










Services on Demand
Journal

Article
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkEcología austral
On-line version ISSN 1667-782X
Ecol. austral vol.26 no.2 Córdoba Aug. 2016
ARTÍCULO ORIGINAL
Biological colonization and leaf decomposition in a subtropical stream
Cristiane Biasi*; Cristina Cerezer &Sandro Santos
Programa de Pós-Graduação em Biodiversidade Animal, Departamento de Ecologia & Evolução, Centro de Ciências Naturais e Exatas, Universidade Federal de Santa Maria. Santa Maria, Rio Grande do Sul, Brasil.
*Editora asociada: Diéguez María cristiane.biasi@gmail.com
Recibido: 30 de octubre de 2015 Aceptado: 7 de mayo de 2016
Abstract
In small streams, aquatic community obtains energy mainly from leaves of the riparian vegetation. Processing these leaves involves physical, chemical and biological factors that may difer among leaf species. We assessed the efects of litter quality on leaf decomposition and colonization by aquatic invertebrates in a subtropical stream. Leaves from two native, highly represented tree species common in subtropical riparian areas, Ficus luschnathiana and Gasearía sylvestris, were incubated in coarse mesh bags in a stream. Bags were sampled weekly during an incubation of 28 days. The decomposition rates were higher in leaves of F. luschnathiana than in those of C. sylvestris. In the frst seven days of the experiment, leaves of F. luschnathiana lost ~60% of the initial mass, while Cascarla sylvestris lost around 30%. The leaves of C. sylvestris were initially more palatable than the leaves of F. luschnathiana, perhaps due to their lower lignin and cellulose content, and lignin to N ratio (Lignin:N). These structural compounds enhance leaf toughness thereby protecting them from herbivores and detritivores. However, leaves of F. luschnathiana were comparatively softer, which apparently facilitated fungal colonization and conditioning. Thus, leaves of F. luschnathiana attained high shredders density, which was refected in a higher leaf mass loss. Our results suggest that litter decomposition was sensitive to fungal biomass and leaf toughness. We observed that microbial conditioning, rather than the initial chemical quality of the leaf litter, decreased leaf toughness thereby stimulating further colonization by shredders and enhancing the decomposition process. The fungal community has a key-role in the degradation process within aquatic environments, breaking down even low-quality litter (i.e., F. luschnathiana) and promoting changes in the composition and structure of the invertebrate community.
Keywords: Shredders; Chironomidae; Leaf-toughness; Fungal biomass
Resumen
La colonización y descomposición biológica de la hojarasca en un arroyo subtropical. En arroyos de bajo orden, la comunidad acuática obtiene energía principalmente del aporte de la hojarasca que proviene de la vegetación de la ribera. El procesamiento de las hojas implica medios físicos, químicos y factores biológicos que pueden diferir entre especies vegetales. En esta investigación se evaluó el efecto de la calidad del sustrato sobre el proceso de descomposición y sobre la colonización por invertebrados acuáticos asociados, en un arroyo subtropical. Las hojas de dos especies de árboles nativos, Ficus luschnathiana y Cascarla sylvestris, se incubaron en bolsas de malla gruesa en un arroyo durante 28 días, colectándose muestras semanalmente. Las tasas de descomposición fueron mayores en F. luschnathiana que en C. sylvestris. En los primeros siete días del experimento, F. luschnathiana perdió ~60% de la masa inicial, mientras que C. sylvestris perdió ~30%. Las hojas de C. sylvestris fueron inicialmente más palatables que las de F. luschnathiana, tal vez debido a su menor contenido de lignina y celulosa, y a su más baja relación Lignina:N. Estos compuestos estructurales le conferen rigidez a las hojas, las protegen de los herbívoros y las hacen más resistentes a los detritívoros. Sin embargo, la menor dureza de la hoja de F. luschnathiana facilitó la colonización fúngica, lo cual se refejó, a su vez, en una mayor colonización por invertebrados trituradores y, en general, una mayor pérdida de masa. Los resultados sugieren que la descomposición de la hojarasca fue sensible a la biomasa fúngica y a la dureza de las hojas. Se observó que la colonización microbiana y la disminución de la dureza de las hojas tienen un efecto más fuerte sobre la colonización por invertebrados trituradores y sobre el proceso de descomposición que la calidad química inicial de la hojarasca. La comunidad fúngica es clave en los procesos de degradación en el medio acuático, ya que actúa en la descomposición de la hojarasca, aun en aquella de baja calidad (i.e., F. luschnathiana), promoviendo cambios también en la composición y en la estructura de la comunidad de invertebrados colonizadores.
Palabras clave: Trituración; Chironomidae; Dureza foliar; Biomasa fúngica
Introduction
In low-order streams in which light limitation restricts primary production, the allochthonous material largely support biotic communities and ecological processes (Vanotte et al. 1980; Webster and Benfield 1986). Once the plant debris enter into streams, organic matter and nutrients leach out while leaves travel downstream or remain on the bank, providing resources for detritivores and decomposers (Pettit et al. 2012). These processes overall reduce the size of the particulate organic matter, releasing nutrients that support aquatic communities (Tanaka et al. 2006). The decomposition dynamic may be affected by abiotic and biotic factors such as climate (Bruder et al. 2013), water flow (Fonseca et al. 2013), quantity, quality and diversity of leaf litter (Ferreira et al. 2012; Pettit et al. 2012; Biasi et al. 2013; König et al. 2014), the abundance of shredders (Patrick 2013), among other factors operating on spatial and temporal scales (Pérez-Harguindeguy et al. 2000; Bastian et al. 2007).
