










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.27 no.1 Córdoba ene. 2017
ARTÏCULOS
Assessing the effects of urbanization on streams in Tierra del Fuego
Jean-Paul A. Zagarola1 ; Guillermo Martínez Pasteur2; María Eugenia Lopez2 & Christopher B. Anderson2,3,*
1 Bonneville Environmental Foundation, Portland, OR, USA.
2 Centro Austral de Investigaciones Científicas (CADIC), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Ushuaia, Argentina.
3 Instituto de Ciencias Polares, Ambiente & Recursos Naturales, Universidad Nacional de Tierra del Fuego, Ushuaia, Argentina.
* canderson@cadic-conicet.gob.ar
Editora asociada: María Diéguez
Recibido: 17 de agosto de 2016
Aceptado: 27 de diciembre de 2016
Abstract
We set out to understand how urbanization affects streams in Ushuaia, Tierra del Fuego. Paired t-tests and linear regressions were used to compare physico-chemical stream habitat variables (i.e., temperature, turbidity, conductivity, dissolved oxygen and pH) and benthic macroinvertebrate community structure (i.e., density, taxonomic richness, Shannon-Weiner diversity) and function (functional feeding groups [FFG]) in four watersheds with urban and reference sites. We then calculated indices of biotic integrity for habitat (rapid visual assessment protocol [RVAP]) and benthos (Ephemeroptera, Plecoptera, Trichoptera richness [EPT], family biotic index [FBI], rapid bioassessment protocol [RBP] and biotic monitoring Patagonian streams [BMPS]). Results indicated that urbanization negatively impacted these streams’ underlying ecological condition, decreasing benthic biodiversity and dissolved oxygen, while increasing conductivity and turbidity. FFG assemblage was similar between both sites, but urbanization increased the existing dominance of collector-gatherers. Additionally, urban sites presented lower values in three of the benthic macroinvertebrate indices (EPT, RBP, BMPS, but not FBI) and also for habitat (RVAP). These data fill an existing gap in stream ecology for southern Patagonia and are useful to create monitoring tools. By incorporating urbanization as a driver of ecosystem change, managers and planners will be better able to confront the issue of sustainable development in this region, which is considered one of the most pristine wilderness areas remaining on the planet but whose human population is concentrated in a few densely populated urban areas.
Keywords: Biomonitoring; Macroinvertebrates; Management of natural resource; Urban ecology; Watersheds
Resumen
Evaluación de los efectos de la urbanización en los arroyos de Tierra del Fuego. Nos propusimos entender cómo la urbanización afecta a los ríos y arroyos en Ushuaia, Tierra del Fuego. Para esto se utilizaron pruebas de t apareadas y regresiones lineales con el fin de comparar variables físico-químicas (i.e., temperatura, turbidez, conductividad, oxígeno disuelto y pH) con la estructura y función (i.e., densidad, riqueza taxonómica, diversidad de Shannon-Weiner, grupos funcionales alimentarios [FFG, en inglés]) de la comunidad de macroinvertebrados bentónicos en cuatro cuencas con sitios urbanos y de referencia. Luego, se calcularon los índices de integridad biótica para el hábitat (rapid visual assessment protocol [RVAP]) y el bentos (riqueza de Ephemeroptera, Plecoptera, Trichoptera [EPT], family biotic index [FBI], rapid bioassensment protocol [RBP] y biotic monitoring Patagonian streams [BMPS]). Los resultados indicaron que la urbanización afectó negativamente la condición ecológica subyacente de estas cuencas, disminuyendo la biodiversidad bentónica y el oxígeno disuelto, mientras que la conductividad y la turbidez aumentaban. El ensamble de FFG fue similar entre ambos sitios, pero la urbanización incrementó la dominancia existente de organismos colectores-recolectores. Además, los sitios urbanos presentaron valores más bajos en tres de los índices de macroinvertebrados bentónicos (EPT, RBP, BMPS, pero no FBI) y también para el hábitat (RVAP). Estos datos llenan un vacío existente en la ecología de los arroyos para el sur de la Patagonia, y son útiles para crear herramientas de monitoreo. Mediante la incorporación de la urbanización como factor de cambio en los ecosistemas, los gestores y planificadores estarán en mejores condiciones para hacer frente a la cuestión del desarrollo sostenible en esta región, que se considera uno de los espacios naturales más prístinos que queda en el planeta, pero cuya población humana se concentra en unas pocas áreas urbanas densamente pobladas.
