas más significativas (Zhang et al. 2001; Brown et al. 2005; van Dijk & Keenan 2007). El establecimiento de forestaciones en sistemas dominados por pastos puede generar reducciones en el rendimiento hídrico de las cuencas como resultado de aumentos en la evapotranspiración (Zhang et al. 2001), asociados tanto a su mayor capacidad de consumo evaporativo de agua (canopeos de mayor altura y conductancia aerodinámica, estación de crecimiento más prolongada) (Kelliher et al. 1993), como a la mayor capacidad de acceder al agua almacenada en el suelo-roca gracias a sus sistemas radicales más extensos (Schenk & Jackson 2002).
Una revisión global de estudios hidrológicos comparativos de pequeñas cuencas pareadas de pastizales forestados con especies de rápido crecimiento (pinos y eucaliptos) y controles no forestados mostró una reducción generalizada del rendimiento hídrico tras la forestación (Farley et al. 2005). En promedio, las forestaciones redujeron las salidas absolutas de agua en un 39 %, con efectos crecientes hacia climas más secos y más notables para el caudal base que para el total. El resultado del establecimiento de plantaciones de pinos y eucaliptos en pastizales sudafricanos que reciben menos de 1000 mm/año de precipitación ha sido especialmente impactante: en esos pastizales se han registrado reducciones completas del caudal de arroyos (Scott & Lesch 1997).
Se han documentado situaciones en las que la forestación tiene efectos positivos sobre la provisión de agua, en general en cuencas que fueron forestadas tras sufrir degradación por sobrepastoreo y/o laboreo del suelo (Ilstedt et al. 2007). En estas condiciones, la forestación puede favorecer la infiltración desviando agua que se perdería por escorrentía, contribuyendo a los picos de caudal, hacia el flujo subterráneo y caudal base de la cuenca. En estos casos el rendimiento hídrico total no aumenta (de hecho por lo general disminuye) pero se logra una oferta más estable de agua que evita crecientes y eventos de erosión y, en menor cantidad de situaciones, eleva el caudal base (Ilstedt et al. 2007).
El sistema montañoso de las Sierras Grandes (macizo de Comechingones, Córdoba, Argentina), de sentido norte-sur, está dominado en sus zonas más elevadas (>1200 m) por pastizales en los que tienen su origen arroyos y ríos que alcanzan la llanura pampeana hacia el este o que infiltran en el terreno sedimentario de los valles interserranos hacia el oeste. En particular dos de estos ríos, el Suquía y el Xanáes, proveen agua a gran parte del área urbana e industrial de la ciudad de Córdoba con 3 millones de habitantes (Censo 2010, www.indec.gov.ar). Si bien las primeras plantaciones forestales serranas se establecieron a fines de la década del cincuenta, en la de los setenta se expandieron más rápidamente motivadas por políticas provinciales de promoción forestal, alcanzando tasas máximas de implantación en 1976 (Izurieta et al. 1993). Las especies implantadas fueron Pinus radiata al comienzo y luego, por su mejor crecimiento y sanidad, P. elliottii y, en menor medida, P. taeda (Dorado et al. 1997). La mayoría de las plantaciones no han recibido podas ni raleos durante su ciclo y hoy en día se explotan en forma parcial tras décadas sin manejo. En la última década se observa la invasión espontánea de pinos en el pastizal adyacente a las plantaciones, siguiendo tendencias descriptas en otras regiones de pastizales y arbustales forestadas con pinos (Richardson 1998).
El objetivo general de este estudio es explorar los efectos hídricos de forestaciones de pino establecidas sobre pastizales, con especial atención al rendimiento hídrico de cuencas serranas. Para ello, en la ladera oriental de las Sierras Grandes se identificaron pequeñas cuencas pareadas (pastizal-plantación) en las que se buscó caracterizar los efectos del establecimiento de plantaciones de Pinus elliottii Engelm. sobre i) la magnitud y variabilidad del caudal base (mediciones estacionales durante tres años), ii) la capacidad evapotranspirativa del canopeo (estimaciones satelitales de índice verde durante el mismo período) y iii) dinámica diaria del caudal total (mediciones continuas durante la transición de una estación seca a la húmeda siguiente).
MATERIALES Y MÉTODOS
La zona de estudio se ubica en el valle de Calamuchita, en la ladera oriental de las Sierras Grandes de Córdoba en Argentina (Figura 1). La precipitación media anual es de 850 mm, concentrada principalmente entre los meses de octubre y abril, y la evapotranspiración potencial alcanza los 1200 mm/año. La temperatura media anual decrece desde los 13.1 a 9.7 ºC al ascender desde 1000 a 1800 m de elevación (Cabido et al. 1997). La litología está dominada por rocas metamórficas (gneis y anatexitas ultrametamórficas, basamento pre-devónico) que suelen presentar relieves homogéneos y abundantes afloramientos (Bonalumi et al. 1999). Una fracción minoritaria de las laderas, en sus sectores más planos o pampas, está ocupado por sedimentos cuaternarios (Acosta et al. 1992). Los suelos son predominantemente someros con 10 a 60 cm de desarrollo hasta la roca y estratos de carbonato de calcio cuando son más profundos. Se encuentra una transición desde Entisoles a Haplumbrempts a Hapludoles y Argiudoles desde las zonas más rocosas a las pampas, mientras que las zonas más bajas y húmedas albergan Haplacuoles (Farley et al. 2009). La cobertura vegetal natural dominante en la actualidad es la de pastizales, compuestos por una mezcla de especies gramíneas C3 y C4, cuya composición relativa cambia de forma abrupta con la elevación, a favor de las especies C3 que, en la faja altitudinal en la que se llevó a cabo este estudio, pasan de ser co-dominantes a dominantes (Cabido et al. 1997). Estos pastizales se encuentran sujetos a pastoreo extensivo por ganado bovino y en menor medida caprino. En la zona de estudio se han establecido parcelas de plantaciones de pinos, principalmente de Pinus ellioti. Las mismas fueron establecidas entre fines de los 70s y comienzos de los 80s, ~25 años antes de la realización de este estudio, con densidades de plantación elevadas (>1000 plantas/ha) y no han recibido tratamientos de poda o raleo, observándose raleo espontáneo únicamente. En la última década varias de estas plantaciones han sido cosechadas con fines comerciales por primera vez.
