










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkEcología austral
versión On-line ISSN 1667-782X
Ecol. austral vol.28 no.2 Córdoba ago. 2018
ARTÍCULO
Growth of the wild potato Solanum commersonii (Solanaceae) among grass tussocks under different defoliation intensities
Osvaldo R. Vignolio*; Verónica N. Ispizúa; Eugenia Garavano & Natalia L. Murillo
Facultad de Ciencias Agrarias, Universidad Nacional de Mar del Plata-Estación Experimental Agropecuaria, Balcarce, Instituto Nacional de Tecnología Agropecuaria, Balcarce, Argentina.
*vignolio.osvaldo@inta.gob.ar; ovignoli@mdp.edu.ar
ABSTRACT
Solanum commersonii is a wild potato, valuable as a socioeconomic resource for a breeding program of feeding crops. In grasslands, this species grows associated to Paspalum quadrifarium, a forage grass managed under grazing conditions. We are interested in knowing in S. commersonii plants a) if they are benefitted growing together with P. quadrifarium plants, b) the effects of defoliation on growth, reproduction and survival, and c) if the vegetative and reproductive a��ributes of the plants differ with sampling site. Potato seeds were collected in two sampling sites of the Paititi Natural Reserve (Buenos Aires, Argentina), the lowest and highest. One potato plant per pot was cultivated. Under greenhouse conditions, it was analyzed the effect of two defoliation intensities, low and high, 50 and 75% plant height reduction compared to pre-defoliation, respectively. Under field conditions, S. commersonii plants were placed among P. quadrifarium tussocks. Control plants were placed outside of the tussocks, on short grass. Defoliation did not cause plant mortality and these regrew through sprouting from stolons. Stems, leaves and total biomass were significantly affected by defoliation intensity. Defoliation intensity also affected green leaves area and height per plant. Plants of the highest site developed more stems and tubers number and green leaves area than from the lowest site. Paspalum quadrifarium architecture generated thermal conditions that were more benign for S. commesonii plant growth. During winter, night temperature was lower (freezing) in control conditions than among tussocks. Shoot dead biomass of the control potato plants was 67% and 13% in the plants growing among P. quadrifarium tussocks. One hundred percent of control plants developed tubers, meanwhile those growing among tussocks only developed the 55%. Berries were only produced in control plants, 1.56 and 0.56 g/plant, in the highest and lowest sites, respectively.
Keywords: Natural reserve; Cutting; Microenvironment; Facilitation; Grass.
RESUMEN
Crecimiento de la papa silvestre Solanum commersonii (Solanaceae) entre matas de gramíneas bajo diferentes intensidades de defoliación.
Solanum commersonii es una papa silvestre de notable valor en los programas de mejoramiento de los cultivares de consumo. En los pastizales, esta especie crece asociada a los pajonales de Paspalum quadrifarium, una gramínea manejada con pastoreo. Estamos interesados en saber a) si las plantas de S. commersonii son beneficiadas cuando crecen con las de P. quadrifarium, b) qué efectos produce la defoliación sobre el crecimiento, la reproducción y la supervivencia de las plantas y c) si los atributos vegetativos y reproductivos difieren según los sitios de muestreos. Las semillas de papa fueron colectadas en dos sitios de la Reserva Natural Paititi (Buenos Aires, Argentina), el más bajo y el más alto. Las plantas, una por maceta, fueron cultivadas en condiciones de invernáculo. Se analizaron los efectos de la defoliación, baja y alta intensidad, 50 y 75%, como reducción de la altura de las plantas antes del corte. En condiciones de campo las plantas de S. commersonii fueron cultivadas entre las plantas de un pajonal de P. quadrifarium. Las plantas control crecieron fuera del pajonal, sobre pastos cortos. La defoliación no causó mortalidad de plantas y éstas rebrotaron desde los estolones. La intensidad de la defoliación afectó la biomasa total, de tallos y de hojas. También afectó el área foliar verde y la altura de las plantas. Las plantas de la Cima desarrollaron más área foliar, mayor número de tallos y tubérculos que las plantas de la Colmena. La arquitectura de las plantas de P. quadrifarium generó condiciones ambientales benignas para el crecimiento de S. commersonii, Durante el invierno, la temperatura nocturna fue más baja (heladas) en las condiciones control que en el pajonal. La biomasa aérea muerta de las plantas de papas control fue 67%; en cambio, en las que crecieron en el pajonal fue 13%. El 100% de las plantas control y el 55% de las que crecieron en el pajonal desarrollaron tubérculos. Las bayas sólo fueron producidas en las plantas control, 1.56 y 0.56 g/planta en el sitio más alto y en el más bajo, respectivamente.
