










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.25 no.3 Córdoba dic. 2015
ARTÍCULO ORIGINAL
Respuesta de la biomasa y la composición C:N:P del primer nivel trófico de un arroyo Pampeano a la mayor sequía de los últimos 20 años: Un estudio de caso
Leonardo R. Leggieri1*& Nicolás A. Ferreiro2
1. Laboratorio de Ecotoxicología Acuática, INIBIOMA (CONICET, CEAN), RP 61 Km 3, 8371, Junín de los Andes, Neuquén, Argentina.
2. Grupo de Suelos, INIBIOMA (CONICET, Universidad Nacional del Comahue), Quintral 1250, 8400, S. C. Bariloche, Río Negro, Argentina
*lrleggieri@gmail.com
Editor asociado: Fernando Unrein
Recibido: 30 de noviembre de 2014, Fin de arbitraje: 9 de marzo de 2015, Última versión revisada: 23 de mayo de 2015,Aceptado: 20 de junio de 2015.
Resumen.
El bentos, epifiton, seston, las algas filamentosas libres y las plantas acuáticas proporcionan recursos alimenticios y refugio al resto de los niveles tróficos del arroyo. Aquí estudiamos el efecto de una sequía sobre su biomasa y composición elemental en el arroyo Pampeano La Choza, comparando el año 2007, con ~1200 mm anual, y el año 2008, con ~600mm anual (40% menos que el histórico). Considerando que su abundancia dependería principalmente de la transferencia de masa en la capa límite a bajas velocidades y de la deriva aguas abajo, postulamos (H1) que un año seco genera condiciones desfavorables para las plantas acuáticas y el seston, y favorables para las algas filamentosas epifíticas, bentónicas y de matas libres en un arroyo Pampeano. Como son arroyos ricos en nutrientes, postulamos también (H2) que no hay cambios interanuales apreciables en sus composiciones elementales. Los bajos caudales del 2008 determinaron una mayor conductividad, pH y luz disponible, sin cambios apreciables en las típicas altas concentraciones de nutrientes. La sequía produjo bajos caudales y velocidades, con un detrimento de la cantidad de plantas acuáticas (2±0.3 gm-2) y seston (20±8 gm-2), apoyando la H1. La biomasa de las algas filamentosas perifíticas no se incrementó, al contrario de la H1, y las filamentosas libres crecieron apreciablemente a partir de la primavera, apoyando parcialmente la H1. Los resultados de la composición elemental apoyaron la H2, siendo en el caso del seston y del perifiton muy variable. El impacto de un año de sequía sobre la Pampa Ondulada provocaría la reducción de la biomasa de plantas y seston, favoreciendo el incremento de la biomasa de algas filamentosas libres de los arroyos y, consecuentemente, afectaría a los animales que se alimentan y refugian en ellos.
Palabras clave: Plantas acuáticas; Bentos; Epifiton; Seston; Año seco; Masa; Estequiometria ecológica; Arroyo de la llanura pampeana
Abstract.
Response of the biomass and the C:N:P composition of the first trophic level of a Pampean stream to the highest drought in the last 20 years: A case study. Benthos, epiphyton, seston, free-floating filamentous algae and aquatic plants provide food and shelter to the rest of trophic levels of the stream. Here we studied the effect of an extreme drought on the biomass and elemental composition of La Choza stream in the Pampas by comparing a wet (2007, ~1200 mm/yr) and a dry year (2008, ~600mm or 40% less than the long term mean). Given that the abundance of these communities depends mainly on the mass transfer in the boundary layer at low speeds and the drift downstream, we postulate (H1) that a dry year creates unfavorable conditions for aquatic plants and seston but favors free-floating, epiphytic and benthic filamentous algae. As Pampean streams are rich in nutrients, we also postulate (H2) that there are no significant inter-annual changes in the elemental composition of biomass. Low flow rates of 2008 resulted in a higher conductivity, pH and available light, without appreciable changes in the typical high nutrient concentrations. The drought resulted in lower discharges (<50 L s-1) and velocities (<9 cm s-1), accompanied by declines in the amount of plants (2±0.3 gm-2) and seston (20±8 gm-2) and increases in free-floating but not in filamentous algae, partial supporting H1. Nutrient composition of the biomass did not change as proposed by H2, being highly variable in the case of seston and periphyton. The impact of a full-year drought on the Wet Pampa could affect animal communities as plants and seston biomass decreases in favor of increase of free-floating algae, compromising food and shelter.
Keywords: Aquatic plants; Benthos; Epiphyton; Seston; Dry-year; Mass; Ecological stoichiometry; Lowland pampean stream
INTRODUCCIÓN
En los arroyos, la magnitud y la frecuencia de las crecidas afectan a las comunidades, lo que produce cambios en el flujo de energía a través de las redes tróficas y el reciclado de nutrientes (Galat et al. 1998). El bentos, epifiton, seston, las algas filamentosas libres y plantas acuáticas constituyen compartimentos fundamentales de los arroyos, ya que proporcionan recursos alimenticios y de refugio, sosteniendo a gran parte de la red trófica del arroyo. Existen valores de corrientes máximas y mínimas que determinan un rango, para cada compartimento basal, donde su abundancia tiende a ser máxima. Este rango depende también del tipo de sustrato (grava, sedimento, limo, etc.) y de factores limitantes como la luz y los nutrientes. Por un lado, la velocidad y el caudal alto, provocado por crecidas importantes y frecuentes, determinan el transporte de parte de esta biota hacia aguas abajo, tanto por la fuerza de la corriente como por la abrasión de los sedimentos suspendidos (Matthaei et al. 2003; Riis & Biggs 2003). En arroyos con baja pendiente y diferentes sustratos, casi todas las plantas acuáticas suelen ser arrastradas a velocidades (v) >80 cm s-1, a causa de la deriva del sustrato que solía sostener sus raíces (Riis & Biggs 2003; Franklin et al. 2008). Asimismo, en arroyos y ríos llanos, a v >100 cm s-1, decrece al mínimo la biomasa del bentos (Matthaei et al. 2003), del epifiton (Biggs 1995) y de las algas filamentosas (Stevenson 1996).