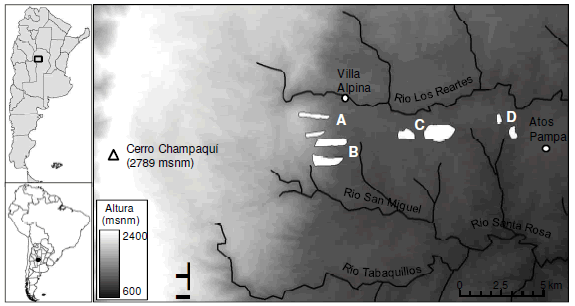
Figura 1. Ubicación del área de estudio y disposición de las cuencas monitoreadas. Cada letra indica un sitio que incluye un par de cuencas vecinas bajo pastizal y plantación de pinos. La elevación del terreno se indica en tonos de gris. Los ríos de mayor magnitud se indican con líneas negras y los poblados con círculos.
Figure 1. Location of the study area and distribution of the monitored watersheds. Each letter indicates a site that includes a pair of neighboring watersheds under grassland and pine plantation. Elevation is shown in grey scale. Major rivers are indicated with black lines and towns with circles.
Sobre el extremo norte de la cuenca del Río Tercero se seleccionaron cuatro pares de microcuencas cubiertas por pastizales y plantaciones de Pinus ellioti (superficie: 27 a 143 ha; elevación: 1097 a 1728 m.s.n.m.). Los pares de cuencas corresponden a los sitios denominados A, B, C y D y alojan arroyos permanentes de primer orden (Tabla 1, Figura 1). Se buscó que en cada sitio las condiciones topográficas y geológicas de las cuencas bajo pastizal y plantación fueran similares, siendo la variación entre sitios mayor que la variación entre cuencas de un mismo sitio.
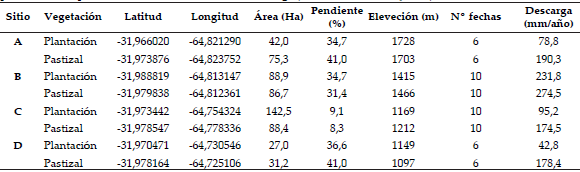
Tabla 1. Caracterización de las cuencas estudiadas incluyendo localización geográfica, área aguas arriba del punto de muestreo, pendiente y elevación media y valores medios de descarga base (caudal base relativo al área de la cuenca).
Table 1. Characterization of the studied watersheds including geographic location, area upstream of the sampling point, mean slope and elevation, and mean base discharge (base flow divided by area).
Las observaciones realizadas incluyeron (i) mediciones a campo de caudal base (ii) mediciones satelitales de índice verde (mayo de 2004 a febrero de 2007) en los cuatro pares de cuencas y (iii) mediciones continuas de caudal en dos pares de cuencas (octubre de 2006 a febrero de 2007). Se midió el caudal base en los cuatro pares de arroyos de manera repetida en seis fechas entre mayo de 2004 y agosto de 2005 y en dos pares de cuencas se continuaron las mediciones agregando cuatro fechas hasta febrero de 2007. Estas mediciones se realizaron por el método de dilución de sales (Day 1976), que consiste en la inyección, a flujo constante de una solución salina (5 mL/s, 300 g NaCl/L durante 5 minutos). A partir de la medición de conductividad eléctrica (Orion Model 115 - Thermo Fisher Scientific Inc.) del curso de agua antes, durante y después de la inyección y de la solución salina diluida con agua del arroyo en factores de 1:102 a 1:105 se calculó el caudal del curso de agua usando la técnica del "plateau" (a partir de la conductividad eléctrica máxima y estable alcanzada durante la inyección) y de la integral (a partir del exceso de conductividad eléctrica generado integrado durante e inmediatamente después de la inyección) (Day 1976). Ambas técnicas arrojaron resultados similares (<5% de diferencia; se eligió el método de la integral). Estas técnicas son en especial adecuadas para cursos de bajo caudal y se aplicaron en segmentos de alta velocidad y bajo volumen de todos los arroyos. En dos fechas este método fue comparado con el de medición de velocidad y sección usando un sensor doppler (Flo-Mate Model 2000, Marsh-McBirney, Inc.) hallándose un buen acuerdo (R2=0.982, n=16).