Palabras clave: Reserva natural; Corte; Microambiente; Facilitación; Pastizal.
Recibido: 4 de febrero de 2017
Aceptado: 26 de febrero de 2018
Editor asociado: Esteban Jobbágy
INTRODUCTION
Solanum commersonii Dunal is a wild potato species, native of South America, valuable as a source of desirable traits in the breeding program of cultivated potato (Heřmanova et al. 2007; González et al. 2013). This species is perennial, herbaceous, diploid, with sexual and asexual spread through seeds, stolons and tubers. Flowering begins in spring and continues until middle autumn (Garavano et al. 2015). In grasslands of Buenos Aires Province (Argentina), S. commersonii grows associated to Paspalum quadrifarium Lam. tussocks (Prieto et al. 2016). Paspalum quadrifarium, a tall C4 grass, is a forage resource, warm-season perennial, native from South America. Its tussocks can reach until 2 m in height (Vignolio, personal observation). Green biomass of P. quadrifarium is consumed by cattle. As potato plants growing on cattle dung were found, we believe that S. commersonii berries and shoot biomass are consumed by the animals. If P. quadrifarium accumulates senescent biomass, it is partially or totally rejected by the cattle. In this case, it is advisable to burn in winter-spring and later defoliation, through grazing, to improve their quality and allow the establishment of other plant species from soil seed bank (Laterra 1997). We are interested in knowing if S. commersonii plants are benefitted growing together with P. quadrifarium plants, and to know the effects of defoliation on potato plant growth, reproduction and survival.
A large number of studies have reported that when plants species grow together, one of them can provide benefits to the other species through amelioration of harsh environmental conditions as wind, freezing temperatures, water stress and defoliation (Callaway 1995; Caviares et al. 2006; Bronstein 2009; Dekker 2011). Plants highly palatable can be ungrazed, avoiding local extinction, if they are growing with other less palatable plants species (Callaway 1995; Mingo and Oesterheld 2009). Vegetative and reproductive responses of S. commersonii plants to defoliation were not explored. Yield loss in Solanum tuberosum L. was correlated with the amount of tissue leaf removed (Ziems et al. 2006). Potato yield losses were 15 and 22% when leaf removal of the crops was 50 and 75%, respectively. High defoliation reduced number and weight of large tubers, increasing the number of small ones (Ziems et al. 2006). Tuber size affects sprout and plant performance. Plants from larger potato tubers exhibited greater physiological growth and higher yield than smaller ones (Masarirambi et al. 2012).
We worked in a Natural Reserve called Paititi (Mar del Plata, Buenos Aires. Hereafter, Reserve), a mountain rocky of Tandilia System (Echeverría et al. 2017). In this ecosystem, we found that S. commersonii plants have a wide ecological distribution growing among P. quadrifarium tussocks (Garavano et al. 2015). Differences in plant morphology were reported by Garavano et al. (2015), who analyzed S. commersonii plants growing in situ along altitudinal gradient of Paititi Reserve. Potato plants have a postraterosette architecture when they grow in open areas and an erect architecture when grow among P. quadrifarium tussocks (Garavano et al. 2015; Prieto et al. 2016). On 7 January 2014 (summer), possibly due to a large amount of standing dead biomass and litter of P. quadrifarium, the Reserve grasslands burned, initiating the processes of secondary succession in the plant communities where S. commersonii was one of the pioneer species. During the succesional process, a continuous colonization and extinction of S. commersonii plants were reported until that the plant communities reached the climax stage, being P. quadrifarium the dominant species with a large amount of standing dead biomass (Garavano et al. 2015; Ispizúa et al. 2015).
Our future objective is to develop in situ conservation strategies of S. commersonii in the context of ecosystem conservation. In situ conservation of natural resources requires knowing biotic and abiotic factors that affect target species growth (Iriondo et al. 2008; Heywood 2015; Volis 2015). The aim of present work was to analyze in S. commersonii plants from two sites of the Reserve, the highest and lowest, a) if they are benefitted growing together with P. quadrifarium tussocks, b) the effects of defoliation on growth, reproduction and survival, and c) if the vegetative and reproductive attributes of the plants differ with sampling site.
MATERIALS AND METHODS
Two experiments were developed in the EEA INTA Balcarce - Facultad de Ciencias Agrarias (UNMdP), Buenos Aires, Argentina, 37°45´ S and 58°18´ W, 130 m above sea level. The first experiment was developed in greenhouse conditions to study the effects of defoliation on growth, reproduction and survival on S. commersonii plants. The second experiment was carried out in field conditions to study if the S. commersonii plants are benefitted growing together with P. quadrifarium tussocks.