Por otro lado, las lluvias escasas pueden determinar velocidades del agua lo suficientemente bajas (< 20 cm s-1) como para afectar la transferencia de masa a través de la capa límite, desde el entorno hacia los organismos. La viscosidad y espesor de la capa límite aumentan a bajas velocidades, limitando la disponibilidad de carbono (Riis & Biggs 2003), la captación de oxígeno (ej. Jørgensen & Des Marais 1990) y la captación de nitrógeno y fósforo (Larned et al. 2004). Por ejemplo, la tasa fotosintética y respiratoria de las plantas tiende a decrecer hacia menores velocidades, dentro del rango de 0 a 10-30 cm s-1 y, consecuentemente, tiende a decrecer su biomasa (Riis & Biggs 2003; Franklin et al. 2008). En el caso de los organismos perifíticos (epifíticos y bentónicos), también suele decrecer la tasa metabólica específica pero esto no necesariamente implicaría una reducción de biomasa; ya que la biomasa de perifiton tiende a aumentar progresivamente hacia menores v del agua, dentro del rango 0-20 cm s-1, porque las algas filamentosas suelen dominar a bajas velocidades mientras que las diatomeas lo hacen a velocidades intermedias (Biggs et al. 1998; Larned et al. 2004). No obstante, estas tendencias dependen también de otros factores como los nutrientes disponibles en el arroyo, el tipo de sustrato al que está adherido el perifiton y la composición de la comunidad perifítica.
La viscosidad de la capa límite y la tasa de deriva aguas abajo también influyen sobre la masa de seston en la columna de agua (Li et al. 2013). Su masa depende también de su composición específica y de la proporción de materia particulada no-viva. Por ejemplo, las cianobacterias son favorecidas por temperaturas >20°C y altas concentraciones de fósforo (Downing et al. 2001), mientras que las algas verdes y diatomeas son favorecidas por flujos macroturbulentos, v > 30 cm s-1 (Sherman et al. 1998). La cantidad de materia orgánica no-viva sestónica suele ser proporcional a las escorrentías recientes y a la velocidad de la corriente, que pone en suspensión a parte del bentos. Dado que en los arroyos Pampeanos no se han detectado autótrofos netamente sestónico, la componente biótica del seston sería proporcional al desprendimiento de perifiton (Giorgi & Feijoó 1999); en cuyo caso la masa de seston incrementaría con la intensidad de la corriente.
Por lo tanto, la accesibilidad a los recursos nutricionales para cada compartimento, obtenidos en la capa límite, aumentan con la velocidad hasta el punto donde la materia es arrastrada por la corriente. Así, la abundancia del primer nivel trófico depende del trade-off entre la transferencia de masa y la deriva aguas abajo. Como consecuencia, la biomasa de la mayoría de las plantas resulta máxima a v∼ 30-50 cm s-1 (Riis & Biggs 2003; Franklin et al. 2008), al igual que la masa de seston; mientras que la masa del perifiton suele ser máxima a v∼ 10-20 cm s-1, dependiendo de la abundancia relativa de algas filamentosas (Biggs et al. 1998; Larned et al. 2004).
La velocidad basal promedio es de 2-40 cm s-1 en los arroyos de la Pampa Ondulada, debido a la baja pendiente de las llanuras Pampeanas (Leggieri et al. 2013). Podría esperarse entonces, a partir de los trabajos previos comentados arriba, que la biomasa de las plantas acuáticas y la masa de seston sean máximas en el extremo superior de este rango (30-40 cm s-1). Mientras que la biomasa del perifiton epifítico y bentónico habría de ser máxima en el rango inferior (2-20 cm s-1). Aquí estudiamos a los representantes del primer nivel trófico del arroyo La Choza (NE de la Pcia. Buenos Aires) durante el 2007 y el 2008. Usualmente, las precipitaciones son de ∼1100 mm anuales, pero el año 2008 correspondió al año más seco de los últimos 20 años, con tan solo 606 mm anual (Anexo I). El menor caudal durante el 2008 respecto del 2007, podría haber determinado la reducción de la masa de plantas y seston, y el aumento de la biomasa de algas filamentosas epifíticas y bentónicas. Nuestra primera hipótesis es que un año seco genera condiciones desfavorables para las plantas acuáticas y el seston, y favorables para el epifiton, el bentos y las algas filamentosas libres de un arroyo Pampeano.
Como los arroyos Pampeanos son ricos en nutrientes (Leggieri et al. 2013), estos no serían la limitante para el crecimiento de los componentes del primer nivel trófico y, por lo tanto, esperaríamos que no hubiese cambios apreciables en su composición C: N:P. La concentración de fósforo y nitrógeno disponible para las plantas de los arroyos Pampeanos suelen ser superior a los umbrales empíricos para su fotosíntesis, incluso en las estaciones con menos precipitaciones (Leggieri et al. 2013). Por lo tanto, es esperable que la composición C:N:P de las plantas sea similar en los años 2007 (con lluvias típicas) y 2008 (con sequía). Para los organismos que componen al perifiton y al seston, los nutrientes y la luz no serían limitantes, pero sus composiciones C:N:P podrían variar por cambios en la composición específica y en la proporción de materia orgánica no-viva en cada compartimento (Leggieri 2014). Dado que no se cuenta con información para establecer a priori si habría cambios o no en su composición C:N:P desde el año 2007 al 2008, proponemos, con visión conservadora, que la composición C:N:P sería similar entre años. Por lo tanto, en la segunda hipótesis proponemos que un año seco no determina cambios apreciables (a escala anual) en la composición C:N:P de los representantes del primer nivel trófico de un arroyo Pampeano.
Materiales y Métodos
Área de estudio y muestreos
La Región Pampeana es una planicie sedimentaria fértil cubierta originalmente por pastizales que se encuentra situada en el centro-este de la República Argentina y se caracteriza por una temperatura anual media de 17°C y precipitaciones anuales promedio de 1100 mm (Soriano et al. 1992). El estudio se llevó a cabo en un tramo de 100 m del arroyo La Choza (34°44'34''S - 59°06'37''O), un tributario de la cuenca Río platense que se origina en leves depresiones del suelo (∼15 km aguas arriba del tramo). Su lecho está compuesto por sedimentos finos (principalmente limo y arcilla), sin ningún tipo de rocas. Su valle aluvial es relativamente ancho (50–100 m), como la mayoría de los arroyos de la llanura pampeana. La vegetación ribereña del tramo estudiado y del sector aguas arriba estaban relativamente bien conservadas en los primeros 30 m desde el borde del agua, con dominancia de gramíneas y ausencia de árboles introducidos.
Las campañas de muestreo se llevaron a cabo durante febrero, mayo, julio, septiembre y diciembre de los años 2007 and 2008 (verano, otoño, invierno, primavera temprana y primavera tardía). Los muestreos se hicieron al menos 5 días después de la última lluvia con el fin de reducir la interferencia de las lluvias-escorrentías sobre las características hidrológicas, físico-químicas y biológicas del arroyo. Los datos de las precipitaciones diarias de los últimos 20 años fueron obtenidos de la estación meteorológica de la Universidad Nacional de Luján, ubicada a 21 Km del tramo estudiado. A partir de los datos diarios se calcularon las precipitaciones acumuladas mensuales y anuales de la Figura 1.