The physical and chemical attributes of leaves vary substantially among species, affecting its utilization by microorganisms and shredders (Wright and Covich 2005; Bastian et al. 2007). Further, certain litter types attract different invertebrate assemblages (Schädler and Brandl 2005). Leaf litter with high nitrogen content, low secondary compounds (i.e., polyphenols and tannins), low levels of structural compounds (i.e., lignin and cellulose) and softer is considered of high quality (Graça et al. 2001). Such features determine also that the leaf litter be more attractive to the organisms, thus enhancing its decomposition rate. Low-quality leaf litter is therefore comparatively more stable and long-lasting, being consumed afterwards (Haapala et al. 2001; Graça and Canhoto 2006). In addition, understanding the effects of leaf chemical composition on the decomposition process is important due to the fact that it may reflect on nutrient cycling (Leroy and Marks 2006).
Leaves colonized by microorganisms become more palatable to leaf-shredding macroinvertebrates, which are within the main functional feeding group in low-order streams (Vannote et al. 1980). Leaf conditioning during breakdown reduce leaf toughness (Graça and Zimmer 2005), resulting in maximum leachate of nitrogen (N), protein and phosphorus (P) (Bärlocher 1985; Suberkropp 1992; Graça et al. 1993) and enhancing palatability to shredders.
Feeding and abundance of shredders have been correlated with increased leaf litter conditioning and with fungal biomass on leaf litter (Chergui and Pattee 1993; Graça et al. 1993; Robinson et al. 1998), raising the question of what the proximate cues are for shredder's food preference (Aßmann et al. 2011). Shredders participate actively in nutrient cycling in freshwater; they convert coarse particulate organic matter into to fine particles (CPOM and FPOM, respectively) absorbing partly carbon and other nutrients in the process (Boyero et al. 2014). In the subtropics the community of shredders is composed by a few taxa (e.g., Cogo and Santos 2013; König et al. 2014).
In this investigation we evaluated the effect of litter quality in terms of lignin cellulose and nitrogen contents on the decomposition and invertebrate colonization of two leaf litter types, Ficus luschnathiana and Casearia sylvestris, in a subtropical stream. We expect leaves bearing higher concentrations of lignin and cellulose that provide rigidity will protect and/or reduce the action of detritivores. On the contrary, high nitrogen concentration in leaf litter that stimulates the activity of fungi will turn the leaf litter more attractive to invertebrates increasing its decomposition rate. Through experimental incubations of leaf litter two hypothesis were tested: i) leaf decay rates are determined by the chemical composition of the detritus and thus, soft and highly nutritious leaf litter will decompose more rapidly than hard leaves, and ii) shredders will select for more nutritious leaf litter.
Materials and Methods
Study area
The decomposition experiment was conducted in a first-order stream located at 29°39'04'' S - 53°31'51'' W in the Planalto and Central Depression areas in a subtropical forest region of southern Brazil (Castillero 1984). The climate of the region is subtropical with annual mean temperature around 19 °C and mean annual rainfall ~1500 mm. Tree diversity in this subtropical forest is relatively high (Cogo and Santos 2013). The riparian vegetation is composed by several tree species, mainly Cabralea canjerana (Vell.) Mart. (deciduous), Campomanesia xanthocarpa O. Berg. (semi-deciduous), Casearia sylvestris Sw. (semi-deciduous), Cupania vernalis Cambess. (semi-deciduous), Enterolobium contortisiliquum (Vell.) Morong (deciduous), Erythrina falcata Benth. (deciduous), Ficus luschnathiana (Miq.) Miq. (deciduous), Inga alata Benoist (evergreen), Ocotea puberula (Rich.) Nees (evergreen), Parapiptadenia rigida (Benth.) Brenan (evergreen), Phytolacca dioica L. (deciduous), Rollinia emarginata Schltdl. (deciduous) and Trema micrantha (L.) Blume (evergreen).
The stream generally flows year-round, but may eventually dry during extremely hot and dry summers (especially in January and February). It has well preserved riparian vegetation on both sides and low human intervention. During the study period mean temperature was moderate 19.8±0.4 °C, the water showed mean oxygen concentration of 7.9±0.8 mg/L, a mean conductivity of 22.37±5.96 mS/cm and pH averaging 6.1±0.1. The average width of the stream was 73±20cm, the mean depth was 43±8cm and the current velocity averaged 0.3±0.0 m/s.
Leaf sampling and chemical analyses
The species used in the experiment were Ficus luschnathiana and Casearia sylvestris. These species were chosen because they are locally abundant and all of them are native to the biome known as "Mata Atlantica". Senescent leaves of both species were collected along the stream banks in October 2011 (spring) and dried in an oven at 30 °C until constant weight.
The initial chemical quality of the litter was characterized before starting the experiment through measurement of leaf toughness, amount of nitrogen, lignin and cellulose levels. A penetrometer (Nolen and Pearson 1993) was used to determine leaf toughness (expressed as kgf/cm2, kilogram-force per square centimeter) by puncturing five points of ten leaves from each species. The total nitrogen content was determined applying the Kjeldahl method (Flindt and Lillebo 2005). Lignin and cellulose percentages were determined through the fiber acid-detergent method following Van Soest (1993).
Leaf decomposition
A total of 32 mesh bags (15x20 cm, 10 mm pore size) containing 4.0±0.1 g of leaf litter which were prepared to perform the experimental incubations in the stream. The experiment was set up with two treatments, one consisting on leaves of C. sylvestris and the second one with leaves of F. luschnathiana. The litter bags were placed in the stream at sites with moderate water flow.