Sampling sites of the potato seeds Paititi Natural Reserve is situated in a mountain rocky ecosystem of Tandilia System, 37°55' 42.7'' S - 57°49'04'' W, at 24.5 km of Mar del Plata city (Buenos Aires, Argentina). Climate is warm, being the average annual temperature 14 °C, maximum temperature of 32 °C and minimum around 0 °C (Echeverría et al. 2017). The annual accumulated precipitation is approximately 1000 mm. Plant communities of the grasslands of the Reserve in climax stage are dominated by P. quadrifarium (Garavano et al. 2015; Ispizúa et al. 2015).
Berries of S. commersonii with seeds were collected during late spring 2014 in two sampling sites, Colmena and Cima, the lowest and highest point of the Reserve, respectively. Table 1 shows the characterization of both sites, the analyses test on the upper soil 15 cm. Total dry biomass of grassland was the accumulated from autumn 2014 to autumn 2015 without grazing, slope field was measured with clinometer and soil depth with a steel skewer.
Table 1. Environmental characterization of sampling sites of Paititi Natural Reserve, Colmena and Cima, where Solanum commersonii seeds were collected.
Tabla 1. Caracterización ambiental de los sitios de colecta de semillas de Solanum commersonii de la Reserva Natural Paititi, Colmena y Cima.

Plant material
On middle January 2015, S. commersonii seeds from Colmena and Cima, were sown on two layers of filter paper (Whatman) in Petri dishes, wetted with distilled water and placed in a growth chamber at 20 °C and 8 h light. Two Petri dishes per site, with 100 seeds each one, were prepared and during three weeks the seedlings were extracted and individually cultivated outdoor in pots of 15 cm diameter x 17 cm height, with drainages holes, and placed into the trays of 25 cm long x 15 cm wide x 4 cm height. The substrate was Argiudol soil, sand and perlite (2:1:1) with pH (soil:H2O, 1:2.5)=8.1; available phosphorus (Bray 1 method)=61.6 ppm; organic matter content=2.70% and nitrate=52.4 ppm (Soil Survey Staff 1999). The plants were used for both experiments.
Experiment 1. Greenhouse conditions
On 6 April 2015, after 30 days of plant acclimation in greenhouse conditions, the effect of defoliation intensity on S. commersonii plants was analyzed. At the beginning of the experiment, potato dry biomass was determined through a destructive sampling of five plants per site. Dry weight was determined after drying at 60 °C until constant weight. Roots, stolons and shoot dry biomass were not significantly different between sites being in average (±SE) 0.94±0.01 (F=0.21; P=0.65), 1.13±0.18 (F=0.04; P=0.84) and 4.55±0.48 g/plant (F=0.21; P=0.65), respectively. Plant maximum height and stems number per plant were not significantly different between sites, 27.1±2.56 cm (F=0.78; P=0.4) and 4.1±0.64 N/ plant (F=1.22; P=0.3), respectively. The plants have not developed tubers.
Fifteen plants per sampling sites were average selected and different defoliation intensities were applied. Low defoliation intensities (LDI), plant height reduced 50% compared to pre-defoliation and high (HDI), plant height reduced 75%. Control plants were not defoliated. Five plants by treatment were used. Cutting shoot dry weight was significantly different (F=11.76; P=0.002) between intensities, being 1.05±0.18 g/plant for LDI and 2.11±0.27 g/plant for HDI.
On 30 June 2015, a second defoliation was applied in all plants at soil level, included the Control treatment. Biomass was collected and dry weight was determined. Green leaf area was determined with a planimeter (Li-Cor LI 3000). During the experimental period, stems number and plants height were recorded every two weeks. Height determination was from soil level until the longest stem apex. The plants were disposed at random, periodically rotated and irrigated. Symptoms of water stress, pests and herbivores attacks were not observed. Air temperature, intercepted photosynthetically active radiation and relative humidity data were recorded daily during experimental period using meteorological station datalogger (Table 2).
Table 2. Monthly means of air temperature (Tm), relative humidity (RH) and solar radiation under greenhouse conditions. Experiment 1.
Tabla 2. Medias mensuales de la temperatura del aire (Tm), humedad relativa (RH) y radiación solar en condiciones de invernáculo. Experimento 1.