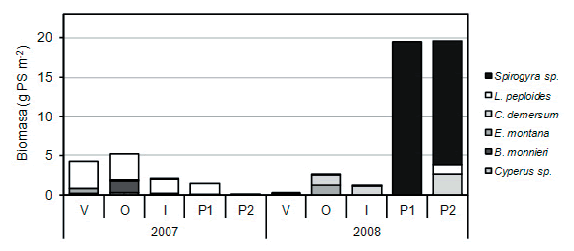
Figura. 1. Biomasa (gramos de peso seco por m2 de arroyo) de las principales especies de plantas acuáticas y algas filamentosas en el arroyo La Choza durante el verano (V), el otoño (O), el invierno (I), la primavera temprana (P1) y la primavera tardía (P2) de los años 2007 y 2008. La biomasa de la especie dominante L. peploides disminuyó hacia el 2008 y la biomasa del alga Spirogyra sp. aumentó considerablemente en la primavera del 2008.
Figure. 1. Biomass (grams of dry weight per m2 of stream) of the main species of aquatic plants and filamentous algae in the La Choza stream during Summer (V), Autumn (O), Winter (I), early Spring (P1) and late Spring (P2) of the 2007 and 2008 years. The dominant specie of aquatic plant was L. peploides. Their biomass decreased in 2008 and biomass of the Spirogyra sp. algae increased considerably in the spring of 2008.
Factores físico-químicos
La estructura física del cauce se estudió de acuerdo a Elosegui & Sabater (2009) a intervalos regulares de 10 m. El caudal y la velocidad media se estimaron mediante el método de adición de sal (Gordon et al. 2004). La temperatura, el pH, la conductividad y el oxígeno se midieron mediante instrumental de campo al mediodía (peachímetro Hanna HI 9023; conductímetro Hanna HI 9033; oxímetro Hach HQ30D). El porcentaje de radiaciones fotosintéticamente activas (PAR) que alcanzaban el fondo del lecho se calcularon como las PARfondo / PARsuperficie del agua por 100. Las mediciones de las PAR se hicieron utilizando un sensor PAR (PAR-Lite, Kipp & Zonen, Delft, Países Bajos) y registrador de datos (CR1000, Campbell Scientific Inc., Utah, EE.UU.), tomando mediciones en el fondo y en la superficie al mediodía de días sin nubes (cuando las PAR son máximas). Se usó el % PAR (y no el valor absoluto de PAR) para reducir la variabilidad entre las estaciones del año. Las muestras de agua para el análisis de nutrientes se colectaron por triplicado en botellas de polietileno, que se almacenaron a 4°C, se transportaron al laboratorio y se analizaron dentro de las 4 h. Las muestras fueron filtradas con filtros de fibra de vidrio (Whatman GF/F, Maidstone, UK; los filtros fueron guardados para la estimación de la masa del seston, ver sección siguiente) y se determinó la concentración de fósforo (P) como fósforo reactivo soluble (PRS) con el método del acido ascórbico, la concentración de nitratos y nitritos mediante la reacción con sulfanilamida y la concentración de amonio mediante el método del fenolhipoclorito (APHA 1998). El nitrógeno inorgánico disuelto (NID) se calculó como la suma de nitratos, nitritos y amonio. El nitrógeno total disuelto (NTD) y el fósforo total disuelto (PTD) se determinó por los mismos métodos descriptos pero con una previa oxidación con peroxi-disulfato (APHA 1998). Las fracciones orgánicas, nitrógeno orgánico disuelto (NOD) y fósforo orgánico disuelto (POD), se calcularon como la diferencia entre los nutrientes totales e inorgánicos. El N y el P particulado (NP, PP) en suspensión se determinó a partir de la materia retenida en los filtros, mediante combustión seca para el NP (autoanalizador Leco TruSpec CN, Columbia Analytical Services, Washington, USA) y, para el PP, mediante combustión a 480°C, digestión con peroxidisulfato y análisis con el método del ácido ascórbico (APHA 1998). Luego, estos contenidos de N y P se extrapolaron al volumen de agua filtrado. El nitrógeno total (NT) y el fósforo total (PT) se calcularon como la suma de las fracciones: NT = NTD + NP y PT = PTD + PP, respectivamente.
Masa
El mapeo de la distribución de plantas acuáticas, algas filamentosas y el bentos se realizó de acuerdo a Elosegui & Sabater (2009) con 20 subtramos de 5 m de largo. Tres muestras de cada compartimento fueron colectadas en cada muestreo. Las muestras de plantas y algas filamentosas fueron colectadas de manchones con un marco cuadrado (400 cm2), almacenadas en bolsas de polietileno y transportadas al laboratorio. Se consideraron las especies de plantas y algas filamentosas que en los dos años tuvieron un promedio mayor a 0.05 g PS m-2. En el laboratorio, las muestras de plantas y algas filamentosas fueron lavadas, separando los sedimentos y las algas adheridas, y se secaron a 60°C hasta peso seco constante (PS). La masa de epifiton fue extraída de fragmentos de L. peploides y estimada de acuerdo a Vilches (2012). Los microorganismos y materia no-viva del fondo se muestrearon conjuntamente, considerándolos como el bentos, de acuerdo a Gómez et al. (2009). Los microorganismos y materia no-viva (> 0.7 µm) de la columna se muestrearon conjuntamente, considerándolos como el seston (Gómez et al. 2009). Las muestras de bentos fueron secadas a 60°C hasta peso seco constante (PS) y calcinadas a 500°C por 2 h (obtención del PS libre de cenizas, PSLC). La masa del bentos se estimó como la diferencia entre el PS y el PSLC. Los filtros GF/F con seston, obtenidos del agua filtrada (ver sección Factores físico-químicos), fueron secados a 60°C hasta PS. La masa de epifiton se expresó también en porcentaje de g PS epifiton / g PS planta. Combinando los datos de PS de cada compartimento con los datos del mapeo y los filtrados, se calculó la masa promedio de cada compartimento por superficie de agua del arroyo (g PS m-2).