Las mismas cuencas también se caracterizaron satelitalmente a partir del índice verde normalizado (NDVI) del sensor MODIS (producto MOD13Q1, grilla de 250 m de resolución espacial, compuestos de 16 días de resolución temporal), el cual ha sido utilizado como indicador de la magnitud y estacionalidad de la productividad primaria y de la evapotranspiración en otras regiones del centro de Argentina (Contreras et al. 2011; Vassallo et al. en prensa). Las microcuencas se definieron como sitios de muestreo y de ellas se extrajeron los valores correspondientes a los píxeles completamente ocupados por pastizal o plantaciones, según el caso, cubriéndose el período de este estudio (2004-2007). Los datos de precipitaciones del aérea de estudio para todo el período considerado se obtuvieron a partir de TRMM (The Tropical Rainfall Measuring Mission), que aporta estimaciones globales con una frecuencia temporal de tres horas mostrando una buena correlación con los valores tomados en el terreno en regiones vecinas (Aragón et al. 2010). En este trabajo se emplearon los valores acumulados mensuales para el período de estudio.
En dos de los cuatro pares de cuencas se realizaron mediciones continuas de nivel entre Octubre de 2006 y Febrero de 2007 y en el mismo período se estableció a 2 km del sitio donde se ubicaron las cuencas del par B una estación meteorológica automatizada (Vantage Pro2, Davis Inc.) que registró cada 10 minutos la precipitación acumulada, temperatura y humedad del aire. Para las determinaciones de caudal se utilizaron transductores de presión (Levelogger, Solinst Inc.) para medir el nivel de agua (resolución temporal 10 minutos y vertical de ±2.5 mm) instalados en pozones cuya salida natural estaba constituida por una garganta de roca sólida y estable. En estos puntos se realizó una calibración de la relación caudal vs. nivel, medido con la técnica de dilución de solución salina descripta anteriormente en cuatro fechas en las que el nivel fue contrastante, lográndose ajustes aceptables con funciones exponenciales (R2 0.991 a 0.997) (Hauer & Lamberti 1996). A partir de estas funciones se calcularon los valores de caudal por interpolación. En aquellas fechas en que los niveles medidos superaron los valores máximos de la calibración, correspondientes en general a crecientes, los caudales no pudieron determinarse con certeza y fueron extrapolados, por lo que sus estimaciones absolutas deben considerarse como tentativas y útiles principalmente para comprender la dinámica temporal de estos eventos extremos de caudal. Las series de caudal continuo obtenidas de esta forma fueron sometidas a un análisis de separación de hidrógrafo con el fin de determinar la contribución del caudal base y pico al total (Arnold & Allen 1999). Si bien este análisis puede realizarse en forma manual sobre gráficas de caudal total, se prefirió utilizar un filtro digital recursivo de las series que consiste en la aplicación de un algoritmo de separación de componente (base y pico) que puede operar en ambos sentidos (hacia adelante y atrás en el tiempo) hasta mostrar una correcta separación gráfica del caudal base (Arnold et al. 1995). Se aplicó este algoritmo a los datos medios diarios de caudal usando tres pasos (adelante, atrás, adelante)(Arnold et al. 1995), encontrándose una reducción del caudal base estimada <1% entre el primer y segundo paso y del 7 a 17% entre el segundo y el tercero. Se prefirió el tercer paso por mostrar una mejor separación gráfica de picos de caudal en todas las cuencas. Para todas las medidas de caudal tanto continuas como puntuales se realizó la conversión a descarga dividiendo los valores obtenidos por la superficie de la cuenca y expresándolos en términos de lámina media por unidad de tiempo.
Para la información de caudal y satelital obtenida en múltiples fechas para el conjunto total de pares de cuencas (n=4) se aplicó un análisis de varianza de dos factores que incluyeron a la vegetación (pastizal vs. plantación) y al sitio de localización del par de cuencas (A,B,C, y D, Tabla 1). Se consideró como factor aleatorio a la fecha con el fin de eliminar la contribución importante que introdujo la variación temporal del sistema a los efectos del tipo de vegetación que se querían evaluar. Los valores de descarga media según tipo de vegetación fueron comparados aplicando una prueba t de Student para casos pareados. Para las series continuas de caudal en las que se compararon solamente dos pares de cuencas se realizaron únicamente análisis descriptivos.
RESULTADOS
La descarga hídrica de las plantaciones forestales fue menor a la de sus pastizales vecinos. De las 32 observaciones pareadas de caudal base transformadas en descarga diaria que se realizaron, 71.8% mostró valores más altos en las cuencas bajo pastizal respecto a los de aquellas ocupadas por plantaciones (Figura 2). La comparación de estos valores para todos los pares de cuencas aplicando el ANOVA con factor aleatorio fecha (6 fechas para las que se contaba con mediciones en todas las cuencas) y factores vegetación (pastizal/ plantación) y sitio (A,B,C y D) mostró un efecto significativo de la vegetación (P=0.0014) y del sitio (P<0.001), mientras que su interacción resultó marginalmente significativa (P=0.072). La descarga anual media del flujo base de las cuencas forestadas fue poco más de la mitad de aquellas cubiertas por pastizal (112 vs. 204 mm/año, P<0.05 según prueba t pareada, Tabla 1), representando respectivamente un 13 y 24% de la precipitación total recibida en el período de estudio (años 2004-2006, 853 mm/año). Las descargas base medias fueron muy variables entre sitios, especialmente para las cuencas cubiertas por plantaciones (Tabla 1). El sitio B, ubicado en una de las laderas más rocosas del área estudiada, mostró los rendimientos máximos y las menores diferencias entre tipos de vegetación mientras que los sitios C y D, ubicados a menor altitud y en zonas con menor superficie de roca expuesta mostraron los rendimientos hídricos menores bajo pastizal y las diferencias mayores entre tipos de vegetación (Tabla 1).