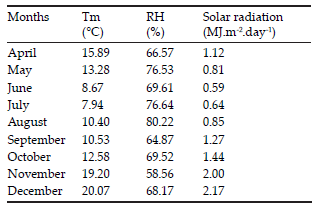
The experiment finished on 11 December 2015. Soil was sieved and tubers were collected, washed, dried at room temperature and weighed. Relationship between tubers number and stems per plant was performed. Shoot biomass senescence began in August 2015 and at the end of the experiments, aerial tissues were degraded, unquantifiable.
Experiment 2. Field conditions
On 7 April 2015, at the beginning of the Autumn, S. commersonii plants were cultivated in field conditions among P. quadrifarium tussocks. The tussocks had an average height of 105.75±3.79 cm and a standing crop of senescent biomass of 2383.16±522.21 g/m2 and green biomass of 17.33±5.84 g/m2 (0.72%). Pots with potato plants of Cima and Colmena were placed among P. quadrifarium tussocks (Figure 1A). Control S. commersonii plants were placed in holes, near to P. quadrifarium tussocks, at soil level in a cutting plants community composed by P. quadrifarium, Festuca arundinacea Schreb Lotus tenuis, Lolium multiflorum (L), Cynodon dactylon (L) Pers. (Figure 1B). Through a destructive sampling was determined that potato roots, stolons and total dry biomass were not significantly different between sites, being the average 0.924±0.025, (F=0.22; P=0.6), 1.10±0.04 g/plant (F=0.04; P=0.8) and 6.62±0.67 g/plant (F=0.04; P=0.84), respectively. Plant maximum height and stems number per plant were not significantly different between sites, 23.44±3.29 cm/plant (F=1.23; P=0.3) and 5.55±0.88 N/plant (F=3.88; P=0.08), respectively.

Figure 1. Solanum commersonii plants growing A) among Paspalum quadrifarium tussocks and B) in control conditions with frozen shoot biomass. Reference: 20 cm.
Figura 1. Plantas de Solanum commersonii creciendo A) entre las matas de Paspalum quadrifarium y B) en condiciones control con la biomasa aérea afectada por la helada. Referencia: 20 cm.
Photosynthetically active radiation (PAR) at the top of the S. commersonii plants was 22.50±7.45%, respect to the incident radiation at the Control conditions. PAR was determined with a radiometer (LI-Cor 188 B) connected to a line quantum sensor LI-Cor 191 SB. During the experimental period, PAR, temperature and relative humidity were recorded with a datalogger placed among the tussocks, together with S. commersonii plants. Other temperature sensor was located outside the tussocks, at 0.30 m above of the P. quadrifarium canopy. The temperature sensor was protected of heat from the solar radiation with a wellventilated pot of telgopor. Complementary meteorological data were provided by the EEA INTA Balcarce station (Table 3).
Table 3. Rainfall, daily evapotranspiration (ETP), solar radiation, relative humidity (RH) and monthly means of maximum (Max.) and minimum (Min.) air temperature during 2015 in control conditions. Data were provided by the EEA INTA Balcarce meteorological station. Experiment 2.
Tabla 3. Lluvias, evapotranspiración diaria (ETP), radiación solar, humedad relativa (RH) y media mensual de las temperaturas máximas (Max.) y mínimas (Min.) del aire durante 2015 en condiciones control. Los datos fueron proporcionados por la estación meteorológica de la EEA INTA Balcarce. Experimento 2.

Total number of plants used was 72 (18 plants x 2 sample sites x 2 treatments). On 30 June 2015, after 84 experimental days, a destructive sampling on 8 plants per site and treatment (32 plants) was performed. Total shoot biomass per plant (green plus senescent), roots, stolons, leaf green area and plant maximum height were determined. The plants have not developed tubers. The other 40 plants (10 plants x 2 sample sites x 2 treatments) were kept in the same field conditions until finishing the experiment, on 22 December 2015. At the end of the experiment, shoot biomass was degraded, unquantifiable. The soil was sieved and the tubers were collected and analyzed as in Experiment 1.
Statistical analysis
Data were analyzed using two-way analysis of variance (ANOVA). Data expressed as percentages were arcsine square root transformed before the analysis and data were back transformed for its presentation. When ANOVA was significant, the means were separated by Fisher´s LSD test at the 5% level of probability (Steel and Torrie 1980). All calculations were performed using Statistica 6.0 software. Tubers weight was transformed as ln(x+1) before the analysis and back transformed for presentation. On 30 June 2015, for both experiments, data of biomass per plant of stems, green leaf, roots and stolons were determined for the analysis. On the same date, the data of the plant height and green leaf area were also determined. Tuber, number and fresh weight were determined on 11 and 22 December 2015 by experiments 1 and 2, respectively. Relationships between stems number (determined on 13 October 2015) and tubers number per plant were determined using a lineal function.