Composición elemental
Los contenidos de carbono (C) y nitrógeno (N) de los compartimentos se determinaron mediante el análisis elemental por combustión seca (autoanalizador Leco TruSpec CN, Columbia Analytical Services, Washington, USA) (APHA 1998). Los contenidos de fósforo (P) se determinaron por combustión a 480°C, seguido por una digestión (con peroxidisulfato para el seston, el epifiton y el bentos; y con HCl 25% para las plantas acuáticas) y la determinación de P mediante el método del ácido ascórbico (APHA 1998). En el caso de las plantas, se seleccionaron y usaron tejidos jóvenes de tallos y hojas con el fin de minimizar el error asociado a la variabilidad de las concentraciones de C, N y P que hay entre los distintos tejidos de la planta. Se consideró a la especie de planta dominante, Ludwigia peploides. No se hizo el análisis elemental de las algas filamentosas porque durante la mayor parte del estudio su presencia fue mínima. A partir de los contenidos de C, N y P, se calcularon las relaciones molares C:N, C:P y N:P de cada compartimento en cada fecha de muestreo.
Análisis de los datos
La normalidad de las variables usadas se evaluó con la prueba de Shapiro-Wilkinson y se transformaron cuando ello fue necesario. Exploramos las diferencias entre los años 2007 (año típico) y el año 2008 (año relativamente seco) en la magnitud de cada variable mediante ANOVAs (cuando era normal o se pudo normalizar) y con la prueba no paramétrica de Wilcoxon (cuando no se pudo normalizar), usando como n a los promedios de cada variable en cada estación del año (n = 5 en cada grupo). Usamos al coeficiente de variación (% CV = 100 desvío estándar / promedio) como estimador de la variabilidad intra-anual de cada variable. Si fuese posible encontrar una diferencia interanual, incluyendo la variabilidad estacional, entonces los cambios producidos por las pocas lluvias del 2008 serán evidentes en los resultados del ANOVA y de la prueba de Wilcoxon. Todos los análisis se realizaron con el programa SPSS 11.5 (Inc., Chicago, Illinois, USA).
Resultados
Factores físico-químicos
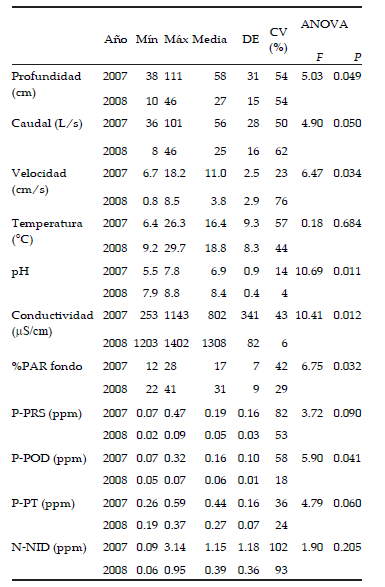
Las características basales del agua presentaron algunas diferencias entre años: la profundidad, la velocidad y el caudal fueron significativamente menores en el año seco (2008). El pH fue neutro en el año típico pero fue básico (>8) durante el año seco. La conductividad fue significativamente mayor en el año seco (2007: ∼ 800 S cm-1; 2008: ∼1300 µS cm-1). Asociado a las menores profundidad y materia en suspensión, el porcentaje de PAR en el fondo fue casi el doble en el 2008 (31 %) que en el 2007 (17 %). Respecto de los nutrientes N y P en la columna, sus concentraciones no variaron significativamente entre años, a excepción del fósforo orgánico disuelto, que fue menor en el año seco (2007: ~ 0.16 ppm; 2008: ~ 0.06 ppm). La variabilidad intra-anual de la concentración de cada nutriente en la columna fue relativamente alta (CV∼ 20-100 %; Tabla 1).
Tabla 1. Factores físico-químicos en el arroyo La Choza durante el año 2007 y 2008. Se muestran los valores mínimos y máximos (mín-máx), los valores medios anuales, el desvío estándar (DE) y los coeficientes de variación CV intra-anuales (n=5). En las dos columnas de la derecha se muestran los resultados del ANOVA: valores F y significancia (P).
Table 1. Physico-chemical factors in the La Choza stream during 2007 and 2008. Columns show minimum and maximum values (min-max), annual means, standard deviation (DE) and intra-annual coefficients of variation CV (n=5). Columns on the right show ANOVA F and p values.
Masa
En el caso de las plantas, la especie más abundante fue L. peploides, con un promedio en los dos años de 1.2 (± 1.3) g m-2, seguido por Ceratophyllum demersum (0.5 ± 0.8 g m-2), Eleocharis montana (0.2 ± 0.4 g m-2), Bacopa monnieri (0.2 ± 0.5 g m-2) y Cyperus sp. (0.1 ± 0.1 g m-2; Figura 1).
En el caso de las algas filamentosas, la única representativa en este arroyo durante los muestreos fue Spirogyra sp., con un promedio en los dos años de 3.5 (± 7.5) g m-2, que corresponde a una muy baja biomasa (< 0.05 g m-2) hasta la primavera de 2008, seguido de un importante incremento de biomasa (> 15 g m-2). La biomasa de L. peploides se redujo a más de un sexto durante el año seco (~0.3 g m-2) respecto del año típico (~2.0 g m-2, p < 0.05; Tabla 2), mientras que no hubo diferencias significativas entre años en la biomasa de las otras especies de plantas y de Spirogyra sp. (ZSpirogyra sp. = -0.17, p < 0.865; ZCyperus sp. = -1.69, p < 0.091; ZE.montana = -0.89, p < 0.373; ZB.monnieri = -1.35, p < 0.176; ZC.demersum = -0.42, p < 0.674).
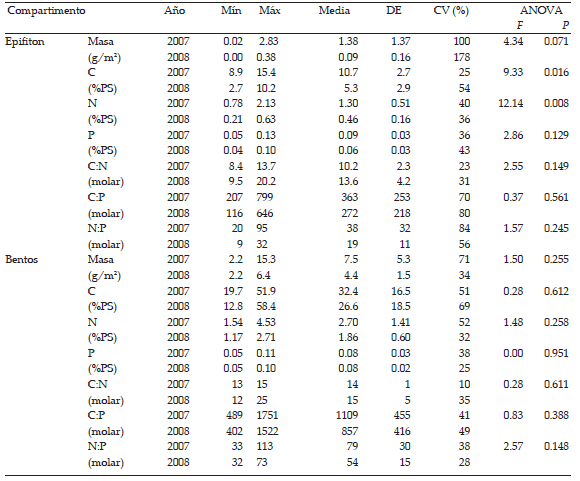
Tabla 2. Cantidad; contenidos de C, N y P (% del peso seco); y sus relaciones molares en la planta L. peploides, el seston, el epifiton y el bentos colectados en el arroyo La Choza durante el año 2007 y 2008. Se muestran los valores mínimos y máximos (mín-máx), los valores medios anuales, el desvío estándar (DE) y los coeficientes de variación CV intra-anuales (n=5). En las dos columnas de la derecha, se muestran los resultados del ANOVA: valores F y significancia (P).
