










Serviços Personalizados
Journal

Artigo
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO  uBio
uBio
Compartilhar

 Permalink
PermalinkEcología austral
versão On-line ISSN 1667-782X
Ecol. austral vol.26 no.2 Córdoba ago. 2016
ARTÍCULO ORIGINAL
Comer o ser comido: factores que determinan la condición nutricional de larvas de Engraulis anchoita de la población patagónica de la especie
Marina Diaz1,2,*; Marina Do Souto1,3,4; Marina Peralta5; Marcelo Pájaro1; Mariela Spinelli3,4; Martín Saraceno6,7,8; Carlos Balestrini9 & Fabiana Capitanio3,4
1 Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP).
2 Instituto de Investigaciones Marinas y Costeras (IIMyC), Consejo Nacional de Investigaciones Científcas y Técnicas (CONICET).
3 Laboratorio de Zooplancton Marino, Departamento de Biodiversidad y Biología Experimental (DBBE), Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires (FCEN-UBA).
4 Instituto de Biodiversidad y Biología Experimental y Aplicada (IBBEA-CONICET-UBA).
5 Departamento de Ecología Genética y Evolución (EGE-FCEN-UBA).
6 Centro de Investigaciones del Mar y la Atmósfera (CIMA-CONICET-UBA).
7 Departamento de Ciencias de la Atmósfera y de los Océanos (DCAO-FCEN-UBA).
8 Unidad Mixta Internacional, Instituto Franco-Argentino de Estudios de Clima y sus Impactos (UMI-IFAECI).
9 Departamento de Oceanografía, Servicio de Hidrografía Naval.
*Editora asociada: María Diéguez mdiaz@inidep.edu.ar
Recibido: 11 de mayo de 2015 Aceptado: 18 de marzo de 2016
Resumen
La población patagónica de Engraulis anchoita habita el sector costero comprendido entre los 42° y 46° S. Sus áreas de desove y cría se asocian al frente de marea de Península Valdés, que se forma a fnes de la primavera y desaparece a principios de otoño. Las condiciones oceanográfcas del frente generan una alta productividad primaria y elevada biomasa del zooplancton asociado, potencial alimento para larvas de peces. Se estimó la condición nutricional de larvas de anchoíta por medio de tres índices: el Factor de Condición de Fulton, la relación ARN/ADN y la performance de crecimiento. Se compararon dos secciones oceanográfcas: una en la que se detectó la presencia del frente de marea y otra sin ese efecto. Los resultados de los índices evidenciaron una condición nutricional mejor en las larvas de anchoíta en la sección oceanográfca con persistencia del frente de marea. Para determinar las variables ambientales que infuencian la condición larval de E. anchoita se realizó un estudio aplicando Modelos Lineales Generalizados, empleando el índice ARN/ADN como variable dependiente y la clorofla superfcial, la abundancia de nauplii, el volumen de plancton gelatinoso y el largo estándar de las larvas como variables explicativas. Se observó una mejora de la condición nutricional con el incremento de la abundancia de nauplii de copépodos, alimento potencial de las larvas. En cambio, el índice ARN/ADN disminuyó con la abundancia de plancton gelatinoso, lo cual sugiere que en la zona estudiada, estos organismos competirían con las larvas de E. anchoita.
Palabras clave: Índice ARN/ADN; Frente de Península Valdés
Abstract
Eat or be eaten: factors that determine nutritional condition of Engraulis anchoita larvae from patagonian population of the species. The Patagonian population of Engraulis anchoita inhabits the coastal sector comprised between 42° and 46° S. Its spawning and rearing areas are associated with the tidal frontal system of Peninsula Valdés, which forms in late spring and breaks during early autumn. Oceanographic frontal conditions can enhance primary production and zooplankton biomass, representing an enriched food source for fsh larvae. Larval nutritional condition was estimated using three indices: Fulton's Condition Index, RNA/DNA ratio and growth performance. Two oceanographic sections were compared, one in which the presence of the frontal system was observed and another devoid of such effect. Condition indices refected an enhanced nutritional condition of anchovy larvae in the section where the front persisted. Generalized linear models were applied to determine the effect of environmental variables on larval condition. RNA/DNA ratio was considered as the dependent variable, whereas surface chlorophyll concentration, copepod nauplii abundance, jellyfsh volume and larval standard length were used as predictors. In general, the nutritional condition improved with the abundance of copepod nauplii, potential food source for the anchovy larvae. In contrast, the RNA/DNA ratio decreased with the increase of jellyfsh abundance, suggesting that, in the study area, these organisms might act as competitors of anchovy larvae.
Keywords: Anchovy larvae; RNA/DNA ratio; Peninsula Valdés frontal system
Introducción
El análisis de la condición nutricional permite evaluar el estado fisiológico individual de las larvas de peces y refleja la situación ambiental a la que han estado expuestas. Por lo tanto, representa un instrumento útil para determinar zonas de crianza favorables y brinda, además, herramientas para el manejo integral de poblaciones sometidas a explotación pesquera, (e.g., al aportar información que podría ser considerada para establecer áreas restringidas o períodos de veda). Existen diferentes índices para evaluar la condición nutricional en larvas de peces, tanto a nivel individual como tisular y celular (Ferron and Legget 1994). Estos índices operan en escalas temporales diferentes. El individual (o de mayor nivel de organización) brinda una información más generalizada de lo acontecido en las últimas semanas, mientras que el celular es el más sensible a los cambios ambientales en el corto plazo (una escala temporal de horas).
Uno de los índices más utilizados en las últimas décadas es el Factor de Condición de Fulton (Fulton 1904), que asume que las larvas de mayor peso a igual longitud estándar se encuentran en un estado nutricional mejor que las larvas de menor peso. Por otra parte, el índice ARN/ADN es un indicador ecofisiológico muy utilizado en la actualidad ya que brinda información acerca del estado fisiológico al proveer una medida de la capacidad de biosíntesis celular. El principio teórico para utilizar la relación ARN/ADN es considerar que el número de células, y por lo tanto la cantidad de ADN, es constante en células somáticas, mientras que el ARN presente en los tejidos es directamente proporcional a la síntesis proteica. Por tanto, el índice ARN/ADN se puede considerar como un indicador del grado de actividad metabólica de la célula; por consiguiente, resulta un indicador confiable de la condición nutricional y del crecimiento reciente en larvas de peces marinos (Clemmesen 1994; Bailey et al. 1995; Diaz et al. 2011).
La anchoíta argentina Engraulis anchoita Hubbs and Marini 1935 representa, en términos de biomasa, el recurso pesquero de mayor importancia del Atlántico Sudoccidental (Hansen 2004). Además, cumple un papel central como soporte en las redes tróficas de diversas especies de gran importancia comercial como la merluza, la caballa, la pescadilla, la anchoa de banco, así como también de aves y mamíferos marinos (Angelescu 1982; Ciechomski and Sánchez 1988). Existen al menos dos poblaciones de adultos: una bonaerense, que se encuentra al norte de los 41° S y que se reproduce durante la primavera, y otra patagónica, que se encuentra entre 41 y 47° S y que se reproduce durante el verano (Brandhorst et al. 1974; Sánchez and Ciechomski 1995). Las abundancias máximas de huevos y larvas de anchoíta pertenecientes a ambas poblaciones se encuentran asociadas a las áreas frontales presentes en el Mar Argentino (Pájaro et al. 2008). Las discontinuidades oceanográficas que caracterizan a los frentes actúan como zonas de retención de partículas y nutrientes (Carreto et al. 1986) y de concentración de huevos y de nauplii de copépodos, los cuales constituyen el principal alimento para las larvas de anchoíta (Viñas and Ramírez 1996). Por lo tanto, las áreas frontales representan importantes áreas de crianza para los estadios tempranos de desarrollo de E. anchoita.