RESULTS
Experiment 1
Defoliation did not cause plant mortality and these regrew through sprouting from stolons.
Defoliation intensity significantly affected the biomass per plant of stems (F=15.02; P=0.00004), green leaves (F=17.31; P=0.00001), standing dead biomass (F=6.89; P=0.0038) and total (F=26.58; P=0.00001) (Figure 2). Green leaves area (F=7.85; P=0.0205) and plants height (F=21.81; P=0.000002) were also significantly affected by defoliation intensities (Table 4). Cima plants developed more stems number (F=10.54; P=0.003), total biomass of tubers (F=4.45; P=0.049) and tubers number (F=4.72; P=0.0381) than those at Colmena site (Table 5). Weight per tuber and tuber number per plant, were not significantly affected by the treatments, 1.02±0.14 g and 13.63±1.88 N/ plant, respectively. Stems number per plant was positively related (R²=0.56; P=0.001) with tubers number (Figure 3).

Figure 2. Biomass of different vegetative variables (average±SE) of Solanum commersonii plants from two sampling sites of the Paititi Natural Reserve. References: Defoliation intensities, low (LDI) and higher (HDI). Variables with distinct letters are significantly different among defoliation intensities (P<0.05), experiment 1.
Figura 2. Biomasa vegetativa de diferentes variables (media±EE) de las plantas de Solanum commersonii procedentes de dos sitios de la Reserva Natural Paititi. Intensidad de defoliación: baja (LDI) y alta (HDI). Variables con distintas letras son significativamente diferentes entre intensidades de defoliación (P<0.05), experimento 1.
Table 4. Vegetative variables (average±SE) of Solanum commersonii from the Paititi Natural Reserve growing under different defoliation intensities. References: Defoliation intensities, low (LDI) and higher (HDI). Variables within each row with distinct letters are different among defoliation conditions (P<0.05). Experiment1.
Tabla 4. Variables vegetativas (media±EE) de Solanum commersonii de dos sitios de muestreo de la Reserva Natural Paititi creciendo bajo diferentes intensidades de defoliación. Referencias: Defoliación baja (LDI) y alta (HDI) . Variables dentro de cada fila con letras distintas son diferentes entre condiciones de defoliación (P<0.05). Experimento1.

Table 5. Vegetative variables (average±SE) of Solanum commersonii from two sampling sites, Cima and Colmena, of the Paititi Natural Reserve growing under different defoliation intensities. Variables within each row with distinct letters are different between sites (P<0.05). Experiment 1.
Tabla 5. Variables vegetativas (media±EE) de Solanum commersonii de dos sitios de muestreo de la Reserva Natural Paititi creciendo bajo diferentes intensidades de defoliación. Variables dentro de cada fila con letras distintas son diferentes entre sitios (P<0.05). Experimento 1.


Figure 3. Relationship between stems and tubers number of Solanum commersonii plants growing in greenhouse and different defoliation intensities. Cima control: ●; Cima 50% defoliation:■ ; Cima 75% defoliation:▲ ; Colmena control: ◦ ; Colmena 50% defoliation: □; Colmena 75% defoliation: Δ. Experiment 1.
Figura 3. Relación entre el número de tallos y de tubérculos de las plantas de Solanum commersonii creciendo en condiciones de invernadero y diferentes intensidades de defoliación. Cima control: ●; Cima 50% defoliación:■ ; Cima 75% defoliación: ▲; Colmena control: ◦ ; Colmena 50% defoliación:□; Colmena 75% defoliación: Δ. Experimento 1.
Experiment 2
The biomass of the stems (F=8.34; P=0.0071), roots (F=17.41; P=0.0002), stolons (F=40.24; P=0.00001), green leaf (F=18.94; P=0.00014) and total of tubers (F=16.33; P=0.0003) were significantly higher in Control than the plants growing among P. quadrifarium tussocks (Figure 4). Dead shoot biomass, due to damage by low temperature (freezing), was significantly higher in Control (F=134.66; P=0.00001) than in the plants growing among P. quadrifarium tussocks (Figure 4). Plant height was affected by the treatment (F=113.84; P=0.00001), sampling site (F=4.36; P=0.045) and interaction treatment x sampling site (F=4.54; P=0.041). Plants growing among tussocks were significantly taller (22.18±1.91 cm/plant) than in Control (3.53±0.28 cm/ plant) conditions. Cima plants growing among tussocks were significantly shorter (18.50±2.09 cm/plant) than in Colmena ones (25.87±2.73 cm/plant).