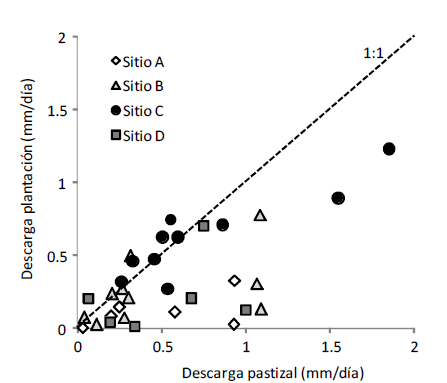
Figura 2. Comparación de la descarga hídrica de cuencas pareadas de plantaciones de pino y pastizales. Cada punto representa el par de valores de descarga base (caudal base relativo al área) de cuencas vecinas ocupadas por plantaciones y por pastizales para 6 a 10 fechas en cuatro sitios de estudio.
Figure 2. Comparison of hydrological discharges in paired watersheds occupied by pine plantations and grasslands. Each data point represents a pair of basal discharge values (basal flow divided by area) for neighboring watersheds occupied by plantations and grasslands for 6 to 10 sampling dates in four study sites.
El período de observaciones de casi tres años abarcó tres estaciones secas completas. El caudal base a lo largo de este período mostró fuertes fluctuaciones con mayores diferencias entre máximos y mínimos para pastizales que plantaciones (Figura 3). En la primavera de 2004 y en el invierno-primavera de 2006 se detectaron los caudales mínimos en pastizales y plantaciones. En este último período la cuenca bajo plantación del sitio C llegó interrumpir completamente su flujo superficial por al menos un mes (Figura 4). Los valores máximos de caudal base en pastizales tuvieron lugar en verano de 2006 (estación húmeda más lluviosa del período) cuando llegaron a sextuplicar el caudal base de las plantaciones. En los otoños se encontró mayor similitud de caudales entre ambos tipos de vegetación (Figura 3).
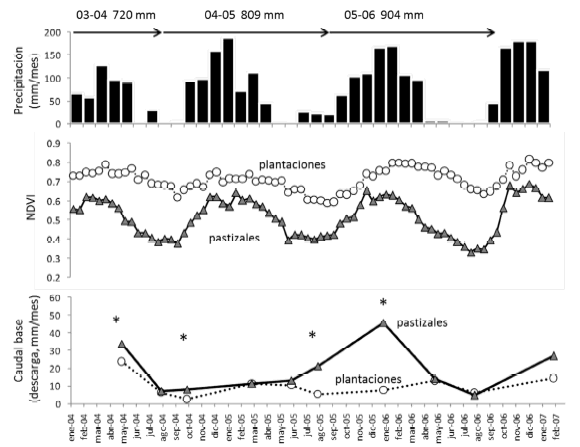
Figura 3. Series temporales (2004-2007) de precipitación, índice verde satelital (NDVI) y caudal base para las cuencas estudiadas. El panel superior corresponde a valores mensuales de precipitación obtenidos del sistema TRMM (barras) y los acumulados de cada año hidrológico (septiembre a agosto, flechas horizontales). El panel central ilustra los valores medios de NDVI correspondientes a las cuencas de los cuatro sitios de estudio. El panel inferior muestra los caudales base medios, expresados como descarga mensual (caudal base relativo al área) de las cuencas bajo plantación y pastizal en cuatro sitios de estudio para las primeras seis fechas y en dos sitios para las siguientes cuatro fechas. Los asteriscos indican diferencias significativas entre tipos de vegetación (P<0.05, prueba t pareada) para cada fecha individual.
Figure 3. Temporal series (2004-2007) of precipitation, vegetation greenness and basal stream flow for the studied watersheds. See details in legend in Spanish.
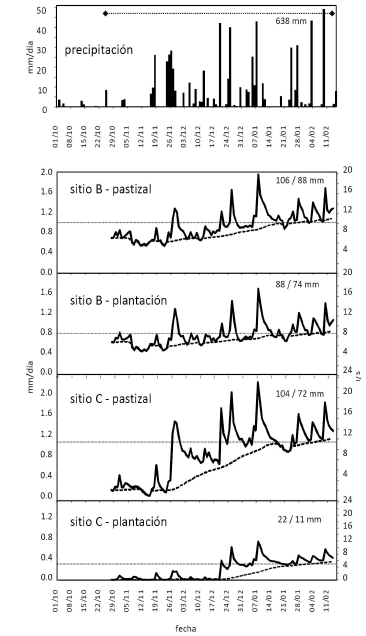
Figura 4. Serie de alta resolución temporal de precipitación y caudales. Los valores de precipitación diaria (barras) obtenidos con una estación automática en el sitio B muestran encima el valor acumulado desde el comienzo de las mediciones de caudal. Los valores de caudal se expresan como tales (eje Y derecho) y como descarga diaria (eje Y izquierdo) y separan valores totales (línea llena) y base (línea punteada) estimado según un algoritmo de filtrado (Arnold & Allen 1999). Las líneas horizontales muestran el valor máximo de calibración caudal-nivel, por encima del cual las medidas tienen mayor incertidumbre. Los valores integrados de descarga total y base se indican arriba y a la derecha de cada panel.
Figure 4. High temporal resolution series of precipitation and flow data. See details in legend in Spanish.