Four litter bags from each treatment were randomly removed from the stream during the incubation period at day 7, 14, 21, and 28. Replicates were packed in plastic bags and transported refrigerated to the laboratory. The samples were rinsed in a 0.25 mm mesh sieve to remove sediment and the associated invertebrates. The invertebrates were identified to the lowest possible taxonomic level, using identification guides (Fernández and Domínguez 2001; Merritt et al. 2008; Mugnai et al. 2010). In order to identify the Chironomidae, semi-permanent slides were prepared using Hoyer's solution and the larvae were identified to genus level using the identification guides by Trivinho-Strixino (2011) and Epler (2001). The taxa were classified in trophic groups according to Cummins et al. (2005), Wantzen and Wagner (2006) and Merritt et al. (2008).
Fungal biomass
Fungal biomass was estimated indirectly through the concentration of ergosterol, a natural constituent of most fungal cells that has been used previously as a proxy of the aquatic fungal biomass (Gessner 2005). For this purpose, 10 leaf discs (12 mm) were obtained from each replicate which were maintained at -20 °C until the extraction protocol was applied. The ergosterol was extracted applying the Microwave Assisted Extraction method (Young 1995) and was quantified by high-performance liquid chromatography (HPLC) by injecting 10 µL during 8 minutes. The mobile phase was 100% methanol and the detection wavelength was 282 nm (Gessner 2005). The HPLC system was equipped with a LiChroCART 250-4 LiChrospher 100 RP-18 (5 mm) column (Merck, Darmstadt, Germany) Results were expressed as µg fungal biomass/g dry mass. The leaves were dried in an air-circulation oven at 60 °C during 72 h to determine the remaining dried mass.
Data analysis
The resulting values of the remaining dry mass (Wt) were fitted to a negative exponential model following Webster and Benfeld (1986):
Wt = W0.e-kt (1)
where W0=initial dry mass, k=breakdown constant and t=incubation time.
For each experimental treatment, the total density of organisms and the density of shredders were calculated after Gotelli and Colwell (2001). The results obtained of remaining dry mass, leaf toughness and macroinvertebrate colonization were compared by means of a two-way analysis of variance (Two-way ANOVA), using leaf litter type (2 levels) and incubation time (3 levels) as main factors. The lignin, cellulose and nitrogen content of the leaf litter types were compared by means of t-tests. Invertebrate density values were log-transformed (log[x+1]). All statistical analyses were conducted using soft-ware R (R Development Core Team 2012).
Results
Chemical Quality and Leaf Breakdown
The leaves of C. sylvestris and F. luschnathiana differed in their lignin, cellulose and N contents, and also showed contrasting Lignin: N ratios and leaf toughness (Table 1). The C. sylvestris showed lower lignin and cellulose content, lower Lignin:N ratio and higher N level as compared to F. luschnathiana. The latter showed lower initial leaf toughness (Table 1). The leaf toughness decreased with the incubation time and resulted higher in C. sylvestris throughout the experiment (Species: GL=1, F=3.94, P=0.0001; Time: GL=4, F=8.10, P=0.0001;Species×Time:GL=4,F=1.11, P=0.0986) (Figure 1). The remaining dry mass differed between the species and among incubation times (P=0.0005; Litter type: GL=1, F=28.52, P=0.0001; Time: GL=4, F=113.41, P=0.0001;Littertype×Time:GL=4,F=4.09).
Table 1. Percentage of lignin, cellulose, nitrogen, lignin to nitrogen ratio and leaf toughness in leaves of Ficus luschnathiana and Casearia sylvestris.
Tabla 1. Porcentaje de la lignina, celulosa, nitrógeno y dureza de las hojas de Ficus luschnathiana y Casearia sylvestris.
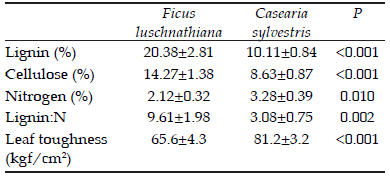

Figure 1. Changes in the toughness (kgf/cm2) of Casearea sylvestris and Ficus luschnathiana leaf litter during the decomposition experiment, at day 7, 14, 21 and 28 of incubation in a subtropical stream.
Figura 1. Cambios en la dureza de la hojarasca (kgf/cm2) de Casearea sylvestris y Ficus luschnathiana durante el experimento de descomposición, a los 7, 14, 21 y 28 días de incubación en un arroyo subtropical.
In the first seven days of the experiment, F. luschnathiana lost approximately 60% of the initial mass, while C. sylvestris lost around 30% (Figure 2). F. luschnathiana showed higher breakdown rates than C. sylvestris (k=0.238±0.022 1/d and k=0.090±0.91 1/d, respectively).

Figure 2. Percentage of remaining leaf litter mass of Casearea sylvestris and Ficus luschnathiana during the decompositíon experiment, at day 7, 14, 21 and 28 of the incubation time in a subtropical stream.
Figura 2. Porcentaje de peso remanente de hojarasca de Casearea sylvestris y Ficus luschnathiana durante el experimento de descomposición, a los 7, 14, 21 y 28 días de incubación en un arroyo subtropical.
Biological colonization
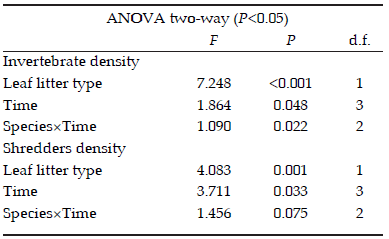
The total density of macroinvertebrates varied significantly between leaf litter types (Table 2) and it was dependent on the incubation time. The highest macroinvertebrate density was found in F. luschnathiana at day 14 of the experimental incubation. The density of shredders differed between leaf species and also among incubation times (Table 2).
Table 2. Results of the Two way ANOVA applied to study the effect of litter type, incubation time and the interaction between these variables on invertebrate and shredders densities (individuals/g dry mass) in experiment of decomposition. d.f.=degrees of freedom.