La "Tríada Fundamental" de Bakun (Bakun 1996) indica que la supervivencia y el reclutamiento de los estadios tempranos de vida de peces pelágicos dependen de la combinación de tres procesos: i) el enriquecimiento de las capas superficiales por ascenso de aguas más profundas ricas en nutrientes, ii) la concentración de organismos planctónicos de los que se alimentan las larvas, y iii) la retención de huevos y larvas dentro de áreas favorables. Bakun y Parrish (1991) analizaron el área de distribución de E. anchoita y establecieron para el sector argentino al menos dos áreas de desove exitosas que cumplen con los postulados de la tríada de Bakun antes descripta: la zona costera bonaerense y la zona de frentes de marea norpatagónicos. El desove de la anchoíta asociado a estas áreas asegura la disponibilidad de alimento suficiente en las primeras etapas del desarrollo ontogénico de la especie (Acha et al. 2004, 2015).
El objetivo de este trabajo fue estimar la condición nutricional de las larvas de Engraulis anchoita hacia finales del verano en la región norpatagónica. Para ello se emplearon tres índices de condición a lo largo del desarrollo ontogénico de las larvas: el Factor de Condición de Fulton, el índice ARN/ADN y la performance de crecimiento. Los tres índices se compararon en dos zonas con diferentes características hidrográficas. Se estudió también la influencia de variables ambientales, de la disponibilidad de presas potenciales y de la abundancia de depredadores y/o competidores sobre la condición nutricional de las larvas de anchoíta.
Materiales y Métodos
Las muestras y datos hidrográficos analizados fueron obtenidos durante la campaña de investigación realizada en marzo de 2013 a bordo del Buque Oceanográfico ARA Puerto Deseado. El área de estudio abarca el sector costero y de plataforma media frente a Península Valdés y Rawson. Se realizaron ocho estaciones de plancton para la estimación de su composición y abundancia, en las que, además, se recolectaron larvas de E. anchoita para estudiar su condición nutricional. Dichas estaciones se situaron sobre dos transectas perpendiculares a la costa: la transecta norte (TN) y la transecta sur (TS) (Figura 1a).

Figura 1 a) Distribución espacial de las estaciones estudiadas, transecta norte (TN), transecta sur (TS). b) Perfil vertical de temperatura (°C) en la TN. c) Perfil vertical de temperatura en la TS. d) Perfil vertical de temperatura en la estación 4 (E4) de la TN. e) Perfil vertical de temperatura en la estación 8 (E8) de la TS.
Figure 1 a) Spatial distribution of the stations studied, northern (TN) and southern (TS) sections. b) Vertical temperature profile (°C) in TN. c) Vertical temperature profile in TS. d) Vertical temperature profile at station 4 (E4) in TN. e) Vertical temperature profile at station 8 (E8) in TS.
Sitio de estudio
La hidrografía de la plataforma norpatagónica está caracterizada por la formación de un sistema frontal que se produce como consecuencia de la dinámica de mareas. Esta estructura frontal se extiende desde el norte de Península Valdés hasta Bahía Camarones, siguiendo aproximadamente la isobata de 75 m (Romero et al. 2006). Este frente de mareas separa aguas costeras homogéneas de aguas estratificadas (Glorioso 1987). Las estructuras frontales no son permanentes, se forman durante la primavera y el verano. Están caracterizadas por la gran disponibilidad de nitratos propios de las zonas de alta productividad biológica ocasionada por floraciones de fitoplancton y grandes agregados de zooplancton (Acha et al. 2004, 2015). Este escenario crea una diversidad de hábitats apropiados para el desove de los peces y como áreas de crianza para sus huevos y larvas (Sánchez and Ciechomski 1995).
Datos hidrográficos
Las condiciones hidrográficas fueron determinadas a partir de las mediciones de temperatura y salinidad obtenidas en cada estación con un CTD Seabird 911. Los datos de CTD fueron procesados con el programa SeaWave provisto por la misma firma del equipo. Se compararon los perfiles descendentes con los ascendentes obtenidos en cada estación, tanto para la temperatura como para la salinidad, con el fin de detectar picos espurios que se eliminaron con un filtro pasabajos con una frecuencia de corte de 0.15 s.
Recolección de muestras
Las muestras de ictioplancton fueron colectadas con una red Bongo con malla de 300 µm de tamaño de poro, realizando lances oblicuos desde las cercanías del fondo hasta la superficie. Las larvas destinadas al análisis de la condición nutricional fueron separadas y congeladas a bordo en nitrógeno líquido, y posteriormente se conservaron a -80 °C hasta su procesamiento. Las muestras de plancton fueron fijadas en formaldehído al 5%. A partir de las muestras obtenidas con la red Bongo se cuantificaron larvas de anchoíta y organismos del plancton gelatinoso. Además, se recolectó una muestra empleando una red MiniBongo con malla de 67 µm de tamaño de poro, que luego fue analizada para cuantificar la densidad de larvas nauplii de copépodos. Empleando botellas Van Dorn se recolectaron muestras de agua para determinar la concentración de clorofila a de la fracción menor a 20 µm (Chl-a<20 µm) mediante el método fluorométrico (Holm-Hansen and Riemann 1978). Para esto se filtraron de manera secuencial 500 mL de agua por una malla de 20 µm de abertura de poro, y luego por filtro GF/F (0.7 µm).
Procesamiento de las muestras
Las larvas fueron descongeladas en el laboratorio, se las fotografió y se midió su largo estándar al micrómetro (LS), empleando el software Axio-Vision. A cada larva se le asignó un estadio de desarrollo de acuerdo a Alheit et al. (1991). De este modo se consideró como larvas en preflexión a aquellas con LS menor a 8 mm, larvas en flexión a aquellas entre 8 y 12.9 mm, y larvas en postflexión a aquellas con LS mayor a 12.9 mm. Luego, se removieron las cabezas de las larvas, y el tronco fue liofilizado durante 24 horas. Posteriormente, se determinó el peso seco (PS) con precisión de microgramo (µg) en una microbalanza Sartorius. Las muestras fueron conservadas a -80 °C hasta su procesamiento.
Para cada larva se estimó el Factor de Condición de Fulton (FCF) de acuerdo a la siguiente fórmula:
FCF=PS (1)
LS3
donde:
PS = peso seco expresado en microgramos
LS = largo estándar expresado en milímetros
Se realizó un ajuste potencial de la relación PS (µg) y LS (mm):
PS= α.LSβ (2)
donde:
α y β= constantes
Esta ecuación fue linealizada aplicando logaritmo natural a ambas variables, y se obtuvo una ecuación de crecimiento con la forma:
ln(PS)= ln.α +β. ln(LS) (3)
dónde:
β= tasa de crecimiento en peso.