Figure 4. Biomass of vegetative variables (average±SE) of Solanum commersonii from two sampling sites of the Paititi Natural Reserva growing under two experimental conditions. Control (short grass) and tussocks, Paspalum quadrifarium plants. A same variable between experimental conditions is significantly different (P<0.05). Experiment 2.
Figura 4. Biomasa de variables vegetativas (media±EE) de las plantas de Solanum commerosnii procedentes de dos sitios de la Reserva Natural Paititi creciendo bajo dos condiciones experimentales. Control (pastos cortos) y matas, plantas de Paspalum quadrifarium. Una misma variable entre condiciones experimentales es significativamente diferente (P<0.05). Experimento 2.
Green leaf area was significantly higher in potato plants growing among tussocks (F=41.13; P=0.00001) than those at Control conditions (Table 6). The tubers were produced between the beginning of winter (30 June 2015) and the end of the experiment, at the beginning of summer (22 December 2015). Tubers were produced in 100% of Control plants and in 55% of the plants growing among tussocks. Plant growing in Control conditions produced more tubers number, being each one more weighed than in tussocks (Table 6). Berries were only produced in Control plants of Cima 1.56±1.01 g/plant and Colmena 0.56±0.24 g/plant.
Table 6. Vegetative variables (average±SE) of Solanum commersonii from two sampling sites of the Paititi Natural Reserva, Colmena y Cima, growing under two experimental conditions, control (short grass) and among Paspalum quadrifarium tussocks. Variables within each row with distinct letters are different among experimental conditions (P<0.05). Experiment 2.
Tabla 6. Variables vegetativas (media±EE) de Solanum commersonii de dos sitios de muestreo de la Reserva Natural Paititi, Cima y Colmena, creciendo bajo dos condiciones experimentales, control y entre las matas de Paspalum quadrifarium. Variables dentro de cada fila con letras distintas son diferentes entre condiciones experimentales (P<0.05). Experimento 2

Air temperature differed between experimental conditions. During the day, the air temperature was higher in Control conditions than among the tussocks. On the other hand, during the night the temperature was higher among tussocks than in Control conditions (Figure 5).

Figure 5. Detail of air temperature during nine experimental days (15 June 2015 - 24 June 2015) in control and among Paspalum quadrifarium tussocks. Arrows indicate air temperature at midday. Experiment 2.
Figura 5. Detalles de la temperatura del aire durante nueve días del experimento (15 de junio de 2015 - 24 junio de 2015) en el control y entre las matas de Paspalum quadrifarium. Las flechas indican la temperatura del aire al mediodía. Experimento 2.
DISCUSSION
Defoliation did not affect the plant survival and these regrew through sprouting from the stolons. Defoliation studies in potato plants have been conducted mainly in commercial cultivars as S. tuberosum, being scarce the reports in wild potato. Altesor et al. (2014, 2016) reported that S. commersonii plants, native of Uruguay, were defoliated by Tequus schrottkyi, a Hymenoptera whose larvae feed the leaves, but its effects on plant reproduction were not informed. Solanum commersonii plant response to defoliation was of partial compensation of shoot biomass respect to control conditions (Ferraro and Oesterheld 2002), and was attributed to the loss of photosynthetic tissue (green leaves area and stems). Tubers number and weight were not significantly affected, showing compensation in this organ.
Our results are coincident with the reported by Cranshaw and Radcliffe (1980) who reported that commercial yield potato was not affected when defoliation, early in the season (stage vegetative), was between 10 and 33% of shoot biomass. On the other hand, defoliation during the mid-season or full bloom resulted in tuber number and weight reduction (Shields and Wyman 1984; Ziems et al. 2006). In our experiment, defoliation in vegetative stage did not affect biomass per tuber and tubers number per plant, because cutting was applied before the beginning tubers formation (Cranshaw and Radcliffe 1980). However, tubers and stems number per plant differed with sampling site, being these attributes higher in Cima than in Colmena. The allometric relationships between the number of stems and tubers suggests that tubers production is influenced by aerial vegetative growth (Ziems et al. 2006; Shayanowako et al. 2015).
Although potato plants grew in pots among the P. quadrifarium tussocks, the results showed that grass canopy attenuates the effects of environmental stress (Callaway 1995; Bruno et al. 2003). Senescent shoot biomass of S. commersonii was higher in plants growing in Control than among the tussocks. Paspalum quadrifarium architecture provided shelter, reducing the effects of cold temperature during the winter nights, generating benign microenvironment for S. commersonii plants. This result is consistent with our observations in Paititi Reserve in winter season, where S. commersonii with shoot green biomass was showed growing among P. quadrifarium tussocks.