Tabla 2. Resultados del ANOVA de dos vías aplicado para estudiar el effecto de la especie de hojarasca, el tempo de incubación y su interacción sobre la abundancia de invertebrados y trituradores (individuos/g de materia seca) en el experimento de descomposición. d.f.=grados de libertad.
Gathering-collector s were the most numerous group (44%), followed by filtering-collectors (25%), scrappers (12.4%), shredders (9.6%) and predators (9%). The shredders group were represented by Grypopterigidae, Tipulidae, Calamoceratidae (Phylloicus), Hyallelidae (Hyallela) and the chironomids Endotribelos, Stenochironomus, Cricotopus and Rheocricotopus. The inclusión chironomids increased ~ 7.5% the abundance of shredders. F. luschnathiana had the highest abundance of shredders (73%), especially after 14 days of incubation (Table 3). The leaves of F. luschnathiana showed higher concentrations of ergosterol during the experiment as compared to C. sylvestris (Figure 3) and both leaf types showed maximum concentration of this compound at day 14 of the incubation.
Table 3. Total abundance of the macroinvertebrate taxa colonizing experimental enclosures with leaf litter from Ficus luschnathiana and Casearea sylvestris at different incubation times (7, 14, 21 and 28 days) in subtropical stream.
Tabla 3. Abundancia total de los taxones de macroinvertebrados que colonizan las clausuras con hojas de Ficus luschnathiana y Casearea sylvestris en las incubaciones experimentales (días 7, 14, 21 y 28) en un arroyo subtropical.
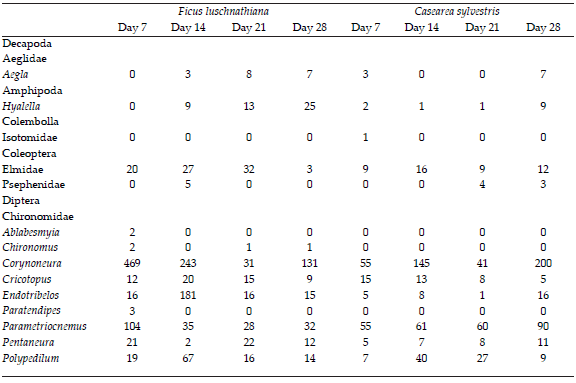
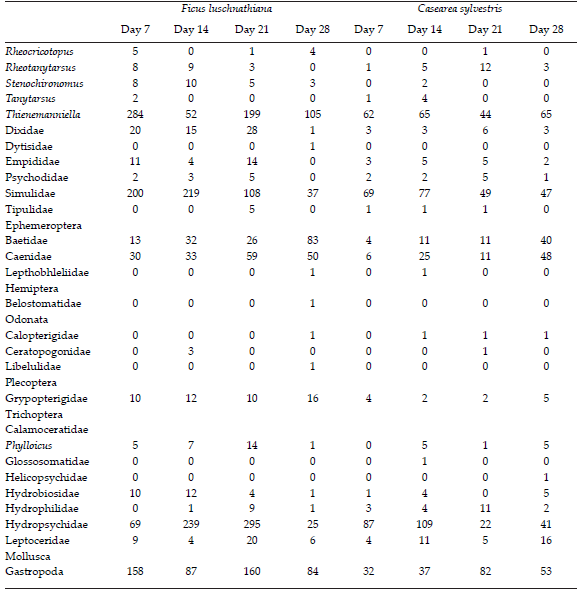

Figure 3. Changes in the concentration of ergosterol (ug/g DM) in leaf litter of Casearea sylvestris and Ficus luschnathiana in the decomposition experiment, at day 7, 14, 21 and 28 of the incubation time in a subtropical stream.
Figura 3. Concentración de ergosterol (µg/gDM) en la hojarasca de Casearea sylvestris y Ficus luschnathiana durante el experimento de descomposición, a los 7, 14, 21 y 28 días de incubación en un arroyo subtropical.
Dlscussion
Chemical Quality and Leaf Breakdown
C. sylvestris and F. luschnathiana present different values as food resource in terms of their chemical quality. Based on Lignin: N ratios, lignin and cellulose concentrations, C. sylvestris leaf litter can be considered as a higher quality food resource compared to the leaf litter of F. luschnathiana, although the leaves of the latter are softer.
The breakdown rates of F. luschnathiana and C. sylvestris were classified as rapid based on Petersen and Cumming (1974), who proposed that decomposition rates can be rapid (k>0.01), intermediate (0.005< k<0.001) or slow (k<0.005), based on the remaining dry mass percentage. Decomposition processes in streams may relate also with temperature and prevailing hydrological conditions, such as the water velocity and the slopes of the banks (Hepp et al. 2008; König et al. 2014). The decomposition rates recorded in the present analysis are higher than those reported in other studies performed in subtropical streams (Biasi et al. 2013; Cogo and Santos 2013; König et al. 2014). Compared to these studies in which temperature was moderate (17.6 °C; 13.5 °C; 14.2 °C, respectively), the temperature during our experimental incubations was on average 4.7 °C higher (19.8 °C). Temperature seasonality can be an important factor determining breakdown rates. The higher temperatures characteristic of the warmer season probably enhanced the activity of the microbial community associated to the leaves thus promoting faster breakdown rates (e.g., Pascoal and Cássio 2004; Ferreira et al. 2006).
F. luschnathiana leaf litter showed the largest mass loss. The leaching recorded in the first days of incubation was apparently the main factor explaining the rapid decomposition. During this process, many hydrosoluble compounds, such as proteins, amino acids, carbohydrates, and lipids are leached, accelerating the loss of leaf mass. The water velocity in the stream (~0.3 m/s) may have contributed to the leaching observed in two species. According to Fonseca et al. (2013), a faster current increases the mass loss during leaching, favoring the transport of fine particulate and dissolved organic matter, and even enhancing the loss of refractory material.