Palabras clave: Biomonitoreo; Macroinvertebrados; Manejo de recursos naturales; Ecología urbana; Cuencas hidrográficas
INTRODUCTION
The scientific community and authorities increasingly recognize that environmental research and management must seek to include both human and natural dimensions and support both ecological and social well-being (e.g., the Inter-Governmental Platform for Biodiversity and Ecosystem Services-IPBES) (Díaz et al. 2015). However, insufficient attention has been paid to the study of the ecological underpinnings of such concepts as ecosystem services (ES) that have been proposed to bridge these divides. Nonetheless, basic ecological information is a necessary first step to: a) properly implement and use ES in decision-making, b) conduct rigorous assessments of their quality and resilience, or c) predict how they may respond in the face of different management strategies or environmental change scenarios (Balnavera et al. 2012). Furthermore, most ecological studies of ES have occurred at regional and global scales, leaving out local ecosystem approaches (O’Farrell et al. 2012).
In short, we must understand not only what society expects the ecosystem to provide, but also have an idea of what it is capable of delivering now and into the future. For example, the MA (2006) found that 60% of the world’s ES are degrading. Particularly, urbanizing landscapes, where the majority of the world’s human population now resides, were found to have impaired capacity to provide for human well-being. Recently, ecological research has improved our understanding of human-nature relationship, overcoming previous biases towards studies of “natural” systems (Anderson et al. 2015). In particular, studies on urban stream ecosystem structure and function provide background knowledge to empirically link watershed ecosystem processes with the ES they provide (Alberti et al. 2007). For example, impervious land cover increases associated with urbanization can have a multitude of effects on stream macroinvertebrate communities via modifications of watershed hydrological and geomorphological processes (Wenger et al. 2009).
Macroinvertebrate communities and their responses to anthropogenic activity also are well documented in streams, and the multiple metrics developed to assess changes in community structure and function could be modified or applied as proxies to help detect perturbations to ES quality caused by urban activities (Purcell et al. 2009). Furthermore, macroinvertebrate taxa richness can be useful beyond assessing a watershed’s ability to support general ES quality, or “stream health,” and indeed may also signal shifts in specific ES. For instance, changes in the assemblage of functional feeding group (FFG), a classification system based on macroinvertebrate food habits (Cummins 1973), provide insights into the availability of different resource types and reflect the ability of a watershed to regulate levels of particulates in the system (Covich 1999).
In this study, we set out to determine the overall biophysical condition of urban watersheds in southern Patagonia’s Tierra del Fuego Archipelago and tested the local relevance of four stream biotic indices, which may be useful as ecosystem quality assessment tools for natural resource managers. Stream physical, chemical and biotic variables were compared between urban and reference sites to analyze how these variables changed as a function of increasing urbanization in four watersheds that drain the area around Ushuaia, Argentina. The region’s overall low human population density classifies it as a “wilderness area” (Mittermeier et al. 2003), but 98% of Argentine Tierra del Fuego’s population resides in just 3 cities (INDEC 2010), a pattern which is observed throughout southern Patagonia (e.g., 93% of the Magellan & Chilean Antarctic Region) (INE 2010). However, to date, little research has incorporated the impacts of urbanization as a driver of ecosystem change in this landscape. Therefore, baseline physical, chemical and biological data is needed for basic ecological understanding of Tierra del Fuego, as well as applied needs, such as assessing the future ability of these watersheds to provide for ES and developing criteria for monitoring programs that can provide guidance into how different modes of development are impacting them now and into the future.
MATERIALS AND METHODS
Study area
This study was conducted in the mainstems of four streams/rivers (Grande, Buena Esperanza, Pipo and Olivia), which are the principle watersheds that encompass the city of Ushuaia, Argentina (54.5° S, 68.2° W). The city has experienced rapid population growth in the last 10 years (~65000 inhabitants) (INDEC 2010). These mountain/valley catchments comprise a total area of 50931 ha and have annual flow rates ranging from 0.37 m3/s to 5.04 m3/s into the Beagle Channel (Table 1). Vegetation is mixed evergreen-deciduous Nothofagus forests, peat bogs and high Andean vegetation communities. The lower elevations receive approximately 520 mm of precipitation per year in the form of rain and snow (Iturraspe and Uriciuolo 2000). In addition to the effects of urbanization, all watersheds in Tierra del Fuego experience the impacts of invasive North American beaver (Castor canadensis) (Anderson et al. 2009).
Table 1. Principle watersheds of Ushuaia, Tierra del Fuego (Argentina) with total area, mean annual flow, river length, % of length in urban zone and % upstream urbanization at study reaches 1 and 2.
Tabla 1. Cuencas principales de Ushuaia, Tierra del Fuego (Argentina). Área total, caudal anual promedio, largo del río, % del largo en la zona urbana y % urbanización río arriba de los tramos de estudio 1 y 2.