Las diferencias hidrológicas entre tipos de vegetación tuvieron su contraparte en la actividad de fotosintética y transpirativa de los canopeos, capturada en forma indirecta por el NDVI satelital. Este índice alcanzó en todo el período valores más altos en las plantaciones que en los pastizales siendo más fluctuante en estos últimos. Las diferencias más notables entre pastizales y plantaciones tuvieron lugar en la estación seca (Figura 3). Es interesante destacar como tras el intervalo seco más extendido del período estudiado (mayo a septiembre de 2006, precipitación total 22 mm), los pastizales vieron descender su NDVI al mínimo registrado, mientras que las plantaciones mantuvieron valores similares o mayores que los registrados en los mismos meses de años anteriores. Esto sugiere que las plantaciones disponen de reservas de agua mayores por explorar una mayor profundidad de suelo que los pastizales. Por otra parte, la recuperación del NDVI en pastizales tras el reinicio de las lluvias en la primavera de 2006 fue muy veloz y para fines del período de estudio ambos tipos de vegetación alcanzaron su NDVI máximo (Figura 3). A pesar de las diferencias de NDVI observadas en la estación húmeda de 2004-2005, los caudales fueron igualmente bajos para ambos tipos de vegetación en esa temporada, lo que alerta sobre el hecho de que a igualdad de consumo de agua, las forestaciones podrían sostener niveles de NDVI más altos.
Las mediciones continuas de caudal se concentraron en el período de transición entre una estación seca particularmente extendida y una húmeda con precipitaciones intensas (Figuras 3 y 4). Se eligieron dos sitios contrastantes, uno con alta pendiente y rocosidad y descargas altas (sitio B) y otro de relieve más plano, baja rocosidad y descargas menores (sitio C). El algoritmo utilizado permitió separar gráficamente la contribución relativa de los caudales base y pico al total si bien las estimaciones de caudal pico y total hacia el final del período resultó más incierta por haberse superado el rango de calibración nivel-caudal de los arroyos estudiados (líneas punteadas horizontales en Figura 4). En el sitio B, las diferencias en la magnitud y distribución temporal de la descarga entre tipos de vegetación fueron pequeñas, con ascensos graduales de caudal base a partir de las lluvias intensas de fin de noviembre. En estas dos cuencas el caudal base sólo se duplicó desde la estación seca a la húmeda y representó 83-84% del caudal total, sugiriendo una fuerte contribución del flujo subterráneo a la alimentación de estas cuencas independientemente del tipo de vegetación (Figura 4). En el sitio C, los contrastes entre tipos de vegetación y entre la estación seca y húmeda fueron mucho mayores. Mientras que en la cuenca de pastizal se mantuvo un caudal base bajo pero sostenido durante la estación seca, en la ocupada por la plantación se interrumpió por completo el flujo de agua en algún momento entre la medición previa de agosto de 2006 y el inicio de las mediciones continuas. Al comenzar las lluvias el caudal base se elevó rápidamente en el cuenca de pastizal mientras que en la de plantación se observaron solamente pequeños episodios de caudal pico y solamente a fin de diciembre de 2006 se reanudó el flujo base (Figura 4). En todo el período de mediciones continuas el caudal total de la plantación del sitio C fue tan solo una quinta parte del medido en la cuenca vecina de pastizal. Por otra parte, mientras que en la cuenca forestada el 50% de ese flujo fue caudal base, en la de pastizal este valor alcanzó el 69%. Las mediciones del sitio alto (B) vs. bajo (C) sugieren un impacto mucho más exacerbado de las plantaciones en cuencas más planas y menos rocosas, posiblemente explicado por un mejor acceso a fuentes profundas de agua por parte de los árboles, que permiten interceptar agua que escaparía del alcance de las raíces de pastizal y retrasar la llegada de excedentes hídricos a cursos superficiales. Al margen del efecto de la vegetación, son especialmente relevantes las diferencias generales halladas en el comportamiento de las cuencas correspondientes al sitio alto y bajo, que sugieren una fuerte influencia de la condición geológica de las cuencas. Los elevados caudales base de las cuencas del sitio alto indicarían que la alta rocosidad de esta cuenca habría favorecido el ingreso rápido de una fracción importante de las precipitaciones al sistema subterráneo de circulación lenta, que quedó fuera del alcance de las raíces tanto del pastizal como de la plantación y alcanzó los cursos de agua con un largo retraso. Si bien las mediciones de caudales máximos escaparon a la zona de calibración del estudio, las mismas sugieren que en las cuencas del sitio alto la forestación no amortiguaría picos de creciente, mientras que en las del sitio bajo lo harían en forma muy efectiva (Figura 4).
DISCUSIÓN
Bajo una vegetación actual dominada por pastos nativos, las cuencas de elevación intermedia de las Sierras Grandes de Córdoba que se estudiaron mantienen una descarga hídrica base de ~200 mm/año, equivalente a un cuarto de los aportes de la precipitación. Los restantes tres cuartos de la precipitación de estas cuencas cubiertas por pastizales se perderían por evapotranspiración y picos de creciente. El establecimiento de plantaciones de pino de alta densidad en este territorio ha reducido la descarga en forma variable según las condiciones de cada cuenca, promediando una contracción del caudal base cercana a 50% (Tabla 1). Cabe aclarar que bajo condiciones semiáridas o subhúmedas como las de la zona de estudio se requiere solamente un pequeño aumento de la evapotranspiración para producir un efecto tan importante sobre la descarga dado que naturalmente la misma representa una fracción minoritaria de los aportes de precipitación (Jobbágy et al. 2008). Los efectos observados coinciden en magnitud con los reportados para estudios de más largo plazo en cuencas montañosas de pastizales y arbustales que fueron forestadas en otras regiones del mundo (Farley et al. 2005) y destacan el papel dominante de la vegetación como control hidrológico en climas subhúmedos.