Para la obtención de los índices ARN/ ADN se utilizó el protocolo descripto por Caldarone et al. (2001), modificado por Diaz y Pájaro (2012). Este procedimiento se basa en la determinación de las concentraciones de ácidos nucleicos en cada larva mediante espectroscopía de fluorescencia. En primer lugar, se procedió a la extracción de ácidos nucleicos totales mediante la lisis celular empleando buffer Sarcosil Tris EDTA. Se utilizó Bromuro de Etidio (0.1 mg/mL) como fluorocromo. Las determinaciones fueron realizadas empleando un espectrofluorómetro Perkin Elmer, excitando la muestra a 360 nm y midiendo la emisión a 590 nm (Ex360nm y Em590nm, respectivamente). Se registró inicialmente la intensidad de la fluorescencia correspondiente a los ácidos nucleicos totales, y luego se procedió al agregado de 50 µL de una ARNasa (RNase A de páncreas bovino Sigma R6513; 20 U/mL) e incubación (30 min; 42 °C) y a una segunda lectura de la fluorescencia, atribuida a la concentración de ADN. La fluorescencia residual luego del agregado de una ADNasa (Sigma D4263 y sus cofactores Mg+2 y Ca+2 correspondientes) resultó muy baja y, por lo tanto, este paso fue omitido en los procedimientos de rutina. La diferencia entre la lectura inicial y la final se consideró como la cantidad de ARN presente en cada una de las larvas.
Las concentraciones de ácidos nucleicos correspondientes a las intensidades de las fluorescencias obtenidas fueron estimadas a partir de curvas de calibración empleando concentraciones de ADN crecientes y conocidas dentro de un rango apropiado (lambda DNA, digerido con Hind III, Roche 10236250001) y ARN (RNA 16S-23S de Escherichia coli, Roche 10206938001). Esta curva se realizó diariamente, previo a la medición de fluorescencia de los extractos de tejidos de las larvas para disminuir posibles errores instrumentales asociados al ambiente. A partir de las concentraciones de ADN y ARN obtenidas para cada larva se calculó el cociente individual de ADN/ARN (RD). La relación promedio entre las pendientes de ADN y ARN de las curvas de calibración fue 2.69. Todos los valores de RD fueron estandarizados (RDs) de acuerdo al procedimiento descrito por Caldarone et al. (2006), y se consideró un valor de pendiente de referencia de 2.4. Los valores de RDs fueron utilizados para los análisis estadísticos.
A su vez, se estimó la tasa de crecimiento (G) para cada larva utilizando el modelo desarrollado por Buckley et al. (2008):
G= 0.0254 RDs-0.0037 T RDs-0.0873 (4)
donde:
G = tasa de crecimiento instantáneo
T = temperatura en el sitio de muestreo (profundidad = 3 m)
Por otro lado, con el fin de estimar el valor umbral del RDs correspondiente a la tasa de crecimiento nulo, se calculó el punto de inflexión para el cual G pasa de ser positivo a negativo.
La performance de crecimiento (Gpf), es decir el cociente entre la tasa de crecimiento observada y la tasa de crecimiento larval bajo condiciones ambientales y de alimentación óptimas (Gmax), proporciona una medida objetiva de la condición larvaria (Buckley et al. 2008). Debido a la falta de un modelo de Gmax para E. anchoita, las tasas de crecimiento de las larvas se compararon con una tasa de crecimiento de referencia (Gref) de acuerdo al modelo multiespecífico propuesto por Houde and Zastrow (1993), ajustado para 80 especies de peces marinos y estuariales criadas bajo condiciones óptimas de temperatura.
Gref=0.0106. T -0.023 (5)
De acuerdo a este modelo, los valores de Gpf mayores a uno (Gpf>1) indican una tasa de crecimiento mayor a la obtenida bajo condiciones óptimas.
Se utilizaron únicamente larvas de tallas mayores (LS) a 4 mm debido a que entre 3 y 4 mm de LS se produce la reabsorción del vitelo y comienzan a alimentarse. No se realizaron correcciones por encogimiento debido a lafalta de esta información para E. anchoita, y se asumió que el grado de encogimiento es homogéneo dentro de cada estadio de desarrollo (Theilacker 1980).
Análisis de los datos
Las pendientes de las ecuaciones LS-PS linealizadas que se obtuvieron para las larvas capturadas en las dos secciones oceanográficas contrastantes, transecta norte (TN) y transecta sur (TS), fueron comparadas mediante una prueba de t de pendientes.
Los valores de FCF, RDs y Gpf fueron comparados entre las capturas de las transectas TN y TS mediante un ANCOVA, considerando la talla de los ejemplares como covariable. También se compararon los valores de los índices mencionados en ambas transectas en función del estadio de desarrollo de las larvas. Las diferencias fueron determinadas a través de un ANOVA de dos vías, siendo los factores principales la transecta (dos niveles, TS y TN) y el estadio de desarrollo (tres niveles: preflexión, flexión y postflexión). Se realizó un análisis de correlación entre los factores transecta y estadio de desarrollo. Posteriormente al ANOVA se realizaron comparaciones múltiples mediante una prueba de Tukey (α=0.05). Para ello se utilizó el software estadístico INFOSTAT®. Para evaluar los efectos de las variables ambientales sobre el estado fisiológico de las larvas se empleó el RDs como indicador de condición debido a su rápida respuesta (en una escala de horas) frente a cambios ambientales, tal como ha sido reportado en numerosos estudios (Foley et al. 2016). Contrariamente, el FCF presenta una menor sensibilidad a los cambios ambientales y muestra cambios en una escala temporal de semanas (Ferron and Legget 1994). La Gpf representa un indicador derivado de RDs en el que se incluye la temperatura a la que se encuentran los ejemplares, variable que fue considerada como potencialmente explicativa. Por estos motivos únicamente el RDs fue seleccionado como variable dependiente para un posterior modelado mediante Modelos Lineales Generalizados (MLG).
Para este modelado, entre las variables explicativas se consideraron: transecta (factor, con niveles, TS y TN), y las covariables largo estándar, peso seco, temperatura, salinidad, hora, profundidad de la estación, densidad de larvas, abundancia de nauplii de copépodos, volumen de gelatinoso y concentración superficial de clorofila. Se partió de un modelo que contenía todas las variables y se fueron eliminando las variables no significativas (P>0.05). Se empleó el criterio de Información de Akaike (AIC) para seleccionar el modelo más adecuado (Burnham and Anderson 2002). El AIC es muy útil para comparar modelos similares con distintos grados de complejidad o modelos iguales (mismas variables) pero con funciones de enlace distintas.
Se realizó un análisis de correlación entre todas las variables potencialmente explicativas con el fin de eliminar aquellas con un alto grado de colinealidad. La variables analizadas fueron: transecta, largo estándar (mm), estadio de desarrollo, peso seco (µg), temperatura (°C), Factor de Condición de Fulton (µg/mm³), salinidad, hora, profundidad de la estación (m), densidad de larvas (ind./m2), abundancia de nauplii de copépodos (ind./m3), volumen de gelatinoso (L/m3) y concentración superficial de clorofila <20µm (µg/L). Se eliminaron las variables que presentaron correlaciones mayores a 0.80.
Se utilizó el software libre R®, versión 2.12.2 (R Development Core Team 2010). Se consideró una distribución normal de la variable dependiente y una función de enlace logarítmica. La significancia del modelo se analizó mediante un test de verosimilitud.
Resultados
Características hidrográficas del área de estudio
La situación oceanográfica detectada durante la campaña fue diferente en las transectas TN y TS (Figura 1b-e). En la transecta TN se detectó la presencia del sistema frontal típico de un frente de marea de la región caracterizado por la separación de las aguas costeras, verticalmente homogéneas, de las aguas de la plataforma que presentan una termoclina estacional (Figuras 1b y d). Por otra parte, en la transecta TS no se registró la existencia de termoclina en ninguno de los perfiles realizados, lo cual sugiere que a esa latitud se disipa el frente de mareas (Figuras 1c y e).