Table 2. Mass; C, N and P contents (%DW); and their molar ratios for L. peploides (plant), seston, epiphyton and benthos sampled in the La Choza stream during 2007 and 2008. Columns show minimum and maximum values (min-max), annual means, standard deviation (DE) and intra-annual coefficients of variation CV (n=5). Columns on the right show ANOVA F and P values.
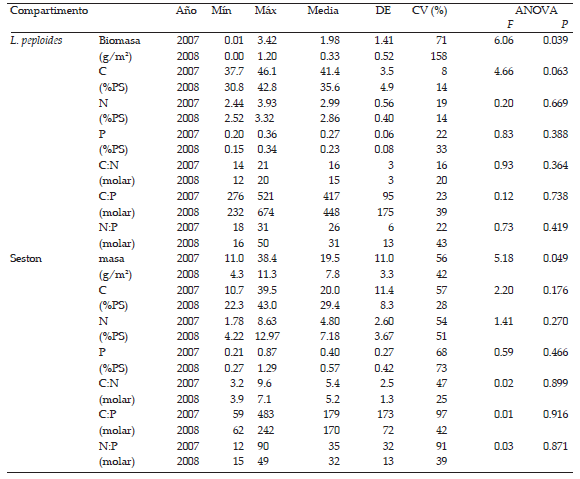
La masa de seston también se redujo a menos de la mitad durante el año seco (2007: ~20 g m-2; 2008: ~8 g m-2). Mientras que no hubo diferencias significativas entre los años 2007 y 2008 en la masa de epifiton y bentos (Tabla 2). Tampoco hubo diferencias significativas
en el porcentaje de epifiton por unidad de biomasa de planta entre años (2007 = 49 ± 39 % p/p y 2008 = 33 ± 18 % p/p; promedio ± desvío estándar) (F = 0.65, p = 0.44).
Composición elemental
Las condiciones del 2008 no afectaron significativamente la composición elemental y las relaciones C:N:P de L. peploides. Éstas fueron muy poco variables tanto intra-anualmente (siempre CV < 50%) como inter-anualmente (p > 0.05). La relación C:N: P promedio de L. peploides de los dos años fue 432:28:1, con una tendencia hacia menores relaciones C:Nutrientes en el 2007 (417:26:1) que en el 2008 (448:31:1). Las condiciones del 2008 tampoco afectaron significativamente la composición elemental del seston y del bentos, mientras que las concentraciones de C y N del epifiton fueron menos de la mitad durante el año menos lluvioso 2008 que durante el 2007. Las relaciones C:N:P del seston, del epifiton y del bentos no tuvieron diferencias apreciables entre años. En promedio, de los dos años, la relación C:N:P del seston fue 174:34:1, del epifiton fue 318:29:1 y del bentos fue 983:67: 1.
Discusión
El descenso en el caudal del arroyo La Choza durante el 2008 tuvo un efecto negativo sobre la biomasa de plantas acuáticas y la masa de seston, apoyando la hipótesis según la cual la sequía genera condiciones desfavorables para estos. Por otro lado, en discordancia con la hipótesis, el descenso del caudal no favoreció a las algas filamentosas perifíticas y sólo parecería haber favorecido a las algas filamentosas libres hacia el comienzo de la primavera del año seco. La sequía parecería favorecer la transición paulatina hacia una menor heterogeneidad del hábitat (menos plantas acuáticas) de invertebrados y peces pequeños, hacia una mayor salinidad (mayor pH y conductividad) y hacia un sistema acuático dominado por algas filamentosas libres, más vulnerable a las perturbaciones hidrológicas. Finalmente, no se detectó cambio apreciable en la composición C:N:P de ningún compartimento basal asociado a la sequía, apoyando la hipótesis de la no limitación por nutrientes para este nivel trófico.
Según las concentraciones de nitrógeno total (1.6-7.0 ppm) y fósforo total (0.2-0.6 ppm), las comunidades del arroyo no habrían tenido limitación por nutrientes (NT>1.5 ppm, PT>0.075 ppm; según el criterio de Dodds et al. 1998). El mantenimiento de las concentraciones altas de nutrientes en las aguas de este arroyo pampeano ante el bajo ingreso de nutrientes por las escorrentías del 2008 puede explicarse por el sustrato mismo del cauce, un sustrato desarrollado y fértil (Toledo 2011).
Efecto de la sequía sobre la abundancia del primer nivel trófico
Los resultados sustentan la primera hipótesis para el caso de las plantas acuáticas (que propone que en un año seco su masa decrece): la biomasa de L. peploides fue seis veces menor durante el 2008 (0.3 g/m2) respecto del 2007 (2 g/m2). Respecto del año anterior, las condiciones hidrológicas del año seco podrían haber aumentado la viscosidad de la capa límite y generado una restricción en la transferencia de masa para el desarrollo de L. peploides, similar a lo observado en otros estudios (Larned et al. 2004). Dado que la concentración de nutrientes siguió siendo alta durante el 2008, la restricción en la transferencia del CO2 y O2 a través de la capa límite podría haber sido más importante que la transferencia de nutrientes.
En el 2008, el incremento de pH a 8-9 implica que gran parte del HCO3- biodisponible pasa a CO3-2, probablemente como CaCO3, sumándose a la posible limitación de C para las plantas. Por otro lado, el crecimiento del epifiton sobre las plantas, que podría haber reducido la disponibilidad de luz, no aumentó significativamente y no habría sido un factor determinante. El menor caudal podría haber favorecido la permanencia de los consumidores detritívoros y herbívoros de plantas, reduciendo también su biomasa (Brinson et al. 1981). En el arroyo La Choza, los análisis de isótopos estables sugieren que L. peploides fue consumida sólo por el caracol Pomacea canaliculata en el 2007 y por P. caniculata y el pez Cyphocharax voga (sabalito) en el año seco (Leggieri 2014; Feijoó et al. 2014). En estudios de largo plazo se ha demostrado que una reducción a la mitad del caudal, similar a lo ocurrido del 2007 al 2008 en el presente estudio, favorece la permanencia de caracoles, de otros macroinvertebrados (Holt et al. 2014) y de peces (Bice et al. 2014). En el arroyo La Choza, Colautti y colaboradores (2009) encontraron que la abundancia relativa de C. voga aumentó de 2% en noviembre del 2007 a más de 20% en junio del 2008. Asociando esto a que. L. peploides no fue consumida por C. voga en el 2007 y que sí formó parte de su dieta en el 2008 (Leggieri 2014), sería factible que C. voga contribuyera también al desmedro de L. peploides durante el año seco. Dado que las plantas, junto con el epifiton que albergan, representan refugio y sirven de alimento para los peces e invertebrados (Ferreiro et al. 2010), la reducción de la biomasa de plantas debido a períodos prolongados de pocas lluvias, como el año 2008, pueden afectar no sólo a las plantas acuáticas sino a todo el ecosistema del arroyo.