In each stream, we studied three 100 m reaches during the austral spring (October, November) of 2011. This period was selected to be after the spring’s initial snowmelt freshets; no major rain events were recorded during this period, and flows were relatively stable. Furthermore, the mid-spring timing allowed sampling before major adult insect emergence began, which reduces the abundance and size classes of individuals remaining in streams. Urban reaches 1 and 2 (UR1 and UR2) were within the city limits, while the reference reach (RR) was found outside the urban limits. To exclude marine influence, UR1 was established approximately 150 m upstream from the river’s mouth. UR2 was located approximately midway between the points where the streams entered the urban boundary and their mouths. RRs were at least 200 m above the urban boundary and outside any apparent anthropogenic impacts. No sites were located in the immediate vicinity of beaver dams. Points where streams entered the urban boundary were determined with GIS, using shape files provided by the province’s Ministry of Social Development and overlaid onto spatial information from Google Earth digital maps.
Biotic and habitat variables
Each study reach was sub-divided for sampling into three transects at the nearest riffle/run habitat closest to the 0, 50, and 100 m point. At each reach, a rapid visual habitat assessment (RVAP) for high gradient streams was conducted, based on Barbour et al. (1999). The RVAP directs the observer to evaluate habitat conditions that are important to in-stream biota. Ten parameters were assessed with a numerical score from 0 (poor) to 20 (optimal). Parameters included in-stream substrate and morphological conditions and riparian features. Values were summed to give a final habitat score for the reach with the following scale: poor=0-50, marginal=60-100, sub-optimal=110-150, and optimal=160-200. Physico-chemical habitat variables were measured at transects one (downstream) and three (upstream) in each reach, including temperature (°C), conductivity (µS/cm), dissolved oxygen (%DO and mg/L DO), and pH. All measures were taken with a YSI 556 and water samples were collected with 20 ml plastic vials and analyzed in the lab for turbidity levels with an Oakton T-1 turbidity meter.
The macroinvertebrate assemblage was assessed at each transect, using a Surber net (0.1 m2), for a total of three collections per reach. Benthic material, then, was transported in 70% ethanol to the laboratory, where macroinvertebrates were separated from detritus and identified to the lowest possible taxonomic level (usually genus), using Merritt and Cummins (1996) and Fernández and Dominguez (2001). Each taxon was further classified per its purported FFG according to Miserendino and Pizzolón (2000) and Anderson and Rosemond (2007). We calculated: a) taxa richness and Shannon-Wiener diversity, b) taxa richness of Ephemeroptera, Plecoptera and Trichoptera (EPT) (Barbour et al. 1999), c) three family biotic indices (rapid bioassessment protocol [RBP] (Barbour et al. 1999), biotic monitoring Patagonian streams (BMPS) (Miserendino & Pizzolón 1999) and family biotic index (FBI) (Figueroa et al. 2003), and d) FFG assemblage. Then, three separate sensitivity values were assigned to each taxon based on criteria from three previously established biotic indices (RBP, BMPS, FBI). The scale for the BMPS (based on level of tolerance) was entered inversely to be directly comparable to the other two indices (based on level of sensitivity). The final index for the RBP and BMPS is based on the sum of tolerance scores for all the taxa found in the sample, while the FBI also incorporates taxa abundance:
where
n=abundance of taxa i
t=tolerance of taxa i
N=total number of individuals in the sample
Data analyses
The three sub-samples were averaged into a mean value per reach. Habitat and biotic variables were then compared between UR1 (most urban) and RR (reference) for each stream (n=4) using paired t-tests. We converted pH (a logarithmic scale) to its hydrogen concentration [H+] for statistical analysis. Reported values are the median on the pH scale. Stream macroinvertebrate community structure and composition were evaluated as invertebrate density (number of individuals/m2), taxonomic richness (s=number of taxa) and Shannon-Weiner diversity (H’). Effects on benthic community function were assessed with relative abundance (%) of all FFGs. To determine which family-level biotic indices most closely reflected empirical measurements of taxonomic richness, regression analysis was conducted for each biotic indicator versus s.
To understand how habitat and biotic variables changed with increasing urbanization, percent urbanization was calculated for each of the urban reaches (UR1 and UR2) for each stream by determining the length of the mainstem that was found within the urban limits from the mouth of the stream to the study reach, using Google Earth®. This length was then divided by the total length of the mainstem from mouth to headwaters, to arrive at a level of urbanization for each reach sampled. Stream macroinvertebrate community structure and composition were calculated as with the urban/reference comparison described above. We then carried out linear regressions for all habitat and biotic variables and FFG with respect to percent urbanization. All statistical analyses were conducted in JMP 9.0 (SAS 2009).
RESULTS
These urban streams were cool, clear and highly oxygenated. Using paired t-tests, no significant differences were detected in any of the physical habitat variables, comparing the most urban reaches (UR1) with the reference sites (RR) (P>0.05) (Table 2). However, we did observe an insignificant trend whereby turbidity and conductivity increased and dissolved oxygen (DO) decreased in urban sites, compared to reference conditions (see below for regression analysis). RVAP scores reflected a significant deterioration in habitat conditions (Table 2). Urbanization reduced the high “optimal” conditions at RR sites to values on the low end of the “sub-optimal” category at U1 sites.