Relación largo-peso de las larvas de E. anchoita
La relación entre el peso seco (PS) y el largo estándar (LS) de las larvas se ajustó a un modelo potencial de crecimiento (n=139) (Figura 2):
PS= 0.0077 LS 4.56

Figura 2. Relación entre el peso seco (µg) y el largo estándar (mm) de las larvas de Engraulis anchoita y su ajuste a un modelo potencial de crecimiento. PS: peso seco, LS: largo estándar, n: número de larvas analizadas.
Figure 2. Relationship between dry weight (µg) and standard length (mm) of Engraulis anchoita larvae and their adjustment to growth potential model.
Se transformó la relación LS-PS en un modelo lineal aplicando a ambas variables logaritmo natural, dando como resultado:
ln (PS)= 4.56 ln(LS) – 4.87
No se observaron diferencias entre las pendientes de las ecuaciones obtenidas para las relaciones PS-LS linealizadas (t=1.099; P=0.273), siendo 4.29 para la transecta norte (TN) y 4.62 para la transecta sur (TS).
Índices de condición de E. anchoita
El análisis de covarianza para los valores obtenidos del Factor de Condición de Fulton (FCF) indicó diferencias significativas entre transectas (P<0.01): la condición nutricional promedio en la transecta norte fue mayor (20.43 µg/mm3) a la observada en la transecta sur (17.21 µg/mm3). Se observó una relación lineal significativa entre el FCF y LS (P<0.01) con pendiente positiva y un coeficiente de 4.60. El análisis de varianza de dos vías no mostró interacción significativa entre los factores transecta y estadio de desarrollo para el FCF (P>0.05) (Tabla 1a). Se observaron diferencias significativas entre los valores medios del FCF para los diferentes estadios de desarrollo (P<0.01) pero no entre las transectas (P>0.05) (Tabla 1). En ambas transectas el FCF medio obtenido para las larvas en postflexión fue significativamente mayor que para las larvas en estadios de desarrollo más tempranos (Tabla 1a; Figura 3a).
Tabla 1. ANOVA de dos vías y comparaciones múltiples para comparar los valores medios del Factor de Condición de a) Fulton (FCF, µg/mm3), b) índice ARN/ADN estandarizado (RDs), c) performance de crecimiento (Gpf), entre las transectas norte y sur en los diferentes estadios de desarrollo larval. ns: no significativo, *: significativo, **: altamente significativo. TN: transecta norte, TS: transecta sur. Letras mayúscula diferentes indican diferencias significativas entre transectas para cada estadio de desarrollo (P<0.05) para la prueba de ANOVA de dos vías seguida de comparaciones múltiples mediante una prueba de Tukey.
Table 1. Two-way ANOVA and post-hoc comparisons to compare mean values of a) Fulton´s condition factor (FCF, µg/mm3), b) standardized RNA/DNA ratio (RDs), c) growth performance (Gpf), between the north and south sections along larval developmental stages. ns: no statistical differences, *: statistically significant, **: extremely significant. TN: northern section, TS: southern section. Different capital letters indicate significant differences between transects for each developmental stage (P<0.05) in the two-way ANOVA followed by Tukey test.
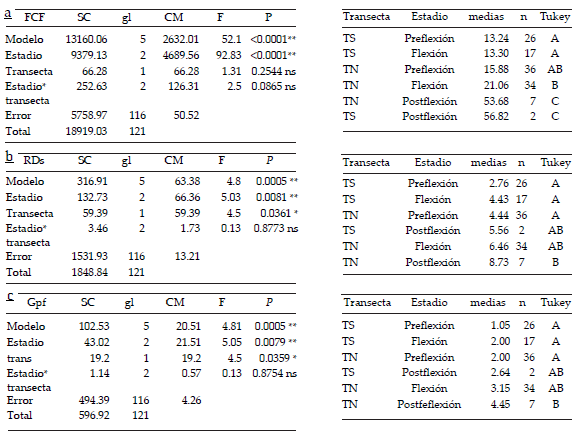

Figura 3. Índices de condición nutricional por estadio larval (preflexión, flexión, postflexión) en cada transecta (TN: transecta norte; TS: transecta sur). a) Factor de Condición de Fulton (FCF, µg/mm3). b) Índice ARN/ ADN estandarizado. c) Performance de crecimiento (Gpf). Sobre las barras se indican los valores medios de la variable.
Figure 3. Nutritional condition indexes by larval stage (preflexion, flexion and postflexion) in each section (TN: north transect; TS: south transect): a) Fulton´s condition factor (FCF), b) standardized RNA/DNA ratio (RDs), c) Growth performance (Gpf). Variables mean values are indicated above bars.
El análisis de covarianza para los valores obtenidos del índice ARN/ADN (RDs) en cada transecta resultó significativo (P<0.01), y se observó una mejor condición nutricional en las larvas de E. anchoita en la transecta norte (RDs promedio=5.51) que en las larvas de la transecta sur (RDs promedio=3.88). Se observó una relación lineal positiva significativa entre el RDs y la talla (coeficiente de correlación=0.83; P<0.01). El análisis de la varianza de dos vías aplicado al índice RDs indicó la ausencia de interacción entre los factores transecta y estadio de desarrollo para (P>0.05) (Tabla 1b). Se observaron diferencias significativas entre los valores promedio del índice RDs de los diferentes estadios de desarrollo y entre las transectas (P<0.05) (Tabla 1b). El índice RDs estimado para las larvas en postflexión de la transecta norte fue significativamente mayor que el obtenido para las larvas en preflexión de esa misma transecta y a su vez fue significativamente mayor que el correspondiente a las larvas en preflexión y en flexión de la transecta sur (P<0.001) (Tabla 1b, Figura 3b).
El valor umbral de relación RDs (tasa de crecimiento nula) obtenido para las larvas de la transecta norte fue 1.025, mientras que en el caso de la transecta sur fue 1.030. En la Figura 4 se presenta el índice RDs en función de la talla de cada uno de los individuos recolectados en las dos transectas estudiadas. La mayoría de las larvas se hallaron por encima del valor umbral del índice RDs (>1.03): en la transecta norte se encontró un 5.26% por debajo de dicho valor, mientras que en la transecta sur, un 17.78%.
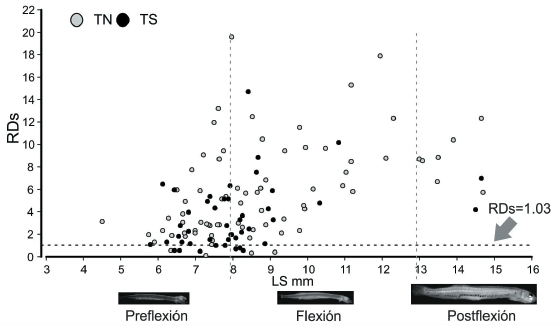
Figura 4. Índice ARN/ADN estandarizado (RDs) en función del Largo Estándar (LS) de las larvas de Engraulis anchoita pertenecientes a las transectas norte (TN) y sur (TS). La línea punteada horizontal indica el valor umbral de RDs para que tasa de crecimiento (G) sea igual a cero, utilizando el modelo RDs-T-G desarrollado por Buckley et al. (2008) y considerando la temperatura media registrada a 3 m de profundidad (16.04 °C). La línea punteada vertical indica la talla de transición entre los estadios de desarrollo larval: preflexión, flexión y postflexión.