Las algas filamentosas estuvieron casi ausentes desde el verano del 2007 hasta la primavera de 2008, cuando comenzó una floración importante (>15 g/m2), que duraría toda la primavera. Es probable que las características morfológicas de Spirogyra sp., como así también su poca o nula adherencia a un sustrato y su flotabilidad elevada, hayan determinado su poco crecimiento localizado y su deriva fácil aguas abajo (Wetzel 2001) durante el año con lluvias típicas, 2007. Mientras tanto, sus tasas elevadas de producción y respiración bajo estabilidad hidrológica habrían favorecido su incremento de biomasa después del invierno del 2008 (Wetzel 2001; Leggieri et al. 2013). Si bien el aumento de biomasa de algas filamentosas tardó más de lo esperado a partir de la primera hipótesis, nuestros resultados apoyarían la idea de que hacia menores velocidades las comunidades de plantas son paulatinamente reemplazadas por las algas filamentosas.
Los resultados del seston sustentan la primera hipótesis, que postula que en un año seco su masa decrece debido al menor arrastre de materia perifítica. Esto pudo deberse al menor ingreso de material abiótico procedentes de la cuenca de drenaje, como así también a la disminución de la componente biótica ante los menores caudales, ya que las especies que suelen encontrarse en la columna de agua son organismos perifíticos, presumiblemente desprendidos por la abrasión de la corriente o como estrategia de dispersión (Giorgi & Feijoó 1999). Esto último se sustenta también por el incremento de clorofila "a" sestónica durante las estaciones del año más lluviosas (Leggieri 2014). Por lo tanto, menores escorrentías y caudales implican una menor masa sestónica, tanto sea por la reducción de componentes abióticos como bióticos. Por último, la masa del perifiton no varió apreciablemente ante los bajos caudales de 2008, aunque existió cierta tendencia a ser menor en el año seco (epifiton, 1.4±0.1 g/m2; bentos, 7.5±4.4 g/m2).
Efecto de la sequía sobre la composición elemental del primer nivel trófico
Los resultados del contenido elemental en las plantas apoyaron la segunda hipótesis, que sostiene que los contenidos elementales de las plantas acuáticas flotantes arraigadas no varían considerablemente en un arroyo eutrófico durante un año con pocas lluvias. Durante este estudio en el arroyo La Choza, las plantas no habrían tenido limitación por nutrientes dado que las concentraciones de nitrógeno y fósforo (0.77±0.91 ppm N-NID, 0.12±0.13 ppm P-PRS) fueron superiores a los umbrales empíricos para el rendimiento de la fotosíntesis de las plantas acuáticas (0.10 ppm N-NID, 0.01 ppm P-PRS) (Demars & Edwards 2007). La no-limitación por nutrientes se evidencia también en que sus contenidos de N y P (2.93±0.46 %N y 0.25±0.07 %P) fueron superiores al 60% de los valores umbrales para el crecimiento (1.82 %N y 0.16 %P; Gerloff 1975) (Colman et al. 1987), y en la baja relación C:P (432±133, promedio±DE) respecto de los resultados de otros estudios: C:P∼517-570 (Atkinson & Smith 1983; Duarte 1992; Demars & Edwards 2007).
Sustentando la segunda hipótesis, la composición elemental del seston, del epifiton y del bentos no cambió apreciablemente entre años, y presentó variaciones intra-anuales (inter-estacionales) del C:N:P (CV~10-100). De modo que la interpretación de la ausencia del efecto de la sequía sobre las composiciones elementales sólo es válida a escala interanual. La variabilidad elevada de las relaciones elementales podría asociarse al pastoreo de distinto tipo de herbívoros (Fitter & Hillebrand 2009), al diferente estado sucesional de las comunidades, a la deriva aguas abajo de los organismos menos adheridos al sustrato (Matthiessen et al. 2010) y a las variaciones en interacción huésped-epífitas (Croll 1983).
En el caso del bentos, es factible que las muestras contuvieran una proporción alta de detritos (material abiótico acumulado en el fondo) pobres en P, ya que la relación C: P fue demasiado elevada (∼400-1700) para una comunidad bentónica que habita en un ecosistema rico en nutrientes. La relación elemental del bentos, similar en el 2007 y el 2008, concuerda con los resultados de Cochero et al. (2013), quienes estudiaron a las algas y bacterias bentónicas durante el 2007 y el 2008 en el arroyo La Choza y no hallaron diferencias significativas entre años. La poca variación del bentos ante los bajos caudales podría tener importancia en el sostenimiento del ecosistema, ya que la parte de la red trófica que hace uso del bentos como recurso mantendría su fuente de alimento ante la reducción de caudal. En este sentido, las consumidores menos afectados por la sequía serían P. argentinus, Aegla sp. e Hypostomus commersoni, que según los análisis estomacales y de isótopos estables consumieron al bentos tanto en el 2007 como en el 2008 (Leggieri 2014; Feijoó et al. 2014).
Conclusiones
Durante el año 2008, el más seco de los últimos 20 años, el caudal y la velocidad del arroyo La Choza fueron la mitad que las medias históricas. En consecuencia, la salinidad y la alcalinidad del agua fueron altas, mientras que los nutrientes disponibles se mantuvieron regularmente altos. Suponemos que esto último contribuyó a la invariabilidad de la composición C:N:P del seston, del perifiton y de las plantas acuáticas. El menor caudal se vinculó a la disminución de la masa de plantas acuáticas y seston, según lo esperábamos, lo que restringió la cantidad de hábitats disponibles para invertebrados y peces. En cambio, la biomasa de algas filamentosas epifíticas y bentónicas no aumentó, al contrario de lo que esperábamos por la reducción de caudal, mientras que las algas filamentosas flotantes comenzaron a ser abundantes a partir de la primavera del año seco. Estos resultados sustentarían la idea de que, con un menor caudal, las comunidades de plantas son paulatinamente reemplazadas por las algas filamentosas. No obstante, sólo contamos con este caso de estudio, que no puede ser extrapolado. Será necesario ahondar con más estudios teóricos y empíricos para comprender cómo la sequía puede afectar a los arroyos pampeanos.