The decomposition rates were not related to the initial quality of the litter. We did not observe an increase in the decomposition rate in litter of C. sylvestris that showed a high nutritional quality. However, we observed a high mass loss at low leaf toughness. Recent studies in tropical streams have shown that leaf toughness rather than N and P concentrations controls litter breakdown rates (Ardón et al. 2009; Li et al. 2009). This leaf trait is related with rigid leaves protected from herbivory, and consequently also from detritivores. The leaf toughness can be related negatively with fragmentation and leaf decomposition (Foucreau et al. 2013). Some studies have also reported no clear relationship between litter initial chemical quality and decomposition (Moore and Fairweather 2006; Swan and Palmer 2004; Lecerf et al. 2007; Abelho 2009; Schindler and Gessner 2009). Different species may have particular chemical composition; the leaves may be changed by the immersion in the stream and the leaching, which overall alter their chemical quality (Pettit et al. 2012).
Biological colonization
We observed a high density of shredders colonizing F. luschnathiana leaf litter. This species, considered "a priori" as a low quality litter, showed higher mass loss and higher fungal biomass than C. sylvestris. Although of nutritionally low quality in terms of nitrogen, F. luschnathiana showed the highest fungal biomass during all the experiments, which likely produced labile detritus enhancing its palatability and favoring consumption by detritivores (Graça and Cressa 2010). The breakdown of herbivore-inhibiting compounds by aquatic fungi may favor assimilation of elements by invertebrates (Graça et al. 2001). Although leaf litter of F. luschnathiana can be considered of lower quality compared to C. sylvestris, the former has lower leaf toughness. This leaf trait has been considered as an important factor affecting leaf breakdown, since it is related with the resistance to physical abrasion and fragmentation (Ratnarajah and Bermuta 2009; Fonseca et al. 2013). A decrease in leaf toughness has been associated with higher densities of shredders (Foucreau et al. 2013). As the fungal community establishes the resistance of leaves decreases, thereby reducing leaf toughness (Aßmann et al. 2011). Thus, litter conditioning and leaf toughness reduction may be recall to explain the higher rates of decomposition found in leaf litter of F. luschnathiana.
Chironomidae was the most numerous group during the entire decomposition process, comprising alone 7.5% of the shredders. The genus Endotribelos increased shredders density, likely contributing to enhance leaf decomposition, as has been shown in other related studies (Janke and Trivinho-Strixino 2007; Chará-Serna et al. 2012; Leite-Rossi and Trivinho-Strixino 2012). Similarly, chironomids have been recorded as important components of the invertebrate communities attaining high abundance and richness. These insects play an important role in nutrient recycling (Oertli 1993; Grubbs et al. 1995; Callisto et al. 2007; Hepp et al. 2008) since many of them are generalists and colonize different types of detritus, regardless of its quality and intervening in its breakdown (Moretti et al. 2007; Landeiro et al. 2008; Li et al. 2009; Biasi et al. 2013). In certain cases, chironomids may participate actively in shredding and eroding the leaves, utilizing this detritus as a food resource (Rosemond et al. 1998). However, since chironomids have a wide range of food habits at the genus and species level, their role in leaf fragmentation may be underestimated. The results presented here highlight the importance of chironomids in the fragmentation of detritus. As chironomid shredders are present at high densities in association with detritus, they should be considered co-participants of the decomposition process in tropical aquatic systems.
The dominance of collector-gatherers and collector-filterers may relate to litter conditioning by fungi and shredders that release fine particulate matter and dissolved organic matter, facilitating colonization by these groups. Collector-gatherers and collector-filterers, are not able to directly participate in the rapid breakdown of leaves. They use leaf litter as substratum and leaf fragments as a food resource. Dudgeon and Wu (1999) suggested that collector-filtering organisms, mainly Simuliidae, use leaves as a substratum, which may explain their occurrence in large numbers in our experimental litter bags.
The shredders, represented mainly by Hyalella, Grypopterigidae, Phylloicus, Endotribelos, Polypedilum and Cricotopus were more numerous in enclosures with leaf litter of F. luschnathiana. The density of shredders was determined by the litter species and degradation time. Several studies have recorded a low occurrence of shredders in tropical streams (Wantzen and Wagner 2006; Gonçalves et al. 2007; Moretti et al. 2007), pointing to the microbial community as the primary contributor to the decomposition of leaves. Although chironomids can produce much of the fragmentation as observed here, most studies have underestimated their potential role in the decomposition process. Furthermore, Aegla, a macroconsumer that can attain high densities, could also contribute to the fragmentation of coarse organic-matter (Cogo and Santos 2013).
The incubation time of detritus is important for invertebrate colonization as has been also observed by Abelho (2001), Leroy and Marks (2006) and Ligeiro et al. (2010). Many studies evaluating invertebrate colonization of decomposing debris have observed a steady increase in density from the beginning towards the end of the experiment (Dudgeon 1982; Webster and Befield 1986; Benstead 1996; Gonçalves et al. 2012). Apparently, the leaf fragmentation process and the release of nutrients create an appropriate substratum for the development of the insects that use detritus in their diet and over time increase its quality, through microbial conditioning. The changes in the community during incubation time may relate to chemical and microbial conditioning, which together induce changes in the detritus making it more attractive to macroinvertebrates (Abelho 2001; Graça et al. 2001). This result suggests that shredders are able to discriminate advanced stages of leaf decomposition, which is consistent with the observation of Ligeiro et al. (2010) on leaf colonization by shredders. It may be possible that the ergosterol content of tough leaves increases while toughness decreases during the conditioning process. Our results also suggest that F. luschnathiana is a resource that becomes palatable depending on its residence time in the stream, during which leaf toughness is reduced by chemical and biological conditioning.