Figure 4. Standardized RNA/DNA ratio (RDs) versus Engraulis anchoita larvae standard length (LS) from the northern (TN) and southern (TS) sections. Horizontal dashed line indicates the threshold value of RDs to growth rate (G) equal to zero, using the RDs-T-G model developed by Buckley et al. (2008) and considering the average temperature 16.04 °C. Vertical dashed line indicates the size of transition between larval developmental stages: preflexion, flexion and postflexion.
El análisis de covarianza para los valores obtenidos para la performance de crecimiento (Gpf) en cada transecta resultó significativo (P<0.01), y se observó una mejor condición nutricional en larvas colectadas en la transecta norte (Gpf promedio=2.61) que en el caso de las larvas de la transecta sur (Gpf promedio=1.68). Se halló una relación lineal positiva significativa entre el RDs y la talla (coeficiente de correlación=0.47;P<0.01). El análisis de varianza de dos vías de la Gpf reveló la ausencia de una interacción significativa entre los factores transecta y estadio de desarrollo (P>0.05) (Tabla 1c). Se observaron diferencias significativas entre los valores medios del índice RDs para los diferentes estadios de desarrollo y entre las transectas (P<0.05) (Tabla 1c). Al igual que para el RDs, la Gpf obtenida para las larvas en postflexión provenientes de la transecta norte fue significativamente mayor (P<0.001) que la obtenida para las larvas en preflexión y flexión recolectadas en la transecta sur y para las larvas en preflexión provenientes de la transecta norte (Tabla 1c; Figura 3c). Ninguno de los índices estudiados se correlacionó significativamente con la temperatura registrada en el sitio de recolección de los ejemplares (correlación de Pearson, P>0.05)
Relación entre el índice ARN/ADN de E. anchoita y variables ambientales
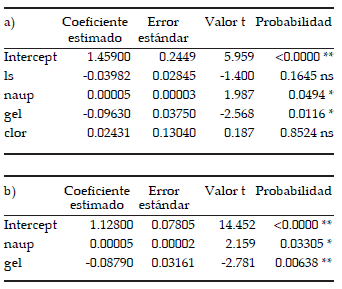
Se eliminaron del análisis las variables transecta, peso seco, temperatura, salinidad, hora y profundidad ya que presentaron correlaciones mayores a 0.80. Las variables restantes fueron removidas de a una hasta obtener un modelo con un valor del criterio de Akaike AIC=449.4, que incluyó el largo estándar, la abundancia de larvas nauplii de copépodos, el volumen de plancton gelatinoso y la concentración de clorofila.
En este modelo, las abundancias de larvas nauplii de copépodos y el volumen de plancton gelatinoso fueron las variables que resultaron estadísticamente significativas (Tabla 2a). En vista de estos resultados, se eliminó primeramente la variable clorofila, que presentó el mayor valor de probabilidad. Se corrió nuevamente el modelo y luego se eliminó la variable largo estándar. El modelo final aceptado incluyó las variables significativas abundancia de nauplii y volumen de plancton gelatinoso (Tabla 2b). Este último MLG presentó un valor menor del criterio de Akaike (AIC=447.8), en comparación con el primer modelo; esto confirmaría que el ajuste del modelo con sólo dos variables explicativas resulta apropiado.
Tabla 2. Modelo Lineal Generalizado. a) Variables empleadas: largo estándar (ls), abundancia de larvas nauplii de copépodos (naup), volumen de zooplancton gelatinoso (gel) y concentración de clorofila (clor). Criterio de Akaike (AIC)=449.43. b) Variables empleadas: abundancia de nauplii (naup) y volumen de zooplancton gelatinoso (gel). Criterio de Akaike (AIC)=447.76. ns: no significativo, *: significativo, **: altamente significativo.
Table 2. Generalized Linear Model. a) Employed variables: standard length (ls), copepod nauplii larvae abundance (naup), shellyfish volume (gel) and chlorophyll concentrations (chlor). Akaike criterion (AIC)=449.43. b) Employed variables: copepod nauplii larvae abundance (naup) and shellyfish volume (gel). Akaike criterion (AIC)=447.76 . ns: no statistical differences, *: statistically significant, **: extremely significant.
Se comprobó la normalidad de los valores de RDs y se analizó la distribución de sus residuales en función de los valores predictivos (Figura 5). En la Figura 5b se observa un leve patrón de residuales con forma de embudo, lo cual indica que éstos aumentaron a mayores valores de las variables predichas, probablemente como resultado del error del instrumento de medición debido a la cercanía de los límites de detección del mismo.
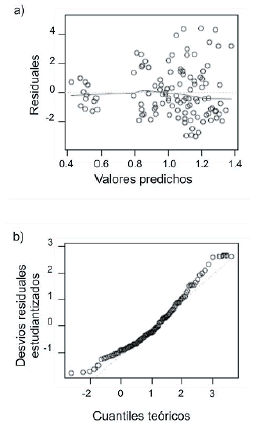
Figura 5. a) Gráfico de residuales del índice ARN/ADN estandarizado (RDs) en función de los valores predichos por el modelo. b) Gráfico Q-Q para observar la normalidad de los datos.
Figure 5. a) Residuals of standardized RNA/DNA ratio (RDs) as a function of the values predicted by the model. b) Q-Q Plot to observe the normality of the data.
El modelo aceptado mostró una relación positiva entre los valores del índice RDs y la abundancia de larvas nauplii de copépodos y una relación negativa entre los índices RDs y el volumen de plancton gelatinoso.
Discusión
Las diferencias observadas en la situación oceanográfica de las TN y la TS se deben a la época del año en la que se realizó la campaña y a las propias características físicas del frente. El frente pierde su estructura generalmente en otoño, cuando desaparece la termoclina estacional que se observaba en verano sobre la plataforma continental.
Se observó un buen estado nutricional de las larvas de Engraulis anchoita en el área de estudio en general, a pesar de que la época analizada coincidió con el final de la formación del frente de Península Valdés. Los resultados obtenidos para los tres índices de condición analizados fueron coincidentes, y se observó una condición nutricional mejor en la transecta norte en presencia del frente de marea de Península Valdés. Los mayores valores observados para los índices de condición nutricional coincidieron con las mayores abundancias de larvas nauplii de copépodos y las menores concentraciones de zooplancton gelatinoso.
Los valores medios del FCF obtenidos en este trabajo resultaron similares a los presentados por Diaz et al. (2011) para larvas de la población bonaerense. También, en forma coincidente, se observó una dependencia entre la condición nutricional y la talla de los ejemplares. Además, las tasas de crecimiento observadas en este trabajo (pendiente de las relaciones largo-peso) fueron superiores a las observadas para esta especie por Diaz et al. (2009) y Diaz y Pájaro (2013), lo que podría indicar que hacia fines del verano las condiciones ambientales en la zona estudiada serían óptimas para el crecimiento y la supervivencia de las larvas de anchoíta.
En este estudio se observaron diferencias en la condición de las larvas colectadas en distintas zonas dentro del área de crianza y a lo largo de su desarrollo ontogenético. En forma coincidente con estos resultados, Clemmesen et al. (1997), quienes estudiaron la condición nutricional de larvas de anchoíta en la misma región pero durante el comienzo del verano, advirtieron que los mayores valores del índice ARN/ADN se presentaron en las estaciones ubicadas en aguas estratificadas. En el mes de diciembre, los valores del índice ARN/ADN estuvieron por debajo de 4, mientras que en el presente trabajo fueron superiores, siendo 4.91 el valor medio del índice RDs hacia el final del verano. Cabe mencionar que los resultados obtenidos por Clemmesen et al. (1997) no fueron estandarizados, y por lo tanto las diferencias en la magnitud de la variable podrían atribuirse a cuestiones metodológicas. A su vez, los valores medios del índice RDs aquí presentados fueron mayores que los observados por Diaz et al. (2011) en el sistema frontal de El Rincón en primavera, área muy importante como zona de cría de larvas de la población bonaerense de anchoíta. Este hecho refuerza la idea de que las condiciones ambientales en el área de estudio serían óptimas para el crecimiento y la supervivencia de las larvas de E. anchoita.