Contexto global: fenómeno de La Niña
Las precipitaciones varían intra-anualmente en el noroeste de la provincia de Buenos Aires, y son mayores en primavera y verano que en otoño e invierno (Figura 2). Esto depende del ingreso de masas de aire seco frío provenientes del sur (polares), que tienen un mayor avance sobre el territorio durante el invierno y el otoño. A su vez, las precipitaciones varían inter-anualmente por otros fenómenos físicos, como el El Niño-Oscilación del Sur (ENOS), que implica un aumento de más de 0.5 °C en un sector del Océano Pacífico, y como el fenómeno de La Niña, que implica un descenso de más de 0.5 °C en el mismo sector. En lo que a nuestro trabajo atañe, el ENOS ocurrió durante el año 2007 (+1.0 °C) y La Niña ocurrió durante el año 2008 (-1.4 °C), ambos clasificados como moderados (entre 1 y 1.5 °C; NOAA 2009). Sobre el sector del arroyo estudiado, la ocurrencia del ENOS durante el 2007, según los registros históricos del Servicio Meteorológico Nacional, implicaría un incremento de las precipitaciones de 130 a 275 mm (SMN 2014). En concordancia con éstas predicciones, el año 2007 presentó una desviación de 194 mm más respecto del promedio histórico (1276 mm vs. 1082 mm; Figura 1). Por otro lado, sobre el sector del arroyo estudiado, la ocurrencia de La Niña durante 2008, implicaría un decrecimiento de las precipitaciones de 25 a 50 mm/año (con un decrecimiento de 50-125 mm en invierno-primavera, y un incremento de 30-75 mm en verano-otoño; SMN 2014). El decrecimiento de las precipitaciones durante el 2008 fue mucho mayor que esta predicción, con una desviación de 476 mm menos respecto del promedio histórico (606 mm menos 1082 mm; Figura 1), relacionado con un verano-otoño con 270 mm de precipitaciones menores a la histórica. Este trabajo es un ejemplo de cómo los procesos físicos ocurridos en el océano Pacífico pueden terminar afectando al desarrollo de compartimentos de un arroyo pampeano tan importantes como las plantas acuáticas (que ofrecen refugio para peces e invertebrados y que son el sustrato del epifiton), y como el seston (que sirve de alimento a filtradores).
NOAA (National Oceanic and Atmospheric Administration). 2009. www.pmel.noaa.gov/tao/elnino/el-nino-story.html
SMN (Servicio Meteorológico Nacional). 2014. Informes de El Niño de cada periodo, informe de enero de 2009, análisis de ENOS del 2007 y del 2008: www.smn.gov.ar/serviciosclimaticos/?mod=vigilancia&id=3.

Figura 2. Arriba: precipitación anual desde 1988 hasta 2012 en la Ciudad de Luján, Buenos Aires. La línea gris muestra el promedio anual desde 1988 hasta 2006 (antes del primer muestreo), 1082 mm. Los rombos grises indican los dos años en que se efectuaron los trabajos de campo, 2007, con 1276 mm/año, y 2008, con 606 mm/año. Obsérvese que el año 2008 fue el menos lluvioso de todo el registro (1988-2012). Abajo: precipitación en los meses de los años 2007 y 2008. La línea gris muestra el promedio histórico de cada mes (1988-2006). Obsérvese que a partir de febrero del 2008, la precipitación mensual siempre fue menor que los promedios históricos.
Figura 2. Above: annual precipitation from 1988 to 2012 in the Luján City, Buenos Aires. Argentina. The gray line shows the annual average from 1988-2006 (before the first sampling date), 1082 mm. The gray diamonds indicate the two years in which the fieldwork were done, year 2007 with 1276 mm and year 2008 with 606 mm. Note that 2008 rainfalls were the lowest of the entire record (1988-2012). Below: rainfalls in the months of the years 2007 and 2008. The gray line shows the historical average of each month (1988-2006). Note that after February 2008, the monthly rainfalls were always lower than historical averages.
Agradecimientos. Agradecemos a J. De Anna, M. Da Silva, E. Zunino, C. Rodríguez Castro, R. Leggieri, A. Giorgi y C. Vilches por su asistencia en los trabajos de campo. Agradecemos el apoyo económico brindado por el FONCyT, por la Universidad Nacional de Luján (Buenos Aires) y por la Fundación BBVA.
Bibliografía
1. APHA (American Public Health Association). 1998. Standard Methods for the Examination of Water and Wastewater. American Public Health Association Inc., Washington DC, USA. [ Links ]
2. Atkinson, MJ & SV Smith. 1983. C:N:P ratios of benthic marine plants. Limnol. Oceanogr., 28:568-574. [ Links ]
3. Bice, CM; SL Gehrig; BP Zampatti; JM Nicol; P Wilson; et al. 2014. Flow-induced alterations to fish assemblages, habitat and fish-habitat associations in a regulated lowland river. Hydrobiologia, 722:205-222. [ Links ]
4. Biggs, BJF. 1995. The contribution of flood disturbance, catchment geology and land use to the habitat template of periphyton in stream ecosystems. Freshwater Biol., 33: 419-438. [ Links ]
5. Biggs, BJF; RJ Stevenson & RL Lowe. 1998. A habitat matrix conceptual model for stream periphyton. Archiv fur Hydrobiologie, 143:21-56. [ Links ]
6. Brinson, MM; AE Lugo & S Brown. 1981. Primary productivity, decomposition and consumer activity in freshwater wetlands. Annu. Rev. Ecol. Syst., 12:123-161. [ Links ]
7. Cochero, J; AM Romaní & N Gómez. 2013. Delayed response of microbial epipelic biofilm to nutrient addition in a Pampean stream. Aquat. Microb. Ecol., 69: 145-155. [ Links ]
8. Colautti, DC; ME Maroñas; ED Sendra; LC Protogino; F Brancolini ; et al. 2009. Ictiofauna del Arroyo La Choza, cuenca del Río de la Reconquista (Buenos Aires, Argentina). Biología Acuática, 26:55-62. [ Links ]
9. Colman, JA; K Sorsa; JP Hoffmann; CS Smith & JH Andrews. 1987. Yield-derived and photosynthesis-derived critical concentrations of tissue phosphorus and their signifcance for growth of Eurasian Water Milfoil, Myriophyllum-Spicatum L. Aquat. Bot., 29:111-122.