Our results suggest that leaf litter decomposition was driven by fungal biomass and leaf toughness. Contrary to the initial hypothesis, we observed that the microbial colonization and the decrease of leaf toughness rather than the initial chemical quality, determine the colonization by shredders and the decomposition process. Conditioning by the fungal community is particularly relevant to the overall decomposition process, since it breaks down low-quality litter, as in the case of F. luschnathiana leaf litter. On the whole, this study indicates that leaf physical traits such as toughness may be more important than leaf chemical traits, thereby driving decomposition rates.
Acknowledgments
We thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and the Conselho Nacional de Desenvolvimento Científco e Tecnológico (CNPq) for the scholarship and productivity grant to C.B. and S.S., respectively. Our thanks are also extended to our colleagues from the Laboratório de Carcinologia for their assistance during the feld experiments, and to Dr. Miguel Durlo for allowing us to conduct the experiments in his property.
References
1. Abelho, M. 2001. From litterfall to breakdown in streams: a review. Sci World 1:656-680. [ Links ]
2. Abelho, M. 2009. Leaf-litter mixtures affect breakdown and macroinvertebrate colonization rates in a stream ecosystem. Internat. Rev Hydrobiol 94:436-451. [ Links ]
3. Aßmann, C., K. Rinke, J. Nechwatal, and E. V. V. Elert. 2011. Consequences of the colonisation of leaves by fungi and oomycetes for leaf consumption by a gammarid shredder. Freshwater Biol 56:839-852. [ Links ]
4. Ardón, M, C. M. Pringle, and S. L. Eggert. 2009. Does leaf chemistry differentially affect breakdown in tropical vs temperate streams? Importance of standardized analytical techniques to measure leaf chemistry. Journal of the North American Benthological Society 28:440-453. [ Links ]
5. Bärlocher, F. 1985. The role of fungi in the nutrition of stream invertebrates. Bot J Linean Soc 91:83-94. [ Links ]
6. Bastian, M., L. Boyero, B. R. Jackes, and R. G. Pearson. 2007. Leaf litter diversity and shredder preferences in an Australian tropical rain-forest stream. J Trop Ecol 23:219-229. [ Links ]
7. Benstead, J. P. 1996. Macroinvertebrates and the processing of leaf litter in a tropical stream. Biotropica 28:367-375. [ Links ]
8. Biasi, C., A. M. Tonin, R. M. Restello, and L. U. Hepp. 2013. Colonisation of leaf litter by Chironomidae (Diptera): infuence of chemical quality and exposure duration in a Neotropical stream. Limnologica 43:427-433. [ Links ]
9. Boyero, L., B. J. Cardinale, M. Bastian, and R. G. Pearson. 2014. Biotic vs. Abiotic Control of Decomposition: A Comparison of the Effects of Simulated Extinctions and Changes in Temperature. Plos One 1:1-6. [ Links ]
10. Bruder, A., M. H. Schindle, M. S. Moretti, and M. O. Gessner. 2013. Litter decomposition in a temperate and a tropical stream: the effects of species mixing, litter quality and shredders. Freshwater Biol 59:438-449. [ Links ]
11. Callisto, M., J. F. Gonçalves Jr, and M. A. S. Graça. 2007. Leaf litter as a possible food source for chironomids (Diptera) in Brazilian and Portuguese headwater streams. Rev Brasil Zool 24:442-448. [ Links ]
12. Castillero, A. C. 1984. Uso da terra por fotografas aéreas no município de Santa Maria, RS. Monografa de Especialização. Universidade Federal de Santa Maria, Santa Maria. [ Links ]
13. Chara-Serna, A. M., J. D. Chara, M. C. Zuniga, R. G. Pearson, and L. Boyero. 2012. Diets of leaf litter-associated invertebrates in three tropical streams. Ann Limnol - Int J Lim 48:139-144. [ Links ]
14. Chergui, H., and E. Pattee. 1993. Fungal and invertebrate colonization of Salix fresh and dry leaves in a Moroccan river system. Archiv Fuür Hydrobiologie 127:57-72. [ Links ]
15. Cogo, G. B., and S. Santos. 2013. The role of aeglids in shredding organic matter in Neotropical streams. J Crustacean Biol 34:519. [ Links ]
16. Covich, A. P., M. C. Austen, F. Bärlocher, E. Chauvet, B. J. Cardinale, et al. 2004. The role of biodiversity in the functioning of freshwater and marine benthic ecosystems. Bio Science 54:767-775. [ Links ]
17. Cummins, K. W., R. W. Merritt, and P. C. N. Andrade. 2005. The use of invertebrate functional groups tocharacterize ecosystem attributes in selected streams and rivers in south Brazil. Stud Neotrop Fauna Environ 40:69-89. [ Links ]