Todos los índices estudiados exhibieron una dependencia con la talla de los ejemplares. Cabe destacar que las diferencias en la condición nutricional observadas entre las transectas TN y TS se registraron únicamente para las larvas de estadios tempranos de desarrollo (larvas en preflexión). Esto se debe al hecho de que a medida que las larvas se desarrollan se hacen más resistentes a la inanición, debido al incremento en la acumulación de reservas. Ante una eventual disminución de la disponibilidad de alimento, las larvas de mayor edad tolerarían las condiciones desfavorables durante más tiempo empleando sus reservas de lípidos y glucógeno (Powell and Chester 1985). Por lo tanto, cuando se estudia la incidencia de la inanición en poblaciones naturales resulta importante contemplar la edad o estadio de desarrollo de los ejemplares al momento de realizar comparaciones.
Muchos estudios sugieren que el índice ARN/ADN es uno de los mejores indicadores de la condición nutricional de diversos organismos marinos (Robinson and Ware 1988; Clemmesen 1994). En la actualidad, es el índice bioquímico más utilizado como indicador de la condición nutricional de larvas de peces. Los organismos en buena condición usualmente presentan valores más elevados de la relación ARN/ADN que aquellos que se encuentran en una condición pobre. Los valores umbrales para la relación RDs obtenidos para las transectas TN y TS fueron 1.025 y 1.030, respectivamente. Por encima de este valor umbral, las larvas presentan tasas de crecimiento positivas. Por lo tanto, se consideró en buena condición nutricional a los ejemplares que presentaron un índice RDs mayor a 1.03, mientras que aquellos por debajo de ese valor umbral presentaban una condición nutricional deficiente. En este estudio, muy pocas larvas presentaron valores bajos de condición nutricional, lo que confirma que E. anchoita encuentra en la zona estudiada las condiciones ambientales propicias para su supervivencia y el crecimiento larval.
Los primeros estadios de desarrollo de los peces viven en un universo dominado por procesos trofodinámicos definidos por las características físicas e hidrodinámicas del sistema. La depredación es generalmente un agente importante de mortalidad larval, y sus efectos están modulados por la temperatura, la abundancia de depredadores, la condición nutricional y las tasas de crecimiento larval. A su vez, estas últimas están determinadas por la disponibilidad de presas, por la diversidad y la talla de los depredadores y por la distribución de tallas de las larvas, entre muchos otros factores ambientales (Houde 2008). Por otra parte, la magnitud de todos los factores mencionados sobre el crecimiento y la supervivencia larval difiere a lo largo de la ontogenia (Clemmesen 1994). Por lo tanto, es importante tener en cuenta la distribución de tallas de las larvas al evaluar la condición larval en una zona determinada. La temperatura actúa de forma directa o indirecta sobre los procesos biológicos, y en muchos taxones explica una porción importante de la variabilidad en el reclutamiento (Houde 2009). La fisiología y actividad metabólica, el comportamiento y el crecimiento de los estadios tempranos de desarrollo de peces responden a la temperatura (Blaxter 1993). Sin embargo, en este trabajo, los índices RDs determinados no se correlacionaron con la temperatura. Buckley et al. (2008) estudiaron un modelo multiespecífico basado en temperatura e índices ARN/ADN y no encontraron una correlación entre estas dos variables cuando las larvas criadas en acuario estaban bien alimentadas. La elevada abundancia de presas en la termoclina explicaría la falta de correlación entre los índices RDs y la temperatura. En este sentido, se requerirían estudios en los que se determinen las relaciones RDs de las larvas, las abundancias de presas potenciales y los contenidos estomacales para poder determinar con certeza si el alimento representa o no un factor limitante.
Es sabido que las larvas de peces que cuentan con una disponibilidad de presas adecuada (en términos de calidad y cantidad) presentan una mayor tasa de crecimiento y una condición nutricional privilegiada. El hecho de que las larvas de anchoíta presenten una buena condición nutricional, reflejada en los altos valores del índice de condición, podría ser una evidencia de que dentro del área de estudio el alimento no resulta un factor limitante para el crecimiento y la supervivencia larval, aun en áreas con grandes concentraciones larvales y elevada competencia intraespecífica.
Al evaluar los parámetros potencialmente explicativos de los valores de RDs en cada estación, el MLG arrojó como variables significativas la abundancia de nauplii y el volumen de organismos pertenecientes al plancton gelatinoso. La relación de la abundancia de nauplii con la condición larval fue positiva. Los nauplios de las especies pequeñas de copépodos constituyen las presas más frecuentes, seguidos por los huevos de copépodos. Sin embargo, las larvas de E. anchoita también consumen en menor proporción tintínidos, dinoflagelados, larvas de moluscos y diatomeas (Viñas and Ramírez 1996; Sato et al. 2011). Zenitani et al. (2007) realizaron una experiencia de acuario empleando larvas de E. japonicus alimentadas exclusivamente con larvas nauplii de copépodos. Estos autores observaron el incremento de la supervivencia larval (%/ día) conforme aumentaban las densidades de larvas nauplii, llegando a una asíntota de aproximadamente 89%/día a densidades mayores a 5000 nauplii/m3. En el presente trabajo, las densidades promedio de larvas nauplii de copépodos fueron cercanas a 5000 ind/m3 en la TN, donde se registró la mejor condición de las larvas de E. anchoita. A pesar de que las densidades de larvas nauplii fueron significativamente menores en la TS, la condición nutricional de las larvas de anchoíta fue buena, lo que indicaría que probablemente se alimentarían también de otras presas, además de las larvas nauplii de copépodos.
La segunda variable significativa, el volumen de cnidarios y ctenóforos de cada estación, presentó una relación negativa con la condición nutricional de E. anchoita. El plancton gelatinoso es considerado un depredador o competidor de las larvas de peces según la disponibilidad de otras presas en el ambiente (Alvariño 1985; Purcell 1985, 1990, 1991; Chícharo 1998a, 1998b; Purcell and Arai 2001; Purcell et al. 2001). La competencia entre el plancton gelatinoso y E. anchoita podría ser de tipo directa por el alimento, o bien por el espacio, ya que una abundancia elevada de zooplancton gelatinoso dificultaría la obtención del alimento de las larvas de E. anchoita, disminuyendo así su condición nutricional. En la transecta TS se observó menor abundancia de larvas nauplii de copépodos y mayor densidad de plancton gelatinoso, lo cual evidenciaría el efecto de competencia interespecífica por el alimento. La combinación de una baja concentración de presas y una mayor concentración de posibles competidores podría causar la reducción de la condición nutricional, fenómeno observado en la transecta TS, donde la estructura frontal no se encontró presente.
Dentro del área analizada, la transecta TN respondería a los postulados de la tríada de Bakun, ya que la estructura frontal garantiza la estabilidad de la columna de agua debido a la estratificación vertical, el enriquecimiento de nutrientes y la retención de los productos del desove dentro de un hábitat favorable. Esta transecta muestra densidades elevadas de larvas nauplii de copépodos y larvas de anchoíta, y también una mejor condición nutricional y de crecimiento de estas últimas (RDs>1, Gpf>1).