Table 2. Mean (±S.E.) physico-chemical habitat and biotic variables and functional feeding group (FFG) composition for Reference and Urban reaches of southern Patagonian streams in Tierra del Fuego. Significant differences between sites (paired t-tests, P<0.05) are indicated with a @ sign.
Tabla 2. Promedio (±E.E.) de variables físico-químicas, bióticas y composición de grupos funcionales alimentarios (FFG) para tramos de Referencia y Urbanos en cuencas de la Patagonia Austral en Tierra del Fuego. Las diferencias significativas entre sitios (pruebas de t apareadas, P<0.05) se indican con un signo @.

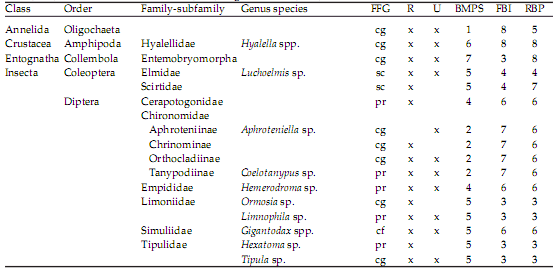
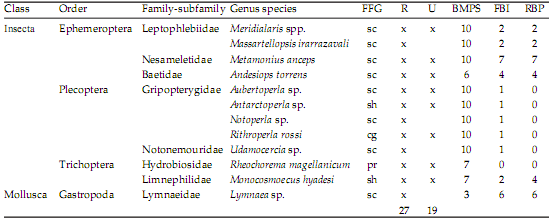
We recorded 28 taxa in these urban streams, including insects, crustaceans, annelids and mollusks (Table 3). Insects had the most identifiable taxa (n=25 of a total of 28) belonging to the orders of the Coleoptera, Diptera, Ephemeroptera, Plecoptera and Trichoptera. Taxonomic richness (P=0.01) and H’ (P=0.01) were significantly lower in UR1 than RR sites, but density (P=0.23) did not show a significant difference between sites (Table 2). FFG composition likewise did not show significant alterations between urban and reference reaches (Table 2). In both RR and UR1 sites, collector-gatherers (CGs) were dominant (55.0±6.2% and 75.0±13.0%, respectively), followed by scrapers (Sc) (25.0±10.4% and 24.0±13.4%, respectively). As with the habitat variables described above, these compositional differences were not significantly different between sites, but showed an increasing trend in the predominance of CG in urbanized sites. Three biotic indices (EPT, BMPS and RBP) showed a significant decrease in scores in UR1 sites compared to RR sites (P<0.02, 0.01, 0.03), but the FBI did not (Table 2). Lastly, the BMPS family biotic index was the best correlate to taxonomic richness, compared to the other indices (r2=0.97, P<0.001).
Table 3. List of all taxa described during the course of the study in reference (R) and urban (U) sites. FFG=functional feeding group; cg=collector-gatherer, pr=predator, cf=collector-filterer, sc=scraper. Tolerance scores are given for each biotic index, using family-level values obtained from Barbour et al. (1999), Miserendino and Pizzolón (1999), Figueroa et al. (2003).
Tabla 3. Lista de todos los taxa colectados durante el presente estudio en sitios de referencia (R) y urbanos (U). FFG=grupos funcionales alimentarios; cg=colector-recolector, pr=predador, cf=colector-filtrador, sc=raspador. Puntuación de tolerancia están dados por cada índice biótico, usando valores al nivel de familia obtenidos de Barbour et al. (1999), Miserendino and Pizzolón (1999), Figueroa et al. (2003).
The percent of upstream watershed urbanization of each urban reach ranged from 3.4% to 67.7% (Table 1), and increasing urbanization was significantly related to decreasing DO (r2=0.70, P=0.0006), taxonomic richness (r2=0.46, P=0.01) and EPT richness (r2=0.51, P=0.009). Decreases in H’ were only marginally related to increases in urbanization (r2=0.31, P=0.06), but on the other hand, turbidity (r2=0.67, P=0.001), conductivity (r2=0.89, P<0.0001), and relative abundance of CGs (r2=0.36, P=0.04) (Table 4) significantly increased as a function of increasing urbanization.
Table 4. Linear regressions of habitat and benthic macroinvertebrate community variables and benthic macroinvertebrate functional feeding groups (FFG), as a function of urbanization (U, expressed as a ratio of urban to non-urban) of southern Patagonian streams in Tierra del Fuego. Shown is the equation of each model, r2 and P values, denoting significant relationships with a @ sign (P<0.05).
Tabla 4. Regresiones lineales de variables ambientales, de la comunidad de macroinvertebrados bentónicos y de grupos funcionales alimentarios (FFG), como una función de la urbanización (U, expresada como una razón entre urbano y no-urbano) de las cuencas de la Patagonia Austral en Tierra del Fuego. Se muestra cada ecuación del modelo, r2 y valores P; se designan relaciones significativas con un signo @ (P<0.05).