10. Croll, RP. 1983. Gastropod chemoreception. Biol. Rev., 58:293-319. [ Links ]
11. Demars, BOL & AC Edwards. 2007. Tissue nutrient concentrations in freshwater aquatic macrophytes: high inter-taxon differences and low phenotypic response to nutrient supply. Freshw. Biol., 52:2073-2086. [ Links ]
12. Dodds, WK; JR Jones & EB Welch. 1998. Suggested classifcation of stream trophic state: distributions of temperate stream types by chlorophyll, total nitrogen, and phosphorus. Water Res., 32:1455-1462.
13. Downing, JA; SB Watson & E McCauley. 2001. Predicting cyanobacteria dominance in lakes. Can. J. Fish. Aquat. Sci., 58:1905-1908. [ Links ]
14. Duarte, CM. 1992. Nutrient concentration of aquatic plants: Patterns across species. Limnol. Oceanogr., 37: 882-889. [ Links ]
15. Elosegui, A & S Sabater. 2009. Conceptos y técnicas en ecología fuvial. Rubes editorial, Girona, España.
16. Feijoó, CS; LR Leggieri; C Ocón; I Muñoz; A Rodrigues Capítulo; et al. 2014. Stoichiometric homeostasis in the food web of a chronically nutrient-rich stream. Freshw. Science, 33:820-831. [ Links ]
17. Ferreiro, N; C Feijoó; A Giorgi & L Leggieri. 2010. Effects of macrophyte heterogeneity and food availability on structural parameters of the macroinvertebrate community in a Pampean stream. Hydrobiologia, 664: 199-211. [ Links ]
18. Fitter, A & H Hillebrand. 2009. Microbial food web structure affects bottom-up effects and elemental stoichiometry in periphyton assemblages. Limnol. Oceanogr., 54:2183-2200. [ Links ]
19. Franklin, P; M Dunbar & P Whitehead. 2008. Flow controls on lowland river macrophytes: A review. Sci. Total Environ., 400:369-378. [ Links ]
20. Galat, DL; LH Fredrickson; D Humburg; KJ Bataille; JR Bodie; et al. 1998. Flooding to restore connectivity of regulated, large-river wetlands. Bioscience, 48:721-733. [ Links ]
21. Gerloff, GC. 1975. Nutritional Ecology of Nuisance Aquatic Plants. EPA, University of Wisconsin, Madison, USA. [ Links ]
22. Giorgi, A & CS Feijoó. 1999. Sedimentación de algas en arroyos de llanura. Actas VII Jornadas Pampeanas de Ciencias Naturales, 163-172. [ Links ]
23. Gómez, N; JC Donato; A Giorgi; H Guasch; P Mateo & S Sabater. 2009. La biota de los ríos: los microorganismos autótrofos. Capítulo 12. Pp. 219-242 en: Elosegi, A & S Sabater (eds.). Conceptos y técnicas en ecología fuvial. Rubes Editorial.
24. Gordon, ND; TA McMahon; BL Finlayson; CJ Gippel & RJ Nathan. 2004. Stream Hydrology. An Introduction for Ecologists, 2° edición John Wiley and Sons, Chichester, Inglaterra. [ Links ]
25. Holt, CR; D Pfitzer; C Scalley; BA Caldwell & DP Batzer. 2014. Macroinvertebrate community responses to annual fow variation from river regulation: An 11-year study. River Res. Appl., doi: 10.1002/rra.2782.
26. Jørgensen, BB & D Des Marais. 1990. The diffusive boundary layer of sediments: Oxygen microgradients over a microbial mat. Limnol. Oceanogr., 35:1343-1355. [ Links ]
27. Larned, ST; VI Nikora & BJF Biggs. 2004. Mass-transfer– limited nitrogen and phosphorus uptake by stream periphyton: A conceptual model and experimental evidence. Limnol. Oceanogr., 49:1992-2000.
28. Leggieri, LR. 2014. Funcionamiento y funcionalidad de los arroyos pampeanos: metabolismo y estequiometría. Tesis Doctoral. Universidad Nacional del Comahue, SC Bariloche, Río Negro. [ Links ]
29. Leggieri, LR; C Feijoo; A Giorgi; N Ferreiro & V Acuña. 2013. Seasonal weather effects on hydrology drive the metabolism of non-forest lowland streams. Hydrobiologia, 716:47-58. [ Links ]
30. Li, F ; H Zhang ; Y Zhu ; Y Xiao & L Chen. 2013. Effect of flow velocity on phytoplankton biomass and composition in a freshwater lake. Sci. Total Environ., 447:64-71. [ Links ]
31. Matthaei, CD; C Guggelberger & H Huber. 2003. Local disturbance history affects patchiness of benthic river algae. Freshwater Biol., 48:1514-1526. [ Links ]
32. Matthiessen, B; R Ptacnik & H Hillebrand. 2010. Diversity and community biomass depend on dispersal and disturbance in microalgal communities. Hydrobiologia, 653:65-78. [ Links ]
33. Riis, T & BJF Biggs. 2003. Hydrologic and hydraulic control of macrophytes in streams. Limnol. Oceanogr., 48:1488-1497. [ Links ]
34. Sherman, BS; IT Webster; GJ Jones & RL Oliver. 1998. Transitions between Aulacoseira and Anabaena in a turbid river weir pool. Limnol. Oceanogr., 43:1902-1915. [ Links ]
35. Soriano, A; RJC León; OE Sala; RS Lavado; VA Deregibus; et al. 1992. Río de la Plata grasslands. Capítulo 19. Pp. 367-407 en: Coupland, RT (ed.). Ecosystems of the World. Natural Grasslands: Introduction and Western Hemisphere. Elsevier, New York, USA. [ Links ]
36. Stevenson, RJ. 1996. The stimulation and drag force of current. Pp. 321-340 en: Stevenson, RJ; ML Bothwell & RL Lowe (eds.). Algal ecology; freshwater benthic ecosystems. Academic Press, San Diego, USA. [ Links ]
37. Toledo, MJ. 2011. El legado Lujanense de Ameghino: revisión estratigráfca de los depósitos pleistocenos-holocenos del valle del Río Luján en su sección tipo. Registro paleoclimático en la Pampa de los estadios Ois 4 al Ois 1. Revista de la Asociación Geológica Argentina, 68:121-167.
38. Vilches, C. 2012. Efectos de la contaminación difusa sobre el perifton de arroyos. Tesis Doctoral. Universidad Nacional de La Plata, Buenos Aires.
39. Wetzel, RG. 2001. Limnology: Lake and River Ecosystems. Academic Press, Londres, Inglaterra. [ Links ]