18. Dudgeon, D. 1982. An investigation of physical biotic processing of two species of leaf litter in Tai Po Kau forest stream, New Territories, Hong Kong. Arch Hydrobiol 64:1-35. [ Links ]
19. Dudgeon, D., and K. K. Y. Wu. 1999. Leaf litter in a tropical stream: food or substrate for macroinvertebrates? Arch Hydrobiol 146:65-82. [ Links ]
20. Epler, J. 2001. Identification manual for the larval Chironomidae (Diptera) of North and South Carolina. Orlando, Departament of Enviromental and Natural Resources. Pp. 378. [ Links ]
21. Ferreira, V., A. C. Encalada, and M. A. S. Graça. 2012. Effects of litter diversity on decomposition and biological colonization of submerged litter in temperate and tropical streams. Freshwater Sci 31:945-962. [ Links ]
22. Ferreira, V., M. A. S. Graça, J. L. M. P. De Lima, and R. Gomes. 2006. Role of physical fragmentation and invertebrate activity in the breakdown rate of leaves. Archiv Hydrobiol 165:493-513. [ Links ]
23. Fernández, H. R., and E. Domínguez. 2001. Guía para La determinación de los artrópodos bentônicos Sudamericanos. Universidad Nacional de Tucumán, Tucumán, Argentina. Pp. 282. [ Links ]
24. Flindt, M. R., and A. I. Lillebo. 2005. Determination of total nitrogen and phosphorus in leaf litter. Pp. 53-59 in: M. A. S. Graça, F. Bärlocher and M. O. Gessner (eds.). Methods to Study Litter Decomposition: A Practical Guide. Springer, Dordrecht. [ Links ]
25. Foucreau, N., S. Puijalon, F. Hervant, and C. Piscart. 2003. Effect of leaf litter characteristics on leaf conditioning and on consumption by Gammarus pulex. Freshwater Biol 58:1672-1681. [ Links ]
26. Fonseca, A. L. S., J. R. I. Bianchini, C. M. M. Pimenta, C. B. P. Soares, and N. Mangiavacchi. 2013. The fow velocity as driving force for decomposition of leaves and twigs. Hydrobiologia 703:59-67. [ Links ]
27. Gonçalves Jr, J. F., M. A. S. Graça, and M. Callisto. 2007. Litter decomposition in a Cerrado savannah stream is retarded by leaf toughness, low dissolved nutrients and a low density of shredders. Freshwater Biol 52:1440-1451. [ Links ]
28. Gonçalves Jr, J. F., R. S. Rezende, N. M. Martins, and R. S. Gregório. 2012. Leaf breakdown in an Atlantic Rain Forest stream. Austral Ecol 37:807-815. [ Links ]
29. Gotelli, N. J., and R. K. Colwell. 2001. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett 4:379-391. [ Links ]
30. Graça, M. A. S., and C. Canhoto. 2006. Leaf litter processing in low order streams. Limnetica 25:1-10. [ Links ]
31. Graça, M. A. S., C. Cressa, M. O. Gessner, M. J. Feio, K. A. Callies, and C. Barrios. 2001. Food quality, feedingpreferences, survival and growth of shredders from temperate and tropical streams. Freshwater Biol 46:1-11. [ Links ]
32. Graça, M. A. S., and C. Cressa. 2010. Leaf quality of some tropical and temperate tree species as food resource for stream shredders. Internat Rev Hydrobiol 95:27-41. [ Links ]
33. Graça, M. A. S., and M. Zimmer. 2005. Leaf toughness. Pp. 121-125 in: M. A. S. Graça, F. Bärlocher and M. O. Gessner (eds.). Methods to Study Litter Decomposition: A Practical Guide. Springer, Dordrecht. [ Links ]
34. Graça, M. A. S., L. Maltby, and P. Calow.1993. Importance of fungi in the diet of Gammarus pulex and Asellus aquaticus. Effects on growth, reproduction and physiology. Oecologia 96:304-309. [ Links ]
35. Grubbs, S. A., R. E. Jacobsen, and K. W. Cummins. 1995. Colonization by Chironomidae (Insecta, Diptera) on distinct leaf substrates in an Appalachian mountain stream. Ann Limnol 31:105-118. [ Links ]
36. Haapala, A., T. Muotka, and A. Markkola. 2001. Breakdown and macroinvertebrate and fungal colonization of alder, birch, and willow leaves in a boreal forest stream. JNABS 20:395-407. [ Links ]
37. Hepp, L. U., C. Biasi, S. V. Milesi, F. O. Veiga, and R. M. Restello. 2008. Chironomidae (Diptera) larvae associated to Eucalyptus globulus and Eugenia unifora leaf litter in a Subtropical stream (Rio Grande do Sul, Brazil). Acta Limnol Brasil 22:345-350. [ Links ]
38. Janke, H., and S. Trivinho-Strixino. 2007. Colonization of leaf litter by aquatic macroinvertebrates: a study in a low order tropical stream. Acta Limnol Brasil 19:109-115. [ Links ]
39. König, R., L. U. Hepp, and S. Santos. 2014. Colonisation of low- and high-quality detritus by benthicmacroinvertebrates during leaf breakdown in a subtropical stream. Limnologica 45:61-68. [ Links ]
40. Landeiro, V. L., N. Hamada, and A. S. Melo. 2008. Responses of aquatic invertebrate assemblages and leaf breakdown to macroconsumer exclusion in Amazonian ‘terra frme' streams. Fund Appl Limnol 172:49-58.