DISCUSSION
Enhancing our understanding of southern Patagonian stream ecology
To date, stream ecology research in southern Patagonia has been biased towards watersheds with little or no impact from cities or towns (Moorman et al. 2006; Moorman et al. 2009; Anderson and Rosemond 2007, 2010; Contador et al. 2015). Furthermore, this region is often highlighted for its “pristine” and “wilderness” status (Mittermeier et al. 2003; Rozzi et al. 2012), in spite of the ample evidence that this landscape and its streams face a host of environmental pressures, such as silviculture and invasive species (Anderson et al. 2009; Moorman et al. 2009; Simanonok et al. 2011). While urbanized streams in the Tierra del Fuego Archipelago are few in number, when one considers the entire landscape, those that exist are densely populated. Plus, when we consider the ES provided by these watersheds, they constitute a very important subset, since the majority of the human population interacts with nature in these areas (Zagarola et al. 2014). Urban stream ecology, therefore, should be incorporated into the research agenda for southern South America, and in this line, we have sought to address some initial questions that are not only relevant here, but world-wide (Wenger et al. 2009).
Overall, our findings coincided with expectations for the effects of urbanization on stream biodiversity, but the nuance of these results indicated that the effect in Tierra del Fuego was more to the benthic community’s structure rather than its functional attributes. Indeed, diversity and richness indices were lower in urban sites, but the overall distribution of FFGs was less affected. This fact coincides with previous work on Fueguian streams that suggested they were less susceptible to food web changes caused by beaver invasion since they are naturally dominated (>50% of the biomass) by amphipod collector-gatherers (Hyalella spp.), which were also enhanced by invasive beaver engineering (see Anderson& Rosemond 2007, 2010). This aspect of the natural history of Fueguian streams also could be affecting our attempts to quantify stream quality, as measured by certain biotic indices. For example, the FBI did not show significant differences between urban and reference sites. We suggest that this outcome was due to the fact that the index uses taxa abundance as well as presence absence, and in so doing, the weight of amphipods, which are classified as a “tolerant” species, drive the biotic index regardless of stream condition. Plus, it bears mentioning that while the RVAP indicated our stream habitats to be degraded, compared to reference conditions, they were still in the third of four categories (i.e. sub-optimal), but nonetheless their benthic biotic indices were 2-3x lower than reference indices. In a similar study in northern Patagonia, Miserendino et al. (2008) found significant decreases in benthic biotic indices, but not by such a large magnitude. This could indicate that there are other disturbance factors not assessed here (e.g., chemical pollution, sewage effluent) that was not taken into account by our study of habitat quality. Overall, though, to define the best monitoring protocols for this region, we suggest that the indices that are based on taxa presence are best for these streams.
Regarding the broader implementation of these results, we first recognize that our urban sites were downstream of reference reaches, but we argue that the patterns observed here are not likely to be merely the result of natural gradients caused by changing habitat conditions, such as substrate or temperature. Indeed, the distances between our reference sites and urban sites were relatively short, ranging from 1.9 to 5.5 km, and with the exception of Buena Esperanza Stream did not involve large variations in elevation. Plus, there is no reason to expect taxonomic diversity to decrease or the number of impact tolerant species to increase as a function of the natural stream gradient. In particular, high latitude streams in the southern hemisphere, in places like New Zealand (Winterbourn and Cowie 1981) and the Tierra del Fuego Archipelago (Contador et al. 2015), have been shown to not follow the gradient expectations from postulates like the River Continuum Concept (Vannotte et al. 1980), an idealized model developed for eastern North American streams. It is, therefore, more probable that these differences are due to anthropogenic activity associated with urbanization, such as increased overland run-off caused by impervious cover and the disturbance of riparian areas (Wood and Armitage 1997).
Linking stream ecology to watershed ecosystem services
Enhancing our understanding of the link between ecosystem function and ES is crucial to ultimately determine relationships to human well-being (Díaz et al. 2015). Therefore, it is necessary to develop appropriate indicators of ES quality and understanding their responses to ecosystem change. These data allow us to begin to relate urbanization’s impacts to specific ES. For example, the reduction in benthic diversity in urban sites usually occurs in part due to micro-habitat homogenization, which is related to changes in water flow regulation (i.e., alteration of the flood regime) and increases in erosion control that together cause sedimentation of the interstitial spaces of substrate (Wood and Armitage 1997). Similarly, water regulation and erosion control services also influence levels turbidity and conductivity, and the deterioration of these regulation services by urbanization results in more particulates in the stream. Plus, Zagarola et al. (2014) demonstrated that urban residents in southern Patagonia highly value the cultural services provided by watersheds. Therefore, changes in the biophysical conditions of streams as a result of urbanization can both compromise the cultural services (i.e. recreation, sense of place, etc.) themselves or the ability of stakeholders to access them (e.g., channelization or paving limiting access to streams).