Los mecanismos que pueden llevar a las plantaciones a evapotranspirar más que los pastizales y a reducir significativamente el caudal base incluyen la capacidad de consumo y de acceso al agua (Calder 1998), siendo lo primero determinante en períodos/lugares húmedos y lo segundo en períodos/lugares secos. La alta capacidad de interceptar y transpirar agua es favorecida por la altura y rugosidad de los canopeos forestales, por su menor albedo y por su mayor área foliar, evidenciada en los sitios de estudio por los datos satelitales (Figura 3) (Kelliher et al. 1993; Nosetto et al. 2005). La posibilidad de acceso a fuentes de agua más profundas que las alcanzadas por la vegetación herbácea en el continuo suelo-roca es propiciada por sistemas radicales de mayor extensión y capacidad de penetración en fisuras rocosas (Heilman et al. 2009). La interacción de estas dos vías de influencia hidrológica de las plantaciones con las condiciones topográficas, geológicas y climáticas de las cuencas puede explicar las diferencias observadas entre sitios. Las condiciones más secas y cálidas de los sitios de menor elevación resaltaría el efecto del mejor acceso al agua sobre el rendimiento hídrico (en estas condiciones el proceso evaporativo es más limitado por la oferta), generando un "vaciamiento de reservas profundas de agua" que solo una vez recuperado durante la estación lluviosa permite el restablecimiento de los caudales base altos (Figura 4). En las cuencas más húmedas y frías de los sitios de mayor elevación el incremento del consumo de agua estaría dictado más fuertemente por la capacidad de interceptación y transpiración (en estas condiciones el proceso evaporativo es más limitado por la demanda), siendo el excedente hídrico naturalmente más alto y la posibilidad de "escape" del agua líquida al acceso de la vegetación, mayor. Las plantaciones de pino respecto a los pastizales pueden estar incrementado la evapotranspiración por dos vías, una mayor capacidad evaporativa del canopeo y/o un mejor acceso al agua del suelo (Calder 1998). El segundo parece ser el mecanismo más importante explicando el menor rendimiento hídrico bajo plantaciones dado que las diferencias de su índice verde respecto al del pastizal fueron mayores en los sitios bajos que en los altos (NDVI de plantación vs. pastizal 75 y 45% mayor, respectivamente).
Si bien los impactos sobre los recursos hídricos que hemos descripto son intensos y pueden afectar la disponibilidad de agua para consumo humano y animal o generación eléctrica, lo hacen hoy sólo a una escala muy local, del orden de los cientos de hectáreas, que es la extensión típica de las áreas forestadas. Por otra parte, nuestros resultados sugieren que el contexto geológico y topográfico de las cuencas en las que se establecen plantaciones forestales puede definir fuertemente su impacto hídrico (e.g., sitio B vs. C). Este cuestión, aún poco clara, debería guiar futuros pasos de investigación y zonificación de la actividad forestal en sistemas montañosos semiáridos. Al trasladar este estudio a la escala de los grandes ríos serranos de Córdoba, debe tenerse en cuenta entonces que: i) actualmente una fracción minoritaria de sus cuencas está forestada, ii) que las mermas hídricas dependen del tipo de paisaje que se haya forestado, y iii) que el aporte de zonas más altas de la sierra, aún no forestadas, puede ser desproporcionadamente alto y por lo tanto determinante del comportamiento de los ríos que abastecen. Es importante, sin embargo destacar tres aspectos que merecen atención y monitoreo de las cuencas de pastizal que son forestadas. El primero está relacionado a la expansión exponencial de las poblaciones espontáneas de pinos (invasión) en el área estudiada (Richardson 1998; Giorgis et al. 2011), que puede llevar a extender el área afectada hídricamente en las próximas décadas. Las mermas de rendimiento hídrico causadas por la invasión de árboles han motivado acciones de remediación de gran escala en países semiáridos con altas limitaciones hídricas como Sudáfrica (Prinsloo & Scott 1999). En el área de estudio debe sumarse a este proceso de invasión el de otras especies exóticas acompañantes como la zarzamora (Rubus ulmifolia Schott) que forma macizos muy densos bajo plantación, especialmente en zonas riparias. El segundo aspecto que requiere atención es el de la cosecha forestal. Si bien los sistemas forestales reducen el caudal base y por lo general también el caudal pico, ambos efectos se invierten en forma breve durante los eventos de cosecha (Jones & Grant 1996). Esto resulta sin duda beneficioso desde el punto de vista del rendimiento hídrico, pero puede generar problemas con caudales pico máximos que superan los niveles experimentados bajo pastizales nativos. Las causas de esto son la reducción brusca de la evapotranspiración y el fuerte disturbio que genera el establecimiento de caminos de madereo en las cuencas (Jones & Grant 1996). En Córdoba el reciente avance de la cosecha y el efecto más devastador que pueden tener los incendios masivos de plantaciones requieren atención por su posible influencia sobre los niveles máximos de las crecientes. Finalmente, el establecimiento de plantaciones puede afectar ya no la cantidad y distribución de los recursos hídricos sino su calidad. Se han reportado en muchos sistemas dominados por pastizales que fueron forestados cambios en las condiciones químicas del agua, siendo la acidificación y aumento de la solubilidad de iones tóxicos como el aluminio uno de ellos. Sin embargo en las Sierras de Córdoba este efecto no se ha detectado hasta el presente. Un trabajo previo que detectó acidificación en cuencas forestadas de Uruguay, no la detectó en las cuencas pareadas de Córdoba que se analizaron aquí (Farley et al. 2009), indicando impactos mínimos sobre la calidad del agua.