In the grasslands, P. quadrifarium plants retain their senescent leaves and this forage can be totally or partially rejected by the cattle. In this case, the standing dead foliage of P. quadrifarium could also exert on S. commersonii plants some effect of protecting against large herbivores (Callaway 1995; Mingo and Oesterheld 2009). However, S. commersonii plants growing among P. quadrifarium tussocks had a cost in terms of tubers and berries production, in other words, in vegetative and sexual reproduction. These results could be as consequence of the reduced photosyntetically active radiation among tussocks canopy.
Morphological attributes of S. commersonii plants as green leaves area, stems number and height, varied with the sampling sites, which can be attributed to phenotypic plasticity and/or genetic diversity (Gratani 2014). Cima plants growing among tussocks were shorter than Colmena one.
Morphological differences in S. commersonii plants growing along the altitudinal gradient of Natural Reserve Paititi were also reported by Garavano et al. (2015). Coinciding with our results, plant height was higher in Colmena (37.46 cm/plant) than in Cima (16.10 cm/plant) (Garavano et al. 2015). Solanum commersonii pollination is entomophily. Flowering time is asynchronous, differs in the time and in the space along of the altitudinal gradient studied (Graravano et al. 2015). Therefore, flowering time could restrict the genetic exchange among plants growing in different sites (Dekker 2011; Garavano et al. 2015).
Solanum commersonii defoliation did not affect plant survival and these regrew from stolons. Paspalum quadrifarium architecture provided shelter, benign microenvironment for S. commersonii growth during the winter. However, it represented a cost in tubers and berries production and securely in its sexual and asexual propagation. Some plant morphological attributes were different between sites, they can be by phenotypic plasticity and/or genetic diversity. Further studies along these investigation lines involving in S. commersonii molecular analysis will help to understand if the ecological distribution at the Reserve corresponds to different populations. This information is important to develop in situ conservation strategies of target plant species as S. commersonii in the context of ecosystem conservation.
ACKNOWLEDGEMENTS
We thank María Rosa Desirello and Sara Garfinkel for critical reading of the text, S. Muñoz by helped us in the field works and E. Antonio González Zugasti, owner of the Paititi Natural Reserve. This work was supported by the Facultad de Ciencias Agrarias, Universidad Nacional de Mar del Plata-Estación Experimental Agropecuaria Balcarce, Instituto Nacional de Tecnología Agropecuaria. Proyecto AGR 484/15. We also thank the referees of journal Ecología Austral for their valuable comments.
REFERENCES
1. Altesor, P., A. García, E. E. Font, A. Rodríguez-Haralambides, F. Vilaró, M. Oesterheld, R. Soler, and A. González. 2014. Glycoalkaloids of wild and cultivated Solanum: Effects on specialist and generalist insect herbivores. J Chem Ecol DOI 10.1007/s10886-014-0447-8. [ Links ]
2. Altesor, P., A. González, and S. Schmidt. 2016. First report of Tequus schrottkyi (Konow) (Hymenoptera: Pergidae) in Uruguay, and information about its host plant and biology. Biodiversity Data Journal 4:e7538. doi:10.3897/ BDJ.4.e7538. [ Links ]
3. Bronstein, J. L. 2009. The evolution of facilitation and mutualism. J Ecol 97:1160-1170. [ Links ]
4. Bruno, J. F., J. J. Stachowicz, and M. D. Bertness. 2003. Inclusion of facilitation into ecological theory. Trends Ecol Evol 18:119-125. [ Links ]
5. Callaway, R. M. 1995. Positive interactions among plants. Bot Rev 61:306-349. [ Links ]
6. Caviares, L. A., E. I. Badano, A. Sierra-Almeida, S. Gómez-González, and M. A. Molina-Montenegro. 2006. Positive interactions between alpine plant species and the nurse cushion plant Laretia acaulis do not increase with elevation in the Andes of central Chile. New Phytol 169:59-69. [ Links ]