In this context, bioindicators have been shown to be more advantageous over exclusive assessments of physico-chemical measurements, since biotic communities integrate longer-term perturbations that might occur in a watershed (Karr and Chu 1999). Here, we found that the impact of urbanization on habitat, chemical and taxonomic variables was paralleled by a decrease in bioindicators of quality, especially the BMPS index. In a comparable study conducted in northern Patagonia, Miserendino et al. (2008) also found that species richness, EPT richness, Shannon-Weiner diversity and the BMPS index were lower at urban sites than reference reaches, and the percentage of CGs increased as a function of urbanization. However, the fact that their sites had approximately 2x higher values (BMPS of ~50 in TDF versus approximately ~111 in northern Patagonia) demonstrates the need to adapt expectations to local conditions. Plus, the fact that Fueguian streams are naturally dominated by collector-gatherers limits our ability to use FFGs as proxies for changes in ecosystem function or specific ES. Taking into account these caveats, we would recommend that the BMPS can be added to the suite of useful indicators for water quality assessment and also be assessed for freshwater provisioning and purification services. Such macroinvertebrate indicators in this biome also are more appropriate than fish, which are very species poor (Moorman et al. 2009) and algae or chemical conditions, which require more specialized equipment and professional expertise to conduct assessments.
CONCLUSIONS
Landscape planning in Tierra del Fuego should take into account urban drivers like impervious surface area, riparian deforestation and the presence of harmful effluent from residential and commercial sectors, which negatively impact local aquatic ecosystems and their biological communities, ultimately affecting the well-being of the human communities that depend on them. This is especially important when considering the high rate of population growth that is occurring in Ushuaia and the need to determine sustainable trajectories for the future expansion of the city and greater intensity of use within these watersheds. Therefore, these data can serve as an initial step in implementing biomonitoring protocols for Tierra del Fuego’s natural resource management agencies to assess the condition of its watersheds. Regionally-appropriate bioindicators, such as the BMPS, are not only ecologically meaningful, but they can also be integrated into citizen science approaches (Penrose and Call 1995) and thereby enhance dialogue between science and society, identified as a crucial area for urban stream ecology worldwide (Wenger et al. 2009). At the same time, new metrics also are needed to evaluate the condition of the cultural services provided by ecosystems, which we previously found are important to both the general community and natural resource scientists/managers (Zagarola et al. 2014; Martínez Pastur et al. 2016). These social dimensions are also an area which has been identified as lacking ES research (Balvanera et al. 2012). Enhancing the use of biophysical and cultural data in the management and communication of watersheds may help protect the diversity of these freshwater biological communities and preserve their ability to support local human community’s social well-being.
ACKNOWLEDGEMENTS. Financial support was provided by an NSF International Research Experience for Students grant (OISE 0854350) to CBA, a U.S. Student Fulbright Scholarship to JPAZ, and a CONICET Doctoral Scholarship to MEL. We thank Drs. M.V. Lencinas and A.E.J. Valenzuela for local laboratory and logistical support to JPAZ. Samples were collected under permit #09/11-Tierra del Fuego Science Commission. OSARA (www.osara.org) facilitated collaborations between Chile, Argentina and the U.S. JPAZ thanks R. Iturraspe, R. Reyes, G. Kreps, H. Ivancich, M. Santibañez, J. Bailey, L. Brandon, J. Fischer, L. Villanueva, C. Celis, J. Person and S. Keirns for their support.
REFERENCES
1. Alberti, M., D. Booth, K. Hill, C. Avolio, B. Coburn, S. Coe, and D. Spirandelli. 2007. The impact of urban patterns on aquatic ecosystems: An empirical analysis in Puget lowland sub-basins. Landscape and Urban Planning 80:345-361. [ Links ]
2. Anderson, C. B., and A. D. Rosemond. 2007. Ecosystem engineering by invasive exotic beavers reduces in-stream diversity and enhances ecosystem function in Cape Horn Chile. Oecologia 154: 141-153. [ Links ]
3. Anderson, C. B., and A. D. Rosemond. 2010. Beaver invasion alters terrestrial subsidies to subantarctic stream food webs. Hydrobiologia 652:349-361. [ Links ]
4. Anderson, C. B., G. Martínez Pastur, M. V. Lencinas, P. K. Wallem, M. C. Moorman, and A. D. Rosemond. 2009. Do introduced North American beavers engineer differently in southern South America? – An overview with implications for restoration. Mammal Review 39:33-52.
5. Anderson, C. B., J. C. Pizarro, R. Estévez, A. Sapoznikow, A. Pauchard, O. Barbosa, A. Moreira-Múñoz, and A. E. J. Valenzuela. 2015. ¿Estamos avanzando hacía una socio-ecología? Reflexiones sobre la integración de las dimensiones “humanas” en la ecología en el sur de América. Ecología Austral 25:263-272.