Este estudio se ha concentrado en las situaciones de máximo impacto esperable de las plantaciones forestales en las Sierras Grandes de Córdoba al considerar pequeñas cuencas completamente plantadas y poblaciones de alta densidad de especies de rápido crecimiento. El avance de la actividad forestal en la región puede generar impactos menores a los observados si incorpora límites máximos del área a forestar en cada cuenca, favorece el establecimiento de especies de menor consumo de agua (e.g., caducifolias de menor crecimiento pero mayor valor forestal), emplea densidades de plantación menores, monitorea y controla en forma permanente la propogación espontánea de especies forestales, y profundiza el conocimiento de la interacción de la vegetación con las condiciones climáticas y geohidrológicas en el marco del ordenamiento territorial. En territorios en los que las cuencas aportan bienestar a los humanos no sólo generando productos agropecuarios y forestales sino también a través de la entrega de un recurso escaso y muy valioso como es el agua (especialmente en una región semiárida con centros urbanos muy poblados), resulta fundamental enmarcar la silvicultura y ordenamiento forestal en el contexto más amplio de los servicios de los ecosistemas (Laterra et al. 2012). Por otra parte se suma como factor adicional a la complejidad de la vegetación y su relación con los recursos hídricos el hecho de que la faja altitudinal que se estudió podría haber alojado ecosistemas nativos dominados por leñosas antes de que se produjera la introducción masiva de ganado (Marcora et al. 2013). Reconocer como la vegetación actual, la potencial y las alternativas de uso forestal influencian los servicios hídricos de las Sierras de Córdoba es un desafío complejo pero urgente.
AGRADECIMIENTOS: Las primeras etapas de este estudio contaron con el valioso aporte de Gervasio Piñeiro, Rob Jackson, Kathleen Farley y Sheila Palmer. Agradecemos las contribuciones de Celina Santoni, Sergio Contreras, Domingo Alcaraz, Matías Jobbágy y Victoria Marchesini al trabajo de campo y los valiosos comentarios de un revisor anónimo y de la editora. Este trabajo fue financiado por el Inter American Institute for Global Change Research (IAI, CRN 2031).
BIBLIOGRAFÍA
1. ACOSTA, A; S DÍAZ; M MENGHI & M CABIDO. 1992. Patrones comunitarios a diferentes escalas espaciales en pastizales de las Sierras de Córdoba, Argentina. Revista Chilena de Historia Natural, 65:195-207. [ Links ]
2. ARAGÓN, RM; EG JOBBÁGY & EF VIGLIZZO. 2010. Surface and groundwater dynamics in the sedimentary plains of the Western Pampas (Argentina). Ecohydrology, 4:433-447. [ Links ]
3. ARNOLD, JG & PM ALLEN. 1999. Automated methods for estimating baseflow and ground water recharge from streamflow records. Journal of the American Water Resources Association, 35:411-424. [ Links ]
4. ARNOLD, JG; PM ALLEN; R MUTTIAH & G BERNHARDT. 1995. Automated base flow separation and recession analysis techniques. Groundwater, 33:1010-1018. [ Links ]
5. BONALUMI, A; R MARTINO; E BALDO; J ZARCO; J SFRAGULLA; ET AL. 1999. Hoja geológica 3166-IV Villa Dolores, provincias de Córdoba, La Rioja y San Luis, 1:250.000. Servicio Geológico Minero Argentino, Instituto de Geología y Recursos Minerales. Buenos Aires. [ Links ]
6. BRADSHAW, CJA; NS SODHI, KS PEH & BW BROOK. 2007. Global evidence that deforestation amplifies flood risk and severity in the developing world. Global Change Biology, 13:2379-2395. [ Links ]
7. BROWN, AE; L ZHANG; TA MCMAHON; AW WESTERN & RA VERTESSY. 2005. A review of paired catchment studies for determining changes in water yield resulting from alterations in vegetation. Journal of Hydrology, 310:28-61. [ Links ]
8. CABIDO, M; N ATECA; ME ASTEGIANO & AM ANTON. 1997. Distribution of C3 and C4 grasses along an altitudinal gradient in Central Argentina. Journal of Biogeography, 24:197-204. [ Links ]
9. CALDER, IR. 1998. Water use by forests, limits and controls. Tree Physiology, 18:625-631. [ Links ]
10. CONTRERAS, S; EG JOBBÁGY; PE VILLAGRA; MD NOSETTO& J PUIGDEFÁBREGAS. 2011. Remote sensing estimates of supplementary water consumption by arid ecosystems of central Argentina. Journal of Hydrology, 397:10-22. [ Links ]
11. DAY, TJ. 1976. On the precision of salt dilution gauging. Journal of Hydrology, 31:293-306. [ Links ]
12. DORADO, M; E ASTINI; G VERZINO; J DI RIENZO & E PERPIÑAL. 1997. Growth curves for Pinus elliotti, Pinus taeda and Pinus radiata in two areas of the Calamuchita Valley (Córdoba, Argentina). Forest Ecology and Management, 95:173-181. [ Links ]
13. FARLEY, KA; EG JOBBÁGY & RB JACKSON. 2005. Effects of afforestation on water yield: a global synthesis with implications for policy. Global Change Biology, 11:1565-1576. [ Links ]