7. Cranshaw, W. S., and E. B. Radcliffe. 1980. Effect of defoliation on yield of potatoes. J Econ Entomol 73:131-134. [ Links ]
8. Dekker, J. 2011. Evolutionary Ecology of Weeds. Copyright 1.1.11. URL: agronwww.agron.iastate.edu/~weeds/PDF_Library/Agron_517/EEW_ClassUse_1.1.11. [ Links ]
9. Echeverría M. L., S. I. Alonso, and V. M. Comparatore. 2017. Survey of the vascular plants of Sierra Chica, the untouched area of the Paititi Natural Reserve (southeastern Tandilia mountain range, Buenos Aires province, Argentina). Check List 13:1003-1036. [ Links ]
10. Ferraro, D. O., and M. Oesterheld. 2002. Effect of defoliation on grass growth. A quantitative review. Oikos 98:125- 133. [ Links ]
11. Garavano, M. E., V. N. Ispizúa, N. O. Maceira, and O. R. Vignolio. 2015. Estudio de Solanum commersonii Dunal creciendo entre matas de Paspalum quadrifarium Lam. Jornadas Argentinas de Botánica. 23 al 26 de septiembre 2015. [ Links ]
12. González, M., G. Galván, M. I. Siri, A. Borges, and F. Vilaró. 2013. Resistencia a la marchitez bacteriana de la papa en Solanum commersonii Dun. Agrociencia Uruguay. 7:45-54. [ Links ]
13. Gratani, L. 2014. Plant phenotypic plasticity in response to environmental factors. Review Article. Adv Bot DOI: dx.doi.org/10.1155/2014/208747 [ Links ]
14. Heřmanova, V., J. Jan Bárta, and V. Curn. 2007. Wild potato species: characterization and biological potential for potato breeding. Review. Czech J Genet Plant Breed 43:73-81. [ Links ]
15. Heywood, V. H. 2015. In situ conservation of plant species-an unattainable goal? Israel Journal of Plant Sciences 63(4): 211-231. DOI: dx.doi.org/10.1080/07929978.2015.1035605. [ Links ]
16. Iriondo, J. M., N. Maxted, and M. E. Dulloo. 2008. Conserving plant genetic diversity in protected areas. Population management of crop wild relatives. Eds. Irondo et al. CABI Head Office Nosworthy Way Wallingford Oxfordshire OX10 8 DE. UK. [ Links ]
17. Ispizúa, V., E. Garavano, O. R. Vignolio, and H. Angellini. 2015. Estudio de la vegetación en un ecosistema del sistema de Tandilia (Buenos Aires, Argentina). Cuarto Congreso Internacional de Servicios Ecosistémicos en los Neotrópicos: de la investigación a la acción. 30 de septiembre al 3 de octubre 2015 - Mar del Plata. Buenos Aires, Argentina. [ Links ]
18. Laterra, P. 1997. Post-burn recovery in the flooding Pampa: impact of an invasive legume. J Range Manage 50:274- 277. [ Links ]
19. Masarirambi, M. T., F. C. Mandisodza, A. B. Mashingaidze, and E. Bheghe. 2012. Influence of plant population and seed tuber size on growth and yield components of potato (Solanum tuberosum). Int J Agric Biol 14:545-549. [ Links ]
20. Mingo, A., and M. Oesterheld. 2009. Retention of dead leaves by grasses as a defense against herbivores. A test on the palatable grass Paspalum dilatatum. Oikos 118:753-757. [ Links ]
21. Prieto, A., V. N. Ispizúa, and A. M. Clausen. 2016. Distribución y variabilidad morfológica de poblaciones de Solanum commersonii (Solanaceae) en la región pampeana de la Argentina. Bol Soc Argent Bot 51:59-71. [ Links ]
22. Shayanowako, A., R. Mangani, T. Mtaita, and U. Mazarura. 2015. Influence of main stem density on irish potato growth and yield: A review. Ann Res Rev Biol 3:229-237. [ Links ]
23. Shields, E. J., and J. A. Wyman. 1984. Effect of defoliation at specific growth stages on potato yields. J Econ Entomol 77:1194-1199. [ Links ]
24. Soil Survey Staff – USDA. 1999. Soil Taxonomy: A Basic System for Classifying Soils. Agriculture Handbook 436, United States Government Printing Office, Washington D.C.
25. Steel, R., and H. T. Torrie. 1980. Principles and procedures of statistics. McGraw-Hill, New York, USA. [ Links ]
26. Volis, S. 2015. Species-targeted plant conservation: time for conceptual integration. Israel J Plant Sci DOI: dx.doi.org/ 10.1080/07929978.2015.1085203 [ Links ]
27. Ziems, J., B. J. Zechmann, W. Wyatt Hoback, F. C. Wallace, R. A. Madsen, T. E. Hunt, and L. G. Higley. 2006. Yield Response of Indeterminate Potato (Solanum tuberosum L.) to Simulated Insect Defoliation. Agron J 98:1435-1441. [ Links ]