6. Balvanera, P., M. Uriarte, L. Almeida-Leñero, A. Altesor, F. DeClerck, T. Gardner, J. Hall, A. Lara, P. Laterra, M. Peña-Claros, D. M. Silva Matos, A. L. Vogl, L. P. Romero-Duque, L. F. Arreola, A. P. Caro-Borrero, F. Gallego, M. Jain, C. Little, R. de Oliviera-Xavier, J. M. Paruelo, J. E. Peinedo, L. Poorter, N. Ascarrunz, F. Correa, M. B. Cunha-Santino, A. P. Hernández-Sánchez, and M. Vallejos. 2012. Ecosystem services research in Latin America: the state of the art. Ecosystem Services 2:56-70. [ Links ]
7. Barbour, M. T., J. Gerritsen, B. D. Snyder, and J. B. Stribling. 1999. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, Second Edition. EPA 841-B-99-002. U.S. Environmental Protection Agency, Office of Water, Washington, D.C. [ Links ]
8. Barbour, M. T., and M. J. Paul. 2010. Adding value to water resource management through biological assessment of rivers. Hydrobiologia 651:17-24. [ Links ]
9. Chapin, F. S. I., E. S. Zavaleta, V. T. Eviner, R. L. Naylor, P. M. Vitousek, H. L. Reynolds, D. U. Hooper, S. Lavorel, O. Sala, S. E. Hobbie, M. C. Mack, and S. C. Díaz. 2000. Consequences of changing biodiversity. Nature 405:234-242. [ Links ]
10. Contador, T., J. Kennedy, J. Ojeda, and R. Rozzi. 2015. Sharp altitudinal gradients in Magellanic Sub-Antarctic streams: patterns along a fluvial system in the Cape Horn Biosphere Reserve (55° S). Polar Biology 38:1853-1866. [ Links ]
11. Covich, A. P., M. A. Palmer, and T. A. Crowl. 1999. The role of benthic macroinvertebrate species in freshwater ecosystems. BioScience 49:119-127. [ Links ]
12. Cummins, K.W. 1973. Trophic relations of aquatic insects. Annual Review of Entomology 18:183-206. [ Links ]
13. Cummins, K. W., R. W. Merritt, and P. C. N. Andrade. 2005. The use of invertebrate functional groups to characterize ecosystem attributes in selected streams and rivers in south Brazil. Studies in Neotropical Fauna and Environment 40:69-89. [ Links ]
14. Díaz, S., et al. 2015. The IPBES conceptual framework - connecting nature and people. Current Opinion in Environmental Sustainability 14:1-16. [ Links ]
15. Fernández, H. R., and E. Domínguez (eds.). 2001. Guía para la determinación de los artrópodos bentónicos sudamericanos. Fundación Miguel Lillo, San Miguel de Tucumán, Tucumán, Argentina. [ Links ]
16. Figueroa, R., C. Valdovinos, E. Araya, and O. Parra. 2003. Macroinvertebrados bentónicos como indicadores de calidad de agua de ríos del sur de Chile (Benthic macroinvertebrates as indicators of river water quality in southern Chile). Revista Chilena de Historia Natural 76:275-285. [ Links ]
17. Martínez Pastur, G., P. L. Peri, M. V. Lencinas, M. García Llorente, and B. Martín López. 2015. Spatial patterns of ecosystem services provision in southern Patagonia. Landscape Ecology 31:383-399. [ Links ]
18. INDEC (National Institute of Statistics and Census, Argentina). 2010. Censo Nacional de Población, Hogares y Viviendas. http://www.indec.mecon.gov.ar/. [ Links ]
19. INE (National Institute of Statistics, Chile). 2010. Censo Nacional de Población. http://www.ine.cl. [ Links ]
20. Iturraspe, R., A. Urciuolo, and J. Bathurst. 2009. Vulnerabilidad de las cuencas hídricas ante la recesión de los glaciares en Tierra del Fuego. XXII Congreso Nacional del Agua, Puerto Madryn, Argentina. [ Links ]
21. Iturraspe, R., and A. Urciuolo. 2000. Clasificación y caracterización de las cuencas hídricas de Tierra del Fuego. Actas del XVIII Congreso Nacional del Agua, Santiago del Estero, Argentina. [ Links ]
22. Karr, J. R., and E. W. Chu. 1999. Restoring Life in Running Waters: Better Biological Monitoring. Island Press, Washington D.C. [ Links ]
23. MA (Millennium Ecosystem Assessment). 2005. Ecosystems and Human Well-Being. Oct. 9 2010 www.maweb.org. [ Links ]
24. Martínez Pastur, G., P. L. Peri, M. V. Lencinas, M. García Llorente, and B. Martín López. 2016. Spatial patterns of cultural ecosystem services provision in southern Patagonia. Landscape Ecology 31:383-399. [ Links ]