14. FARLEY, KA; G PIÑEIRO; SM PALMER; EG JOBBÁGY & RB JACKSON. 2009. Stream acidification and base cation losses with grassland afforestation. Water Resources Research, 45:dx.doi.org/10.1029/2007WR006659. [ Links ]
15. GIORGIS, MA; PA TECCO; AM CINGOLANI; D RENISON; P MARCORA; ET AL. 2011. Factors associated with woody alien species distribution in a newly invaded mountain system of central Argentina. Biological Invasions, 13:1423-1434. [ Links ]
16. HAUER, RM & GA LAMBERTI. 1996. Methods in stream Ecology. Academic Press, San Diego, EEUU. [ Links ]
17. HEILMAN, JL; KJ MCINNES; JF KJELGAARD; M KEITH OWENS & S SCHWINNING. 2009. Energy balance and water use in a subtropical karst woodland on the Edwards Plateau, Texas. Journal of Hydrology, 373:426-435. [ Links ]
18. HUXMAN, TE; BP WILCOX; DD BRESHEARS; RL SCOTT; KA SNYDER; ET AL. 2005. Ecohydrological implications of woody plant encroachment. Ecology, 86:308-319. [ Links ]
19. ILSTEDT, U; A MALMER; E VERBEETEN & D MURDIYARSO. 2007. The effect of afforestation on water infiltration in the tropics: A systematic review and meta-analysis. Forest Ecology and Management, 251:45-51. [ Links ]
20. IZURIETA, G; D ABUD & J IZAURRALDE. 1993. Plantaciones de Pinos de la Provincia de Córdoba. Pp. 103 en: Congreso Forestal Argentino y Latinoamericano. A.Fo.A, Paraná, Argentina. [ Links ]
21. JACKSON, RB; SR CARPENTER; CN DAHM; DM MCKNIGHT; RJ NAIMAN; ET AL. 2001. Water in a changing world. Ecological Applications, 11:1027-1045. [ Links ]
22. JACKSON, RB; EG JOBBÁGY & MD NOSETTO. 2009. Ecohydrology in a human-dominated landscape. Ecohydrology, 2:383-389. [ Links ]
23. JOBBÁGY, EG; MD NOSETTO; C SANTONI & G BALDI. 2008. El desafío ecohidrológico de las transiciones entre sistemas leñosos y herbáceos en la llanura Chaco-Pampeana. Ecología Austral, 18:305-322. [ Links ]
24. JOBBÁGY, EG; MD NOSETTO; PE VILLAGRA & RB JACKSON. 2011. Water subsidies from mountains to deserts: Their role in sustaining groundwater-fed oases in a sandy landscape. Ecological Applications, 21:678-694. [ Links ]
25. JONES, JA & GE GRANT. 1996. Peak flow responses to clearcutting and roads in small and large basins, western Cascades, Oregon. Water Resources Research, 32:959-974. [ Links ]
26. KELLIHER, FM; R LEUNING & ED SCHULZE. 1993. Evaporation and canopy characteristics of coniferous forests and grasslands. Oecologia, 95:153-163. [ Links ]
27. LATERRA, P; ME ORUE & GC BOOMAN. 2012. Spatial complexity and ecosystem services in rural landscapes. Agriculture, Ecosystems & Environment, 154:56-67. [ Links ]
28. MARCORA, PI; D RENISON; AI PAÍS-BOSCH; MR CABIDO& PA TECCO. 2013. The effect of altitude and grazing on seedling establishment of woody species in central Argentina. Forest Ecology and Management, 291:300-307. [ Links ]
29. NOSETTO, MD; EG JOBBÁGY; AB BRIZUELA & RB JACKSON. 2012. The hydrologic consequences of land cover change in central Argentina. Agriculture, Ecosystems and Environment, 154:2-11. [ Links ]
30. NOSETTO, MD; EG JOBBÁGY & JM PARUELO. 2005. Land use change and water losses: The case of grassland afforestation across a soil textural gradient in Central Argentina. Global Change Biology, 11:1101-1117. [ Links ]
31. PRINSLOO, FW & DF SCOTT. 1999. Streamflow responses to the clearing of alien invasive trees from riparian zones at three sites in the Western Cape Province. Southern African Forestry Journal, 185:1-7. [ Links ]
32. RICHARDSON, DM. 1998. Forestry trees as invasive aliens. Conservation Biology, 12:18-26. [ Links ]
33. SCANLON, BR; KE KEESE; AL FLINT; LE FLINT; CB GAYE; ET AL. 2006. Global synthesis of groundwater recharge in semiarid and arid regions. Hydrological Processes, 20:3335-3370. [ Links ]
34. SCOTT, DF & W LESCH. 1997. Streamflow responses to afforestation with Eucalyptus grandis and Pinus patula and to felling in the Mokobulaan experimental catchments, South Africa. Journal of Hydrology, 199:360-377. [ Links ]
35. SCHENK, HJ & RB JACKSON. 2002. The global biogeography of roots. Ecological Monographs, 72:311-328. [ Links ]
36. VAN DIJK, AIJM & RJ KEENAN. 2007. Planted forests and water in perspective. Forest Ecology and Management, 251:1-9. [ Links ]
37. VASSALLO, MM; HD DIEGUEZ; MF GARBULSKY; EG JOBBÁGY& JM PARUELO (en prensa). Grassland afforestation impact on primary productivity: a remote sensing approach. Applied Vegetation Science. [ Links ]
38. VIVIROLI, D & R WEINGARTNER. 2004. The hydrological signifance of mountains: From regional to global scale. Hydrology and Earth System Sciences, 8:1016-1029. [ Links ]
39. ZHANG, L; WR DAWES & GR WALKER. 2001. Response of mean annual evapotranspiration to vegetation changes at catchment scale. Water Resources Research, 37:701-708. [ Links ]