Por otra parte, para estimar la importancia de la inanición en el océano es necesario establecer calibraciones de laboratorio y determinar el valor crítico de la relación ARN/ADN por debajo del cual las larvas se encontrarían en inanición o próxima a ella. Sin embargo los índices ARN/ADN aquí obtenidos fueron superiores a los valores críticos de este índice reportados en la bibliografía, que típicamente se encuentran entre 1 y 3 (Ferron and Leggett 1994; Foley et al. 2016). En particular para los clupeiformes, los índices ARN/ADN fluctúan entre 1 y 1.3 (Clemmesen 1994; Chícharo 1997; Kimura et al. 2000), dependiendo de la temperatura y el estadio de desarrollo, entre otros factores.
La importancia de este tipo de estudios radica en que permiten determinar la existencia de zonas favorables para el crecimiento y la supervivencia de las larvas. Por otra parte, aportan información que puede ser considerada para establecer áreas restringidas y períodos de veda apropiados. En consecuencia, constituyen herramientas para el manejo integral de poblaciones sometidas a explotación pesquera.
Si bien E. anchoita constituye un recurso poco explotado en la actualidad, presenta una gran potencialidad. Debido a la situación pesquera mundial y regional, esta especie podría ser explotada de manera más intensa en el futuro. Por otra parte, en 2011 se concretó la certificación de la pesquería de la población bonaerense de anchoíta (Acta 31/2011 del Consejo Federal Pesquero), y en la actualidad se discute su importancia como especie piloto para aplicar el enfoque ecosistémico en el manejo de la pesquería, hechos que refuerzan la necesidad de profundizar los conocimientos sobre esta especie.
Agradecimientos
Los autores expresan su gratitud a las autoridades del INIDEP y al personal del Buque Oceanográfico ARA Puerto Deseado por toda la colaboración brindada para la obtención del material aquí empleado. A ML Presta y N Curcio por la colaboración durante la campaña. Este trabajo fue financiado por el INIDEP y por los siguientes proyectos: EUMETSAT/CNES DSP/OT/12-2118 (M. Saraceno), CONICET-YPF PIO 133-20130100242 (M. Saraceno); PIP CONICET 11220110100351CO y UBACYT 2013-2015 (F. Capitanio). Contribución INIDEP nº 1947. Agradecemos todos los comentarios y sugerencias realizados por dos revisores anónimos y a los editores ya que han contribuido a una versión considerablemente mejorada del manuscrito.
Referencias
1. Acha, E. M., H. W. Mianzan, R. A. Guerrero, M. Favero, and J. Bava. 2004. Marine fronts at the continental shelves of austral South America Physical and ecological processes. J Marine Syst 44:83-105. [ Links ]
2. Acha, E. M., A. R. Piola, O. O. Iribarne, and H. W. Mianzan. 2015. Ecological Processes at Marine Fronts. Oases in the Ocean. Springer, London, England. Pp. 73. [ Links ]
3. Alheit, J., J. D. Ciechomski, L. Djurfeldt, C. Ebel, M. D. Ehrlich, J. C. Elgue, G. Mantero, Y. Matsuura, H. W. Mianzan, W. Nellen, C. Oderbrecht, F. C. Ramírez, R. P. Sánchez, G. Shaffer, and M. D. Viñas. 1991. SARP Studies on Southwest Atlantic anchovy, Engraulis anchoita, off Argentina, Uruguay and Brasil. ICES Biological Oceanography Committee 46. Pp. 80. [ Links ]
4. Alvariño, A. 1985. Las surgencias en la región de California-Baja California: Relaciones con el zooplancton y poblaciones de Engraulis mordax (pisces). Invest Mar CICIMAR 2:81-102. [ Links ]
5. Angelescu, V. 1982. Ecología trófca de la anchoíta del Mar Argentino (Engraulidae, Engraulis anchoita). Parte II. Alimentación, comportamiento y relaciones trófcas en el ecosistema. Contrib. Contrib. INIDEP (Mar del Plata) Nro. 409. Pp. 83. [ Links ]
6. Bailey, K. M., M. F. Canino, J. N. Napp; S. M. Spring, and A. L. Brown. 1995. Contrasting years of prey level, feeding conditions, and mortality of larval walleye Pollock Theragra chalcogramma in the western Gulf of Alaska. Mar Ecol Prog Ser 119:11-23. [ Links ]
7. Bakun, A. 1996. Patterns in the ocean. Ocean processes and marine population dynamics. University of California Sea Grant, California, USA, in cooperation with Centro de Investigaciones Biológicas de Noroeste, La Paz, Baja California Sur, México. Pp. 323. [ Links ]
8. Bakun, A., and R. H. Parrish. 1991. Comparative studies of coastal pelagic fsh reproductive habitats: The anchovy (Engraulis anchoita) of the southwestern Atlantic. ICES J Mar Sci 48:343-361. [ Links ]
9. Blaxter, J. H. S. 1993 The effect of temperature on larval fsh. Neth J Zool 42:336-357. [ Links ]
10. Brandhorst, W., J. P. Castello, M. B. Cousseau, and D. A. Capezzani. 1974. Evaluación de los recursos de anchoíta (Engraulis anchoita) frente a Argentina y Uruguay. VIII. Desove, crecimiento, mortalidad y estructura de la población. Physis, Sec. A 33(86):37-58. [ Links ]
11. Buckley, L. J., E. M. Caldarone, and C. Clemmesen. 2008. Multi-species larval fsh growth model based on temperature and fuorometrically derived RNA/DNA ratios: results from a meta-analysis. Mar Ecol Prog Ser 371:221-232. [ Links ]
12. Burnham, K. P., and D. R. Anderson. 2002. Model selection and multimodel inference: a practical information-theoretic approach. 2nd ed. Springer, New York. Pp. 515. [ Links ]
13. Caldarone, E. M., M. Wagner, J. St. Onge-Burns, and L. J. Buckley. 2001. Protocol and guide for estimating nucleic acids in larval fsh using a fuorescence microplate reader. National Marine Fisheries Service, 166 Water Street, Woods Hole, MA 02543-1026. Pp. 22. [ Links ]
14. Caldarone, E. M., M. C. Clemmesen, E. Berdalet, T. J. Miller, A. Folkvord, G. J. Holt, M. P. Olivar, and I. M. Suthers. 2006. Intercalibration of four spectrofuorometric protocols for measuring RNA:DNA ratios in larval and juvenile fsh. Limnol Oceanogr Methods 4:153-163. [ Links ]
15. Carreto, J. I., H. R. Benavides, R. M. Negri, and P. D. Glorioso. 1986. Toxic red-tide in the Argentine Sea. Phytoplankton distribution and survival of the toxic dinofagellate Gonyaulax excavata in a frontal area. J Plankton Res 8:15-28. [ Links ]
16. Chícharo, M. A. 1997. Starvation percentages in feld caught Sardina pilchardus larvae off southern Portugal. Sci. Mar. 61:507-516. [ Links ]
17. Chícharo, M. A., L. M. Chícharo, L. Valdez, E. López-Jamar, and P. Re. 1998a. Does the nutritional condition limit survival potential of sardine Sardina pilchardus (Walbaum 1792) larvae off the north coast of Spain? RNA/DNA ratios and their variability. Fish. Res. 39:43-54. [ Links ]