25. Merritt, R. W., and K. W. Cummins (eds.). 1996. An Introduction to the Aquatic Insects of North America. Third edition. Kendall/Hunt Publishing Co., Dubuque, Iowa, USA. [ Links ]
26. Miserendino, M. L., and L. A. Pizzolón. 1999. Rapid assessment of river water quality using macroinvertebrates: a family level biotic index for the Patagonian Andean zone. Acta Limnologica Brasiliensa 11:137-148. [ Links ]
27. Miserendino, M. L., and L. A. Pizzolón. 2000. Macroinvertebrates of a fluvial system in Patagonia: altitudinal zonation and functional structure. Archiv fur Hydrobiolgie 150:55-83. [ Links ]
28. Miserendino, M. L., C. Brand, and C. Y. Di Prinzio. 2008. Assessing urban impacts on water quality, benthic communities and fish in streams of the Andes Mountains, Patagonia (Argentina). Water Air and Soil Pollution 194:91-110. [ Links ]
29. Mittermeier, R. A., C. G. Mittermeier, J. Brooks, J. Pilgrim, J. Konstant, G. A. B. da Fonseca, and C. Kormos. 2003. Wilderness and biodiversity conservation. Proceedings of the National Academies of Science 100:10309-10313. [ Links ]
30. Moorman, M. C., D. B. Eggleston, C. B. Anderson, A. Mansilla, and P. Szejner. 2009. The implications of North American beaver and trout invasion on native diadromous fish in the Cape Horn Biosphere Reserve, Chile. Transactions of the American Fisheries Society 138:306-313. [ Links ]
31. O’Farrell, P. J., P. M. L. Anderson, D. C. Le Maitre, and P. M. Holmes. 2012. Insights and opportunities offered by a rapid ecosystem service assessment in promoting a conservation agenda in an urban biodiversity hotspot. Ecology and Society 17:27. http://dx.doi.org/10.5751/ES-04886-170327.
32. Penrose, D., and S. M. Call. 1995. Volunteer monitoring of benthic macroinvertebrates – regulatory biologists perspectives. Journal of the North American Benthological Society 14:203-209.
33. Purcell, A. H., D. W. Bressler, J. M. Paul, M. T. Barbour, R. Rankin, and V. H. Resh. 2009. Assessment tools for urban catchments: developing stressor gradients. Journal of the American Water Resources Association 45:291-305. [ Links ]
34. Rozzi, R., J. Armesto, J. Gutiérrez, F. Massardo, G. E. Likens, C. B. Anderson, A. Poole, K. Moses, E. Hargrove, A. Mansilla, J. Kennedy, M. Willson, K. Jax, C. Jones, J. B. Callicott, and M. Arroyo. 2012. Integrating ecology and environmental ethics: earth stewardship in the southern end of the Americas. BioScience 62:226-236 [ Links ]
35. SAS Institute Inc. 2009. Using JMP 9. [ Links ]
36. Simanonok, M. P., C. B. Anderson, G. M. Pastur, M. V. Lencinas, and J. H. Kennedy. 2011. A comparison of impacts from silviculture practices and North American beaver invasion on stream benthic macroinvertebrate community structure and function in Nothofagus forests of Tierra del Fuego. Forest Ecology and Management 262:263-269. [ Links ]
37. Vannote, R. R., G. W. Minshall, K. W. Cummins, J. R. Sedell, and C. E. Cushing. 1980. The river continuum concept. Canadian Journal of Fisheries and Aquatic Sciences 37:130-137. [ Links ]
38. Wenger, S. J., A. H. Roy, C. R. Jackson, E. S. Bernhardt, T. L. Carter, S. Filoso, C. A. Gibson, N. B. Grimm, W. C. Hession, S. S. Kaushal, E. Martí, J. L. Meyer, M. A. Palmer, M. J. Paul, A. H. Purcell, A. Ramírez, A. D. Rosemond, K. A. Schofield, T. R. Schueler, E. Sudduth, and C. J. Walsh. 2009. Twenty-six key research questions in urban stream ecology: an assessment of the state of the science. Journal of the North American Benthological Society 28:1080-1098. [ Links ]
39. Winterbourn, M. J., J. S. Rounick, and B. Cowie. 1981. Are New Zealand streams really different? New Zealand Journal of Marine and Freshwater Research 15:321-328. [ Links ]
40. Wood, P. J., and P. D. Armitage. 1997. Biological effects of fine sediment in the lotic environment. Environmental Management 21:203-217. [ Links ]
41. Zagarola, J. P., C. B. Anderson, and J. Veteto. 2014. Perceiving Patagonia - An assessment of social values and perspectives regarding watershed ecosystem services and management in southern South America. Environmental Management 53:769-782. [ Links ]