41. Lecerf, A., G. Risnoveanu, C. Popescu, M. O. Gessner, and E. Chauvet. 2007. Decomposition of diverse litter mixtures in streams. Ecology 88:219-227. [ Links ]
42. Leite-Rossi, L. A., and S. Trivinho-Strixino. 2012. Are sugar cane leaf-detritus well colonized by aquatic macroinvertebrates? Acta Limnol Brasil 24:303-313. [ Links ]
43. Leroy, C. J., and J. C. Marks. 2006. Litter quality, stream characteristics and litter diversity infuence decomposition rates and macroinvertebrates. Freshwater Biol 51:605-617. [ Links ]
44. Li, A. O. Y., L. C. Y. Ng, and D. Dudgeon. 2009 Effects of leaf toughness and nitrogen content on litterbreakdown and macroinvertebrates in a tropical stream. Aquat Sci 71:80-93. [ Links ]
45. Ligeiro, R., M. S. Moretti, J. R. Gonçalves Jr, and M. Callisto. 2010. What is more important for invertebrate colonization in a stream with low-quality litter inputs: exposure time or leaf species? Hydrobiologia 654:125-136. [ Links ]
46. Merritt, R. W., R. W. Cummins, and M. B. Berg. 2008. An introduction to the aquatic insects of North America. Dubuque. Kendal/Hunt. USA. Pp. 1214. [ Links ]
47. Moore, T. N., and P. G. Fairweather. 2006. Decay of multiple species of seagrass detritus is dominated by species identity, with an important infuence of mixing litter. Oikos 114:329-337. [ Links ]
48. Moretti, M. S., J. F. Gonçalves Jr, and M. Callisto. 2007. Leaf breakdown in two tropical streams: Differences between single and mixed species packs. Limnologica 37:250-258. [ Links ]
49. Mugnai, R., J. L. Nessimian, and D. F. Baptista. 2010. Manual de identifcação de macroinvertebrados aquáticos do Estado do Rio de Janeiro. Rio de Janeiro: Technical Books Editora. Pp. 174. [ Links ]
50. Nolen, J. A., and R. G. Pearson. 1993. Factors affecting litter processing by Anisocentropus kirranus (Trichoptera: Calamoceratidae) from an Australian tropical rainforest stream. Freshwater Biol 29:469-479. [ Links ]
51. Oertli, B. 1993. Leaf litter processing and energy fow through macroinvertebrates in a woodland pond (Switzerland). Oecologia 96:466-477. [ Links ]
52. Pascoal, C., and F. Cássio. 2004. Contribution of fungi and bacteria to leaf litter decomposition in a polluted river. Appl Environm Microbiol 70:5266-5273. [ Links ]
53. Patrick, C. J. 2013. The effect of shredder community composition on the production and quality of fne particulate organic matter. Freshwat Sci 32:1026-1035. [ Links ]
54. Pérez-Harguindeguy, N., S. Díaz, J. H. C. Cornelissen, F. Vendramini, M. Cabido, and A. Castellanos. 2000. Chemistry and toughness predict leaf litter decomposition rates over a wide spectrum of functional types and taxa in central Argentina. Plant and Soil 218:21-30. [ Links ]
55. Petersen, R. C., and K. W. Cummins. 1974. Leaf processing in a woodland stream. Freshwater Biol 4:343-368. [ Links ]
56. Pettit, N. E., T. Davies, J. B. Fellman, P. F. Grierson, D. M. Warfe, and P. M. Davies. 2012. Leaf litter chemistry, decomposition and assimilation by macroinvertebrates in two tropical streams. Hydrobiologia 680:63-77. [ Links ]
57. R, Development Core Team. 2012. R: A Language and Environment for Statistical Computing, Reference Index Version 2.15.2. R Foundation for Statistical Computing, Vienna, Austria. [ Links ]
58. Ratnaraja, L., and L. A. Barmuta. 2009. The effects of leaf toughness on feeding preference by two Tasmanian shredders. Hydrobiologia 636:173-178. [ Links ]
59. Robinson, C. T., M. O. Gessner, and J. V. Ward. 1998. Leaf breakdown and associated macroinvertebrates in alpine glacial streams. Freshwater Biol 40:215-228. [ Links ]
60. Rosemond, A. D., C. M. Pringle, and A. Ramírez. 1998. Macroconsumer effects on insect detritivores and detritus processing in a tropical stream. Freshwater Biol 39:515-523. [ Links ]
61. Schädler, M., and R. Brandl. 2005. Do invertebrate decomposers affect the disappearance rate of litter mixtures? Soil Biol Biochem 37:329-337. [ Links ]
62. Schindler, M. H., and M. O. Gessner. 2009. Functional leaf traits and biodiversity effects on litter decomposition in a stream. Ecology 90:1641-1649. [ Links ]
63. Suberkropp, K. 1992. Interactions with invertebrates. Pp. 118-134 in: H. Remmert, F. B. Golley, W. D. Billings, O. L. Lange and J. S. Olson (eds.). The Ecology of Aquatic Hyphomycetes. Springer-Verlag, Heidelberg & New York. [ Links ]
64. Swan, C. M., and M. A. Palmer. 2004. Leaf diversity alters litter breakdown in a Piedmont stream. JNABS 23:15-28. [ Links ]
65. Tanaka, M. O., A. C. A. Ribas, and A. L. T. Souza. 2006. Macroinvertebrate succession during leaf litter breakdown in a perennial karstic river in Western Brazil. Hydrobiologia 568:493-498. [ Links ]
66. Trivinho-Strixino, S. 2011. Larvas de Chironomidae: guia de identifcação. Departamento de Hidrobiologia/Laboratório de Entomologia Aquática. UFSCar, São Carlos. Pp. 229. [ Links ]
67. Vannote, R. L., G. W. Minshall, K. W. Cummins, J. R. Sedell, and C. E. Cushing. 1980. The river continuum concept. Can J Fish Aquat Sci 37:130-137. [ Links ]
68. Van Soest, P. J. 1993. Use of detergents in the analysis of fbrous feeds. II. A rapid method for the determination of fbre and lignin. J Assoc of Ana Chem 46:829-835. [ Links ]
69. Wantzen, K. M., and R. Wagner. 2006. Detritus processing by invertebrate shredders: a neotropical temperate comparison. JNABS 25:216-232. [ Links ]
70. Webster, J. R., and E. F. Benfeld. 1986. Vascular plant breakdown in freshwater ecosystems. Annu Rev Ecol Syst 17: 567-594. [ Links ]
71. Wright, M. S., and A. P. Covich. 2005. Relative importance of bacteria and fungi in a tropical headwater stream: leaf decomposition and invertebrate feeding preference. Microbial Ecol 49:536-546. [ Links ]
72. Young, J. C. 1995. Microwave-assisted extraction of the fungal metabolite ergosterol and total fatty acids. J Agric Food Chem 43:2904-2910. [ Links ]