18. Chícharo, M. A., L. M. Chícharo, L. Valdez, E. López-Jamar, and P. Re. 1998b. Estimation of starvation and diel variation of the RNA/DNA ratios in feld-caught Sardina pilchardus larvae off the north of Spain. Mar Ecol Prog Ser 164:273-283. [ Links ]
19. Ciechomski, J., and R. P. Sánchez. 1988. Análisis comparativo de las estimaciones de biomasa de la anchoíta (Engraulis anchoita) en el Atlántico Sudoccidental en diferentes años y con distintas metodologías. Frente Marit 4:117-131. [ Links ]
20. Clemmesen, C. 1994. The effect of food availability, age or size on the RNA/DNA ratio of individually measured herring larvae: laboratory calibration. Mar Biol 118:377-382. [ Links ]
21. Clemmesen, C.M., R. P. Sánchez, and C. Wongtschowski. 1997. A regional comparison of nutritional condition of SW Atlantic anchovy larvae (Engraulis anchoita) based on RNA/DNA ratios. Arch Fish Mar Res 45:17-43. [ Links ]
22. Diaz, M. V., and M. Pájaro. 2012. Protocolo para la determinación de ácidos nucleicos en larvas de peces. Inf Invest INIDEP 20:1-9. [ Links ]
23. Diaz, M. V., y M. Pájaro. 2013. Estudio de la condición nutricional de larvas de la población bonaerense de anchoíta (Engraulis anchoita) en relación con las características hidrográfcas del área de crianza. Rev Invest Des Pesq 23:107-127. [ Links ]
24. Diaz, M. V., M. Pájaro, M. P. Olivar, P. Martos, and G. J. Macchi. 2011. Nutritional condition of Argentine anchovy, Engraulis anchoita, larvae in connection with nursery ground properties. Fish. Res. 109:330-341. [ Links ]
25. Diaz, M. V.; M. Pájaro, and R. P. Sánchez. 2009. Employment of morphometric variables to assess nutritional condition of Argentine anchovy larvae Engraulis anchoita Hubbs and Marini. 1935. Rev Biol Mar Oceanogr 44:539-549. [ Links ]
26. Ferron, A., and W. C. Leggett. 1994. An appraisal of condition measures for marine fsh larvae. Adv Mar Biol 30: 217-303. [ Links ]
27. Foley, C. J., D. L. Bradleyc, and T. O. Hööka. 2016. A review and assessment of the potential use of RNA:DNA ratios to assess the condition of entrained fsh larvae. Ecol. Indic. 60:346-357. [ Links ]
28. Fulton, T. W. 1904. The rate of growth of fishes. Ann. Rept. Fish. Board Scotland 22(3):141-241. [ Links ]
29. Glorioso, P. 1987. Temperature distribution related to shelf-fronts on the Patagonian Shelf Cont Shelf Res 7:27-34. [ Links ]
30. Hansen, J. E. 2004. Anchoíta (Engraulis anchoita). Pp 101-115 en: R. P. Sánchez, S. I. Bezzi and E. E. Boschi (eds.). El mar argentino y sus recursos pesqueros. Publicaciones especiales INIDEP, Mar del Plata. Argentina. [ Links ]
31. Holm-Hansen, O., and B. Riemann. 1978. Chlorophyll a Determination: Improvements in Methodology. Oikos 30(3): 438-447. [ Links ]
32. Houde, E. D. 2008. Emerging from Hjort's Shadow. J Northwest Atl Fish Soc 41:53-70. [ Links ]
33. Houde, E. D. 2009. Recruitment variability. Ch. 3. Pp. 91-171 en: T. Jakobsen, M. Fogarty, B. A. Megrey and E. Moksness (eds.). Fish Reproductive Biology and Its Implications for Assessment and Management. Wiley-Blackwell, Oxford. [ Links ]
34. Houde, E. D., and C. E. Zastrow. 1993. Ecosystem -and taxon- specifc dynamic and energetics properties of larval fsh assemblages. Bull Mater Sci 54:290-335. [ Links ]
35. Kimura, R., Y. Watanabe, and H. Zenitani. 2000. Nutritional condition of frst-feeding larvae of japanese sardine in the coastal and oceanic waters along the Kuroshio Current. ICES J Mar Sci 57:240-248. [ Links ]
36. Pájaro, M., P. Martos, E. Leonarduzzi, G. J. Macchi, M. V. Díaz, and D. Brown. 2008. Estrategia de puesta de la Anchoíta (Engraulis anchoita) en el Mar Argentino y Zona Común de Pesca Argentino-Uruguaya. Inf Téc Of INIDEP 11:1-14. [ Links ]
37. Powell, A., and A. J. Chester. 1985. Morphometric indices of nutritional condition and sensitivity to starvation of spot larvae. Trans Amer Fish Soc 114:338-347. [ Links ]
38. Purcell, J. E. 1985. Predation on fsh eggs and larvae by pelagic cnidarians and ctenophores. Bull Mar Sci 37:739-755. [ Links ]
39. Purcell, J. E. 1989. Predation on fsh larvae and eggs by the hydromedusa Aequorea victoria at a herring spawning ground in British Columbia. Can J Fish Aquat Sci 46:1415-1427. [ Links ]
40. Purcell, J. E. 1990. Soft-bodied zooplankton predators and competitors of larval herring (Clupea harengus pallasi) at herring spawning grounds in British Columbia. Can J Fish Aquat Sci 47:505-515. [ Links ]
41. Purcell, J. E., and M. N. Arai. 2001. Interactions of pelagic cnidarians and ctenophores with fsh: a review. Hydrobiologia 451:27-44. [ Links ]
42. Purcell, J. E., T. A. Shiganova, M. B. Decker, and E. D. Houde. 2001. The ctenophore Mnemiopsis in native and exotic habitats: U.S. estuaries versus the Black Sea basin. Hydrobiologia 451:145-176. [ Links ]
43. R: A language and environment for statistical computing. 2010. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0. URL: http://www.R-project.org. [ Links ]
44. Robinson, S. M., and D. Ware. 1988 Ontogenetic development of growth, rates in larval Pacifc herring, Clupea harengus pallasi, measured with RNA/DNA ratios in the Strait of Georgia, British Columbia. Can J Fish Aquat Sci 45:1422-1429. [ Links ]
45. Romero, S. I., A. R. Piola, M. Charo, and C. A. Eiras García. 2006. Chlorophyll-a variability off Patagonia based on SeaWiFS data. J Geophys Res C. 111 C05021, doi: 10.1029/2005JC003244. [ Links ]
46. Sánchez, R. P., and J. D. Ciechomski. 1995. Spawning and nursery grounds of pelagic fsh species in the sea-shelf off Argentina and adjacent areas. Sci Mar 59:455-478. [ Links ]
47. Sato, N. E., D. Hernández, and M. D. Viñas. 2011. Hábitos alimentarios de las larvas de Engraulis anchoita (Hubbs and Marini 1935) en las aguas costeras de la Provincia de Buenos Aires, Argentina. Lat Am J Aquat Res 39:16-24. [ Links ]
48. Theilacker, G. H. 1980. Changes in body measurements of larval northern anchovy, Engraulis mordax, and other fshes due to handling and preservation. Fish Bull 78:685-692. [ Links ]
49. Viñas, M. D., and F. C. RAMÍREZ. 1996. Gut analysis of frst-feeding anchovy larvae from Patagonian spawning area in relation to food availability. Arch Fish Mar Res 43:231-256. [ Links ]
50. Zenitani, H., N. Kono, and Y. Tsukamoto. 2007. Relationship between daily survival rates of larval Japanese anchovy (Engraulis japonicus) and concentrations of copepod nauplii in the Seto Inland Sea, Japan. Fish Oceanogr 16:473-478. [ Links ]

