










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgriscientia
versión On-line ISSN 1668-298X
Agriscientia v.23 n.2 Córdoba jul./dic. 2006
Contribución relativa del nitrógeno del suelo y del fijado biológicamente a la economía de la nutrición nitrogenada de maní (Arachis hypogaea L.) en diferentes condiciones de fertilidad
Castro, S., G. Cerioni, O. Giayetto y A.Fabra
S. Castro y A.Fabra. Departamento Ciencias Naturales, Facultad de Ciencias Exactas, Físico-Químicas y Naturales. G. Cerioni y O. Giayetto. Departamento Producción Vegetal, Facultad de Agronomía y Veterinaria. Universidad Nacional de Río Cuarto. Ruta 36, Km. 601. 5800 Río Cuarto - Argentina. ogiayetto@ayv.unrc.edu.ar
Resumen
La producción de maní en Argentina se concentra en la región central de la provincia de Córdoba, la cual experimentó últimamente una pérdida importante de la productividad de los suelos y una declinación aleatoria del rendimiento de los cultivos. La contribución relativa de la fijación biológica (FBN) de nitrógeno al maní en suelos de diferente fertilidad no ha sido suficientemente estudiada. Entonces, se evaluó el efecto de cepas de rizobios (TTOO2R, SEMIA 6144R y TAL 1000R) sobre el rendimiento y el balance de nitrógeno de maní cultivado en suelos con alto y bajo contenido del nutriente. No hubo diferencias significativas en los parámetros simbióticos y de rendimiento del cultivo entre las cepas introducidas y las nativas, pero se observó una contribución relativa mayor de la FBN en el suelo con bajo contenido de nitrógeno (~58% de contribución) que en el suelo con alto contenido (~27% de contribución). Esta comprobación del aporte relativo de la FBN asociada a la fertilidad del suelo, no registra antecedentes en la región central de Córdoba y debería recibir mayor consideración en el manejo del cultivo particularmente por su localización actual al sur de la provincia, donde los suelos presentan menores niveles de fertilidad. El rendimiento de maní confitería mostró mayores valores, si bien no significativos, con la inoculación en los 3 años del estudio.
Palabras clave: Maní; Bradyrhizobium sp.; Fijación biológica del nitrógeno; Nitrógeno del suelo; Ocupación de nódulos.
Castro, S., G. Cerioni, O. Giayetto and A.Fabra, 2006. Relative contribution of biological fixed nitrogen and soil nitrogen to the nutrition economy of peanut (Arachis hypogaea L.) under different conditions of soil fertility. Agriscientia XXIII (2): 55-66
Summary
The peanut production in Argentina is concentrated in the central region of Córdoba province. At present, losses of soil fertility and a random decline peanut yield have been reported for this area. The relative contribution of biological nitrogen fixation (FBN) in peanut plants cropped in soils with different fertility, has not been extensively studied. An experiment was carried out to determine the effects of rhizobia strains (TTOO2R, SEMIA 6144R and TAL 1000R) on peanut crop yield and plant nitrogen balance under different conditions of soil nitrogen. The results did not show significant differences in the symbiotic parameters and peanut crop yield between native and non-native strains. However, a relative higher contribution of biological nitrogen fixation was observed in low nitrogen (~58% contribution) than in high nitrogen soil content (~27% contribution). The relative contribution of FBN associated with soil fertility has not been investigated in the central region of Córdoba and it becomes particularly important in crop management for the current Southern cropping region where soils have lower fertility levels. The peanut crop showed a confectionary higher yield, although no significant, during the three years of experiments.
Key words: Peanut; Bradyrhizobium sp.; Biological nitrogen fixation; Soil nitrogen; Nodule occupancy.
Fecha de recepción: 10/05/06; fecha de aceptación: 27/12/06
Introducción
Las bacterias capaces de formar nódulos en plantas leguminosas, colectivamente denominadas rizobios, son importantes habitantes del suelo. Tanto su número como su especificidad dependen de las condiciones bióticas y abióticas del ambiente edáfico y de las especies vegetales leguminosas, nativas o cultivadas, que crecen en el área (Simon et al., 1996). Además de la asimilación de nitrógeno atmosférico, a través de la simbiosis con bacterias formadoras de nódulos, las especies leguminosas utilizan para su nutrición mineral el nitrógeno proveniente del suelo o el aplicado como fertilizante. En consecuencia, son tres las fuentes de nitrógeno disponibles para el crecimiento de estas plantas y el llenado de sus semillas: nitrógeno mineral proveniente del suelo, nitrógeno atmosférico procedente de la fijación biológica y aquél movilizado desde órganos de acumulación temporaria en la propia planta (Luyindula & Weaver, 1989). Harpaz (1975) refiere, como otro aporte de N externo al sistema, el adicionado por las precipitaciones ocurridas durante el ciclo de desarrollo del cultivo.
La relación entre el nitrógeno absorbido desde el suelo y el proveniente de la simbiosis varía con la especie de leguminosa, las condiciones ambientales y las características genéticas del microsimbionte (Shabaev et al., 1996). Aunque las cepas de rizobios nativas presentes en los campos de producción de maní poseen la capacidad de formar nódulos, diversos estudios indican las ventajas de la técnica de inoculación como un medio para alcanzar mejores rendimientos (Reddy et al., 1981; Cholaky et al., 1983; Reddy et al., 1998). Por otra parte, Stalker (1997) ha señalado que el cultivo de maní no produce rendimientos altos en suelos con problemas de fertilidad y que tampoco responde a la fertilización directa.
En la provincia de Córdoba (Argentina), la pérdida de productividad de las tierras de la región manisera y la inestabilidad de los rendimientos de los cultivos asociada, ha sido claramente documentada (Bricchi et al., 1996; Cisneros et al., 1998; Giayetto et al., 1998). Entre los indicadores del deterioro, se destaca la disminución del contenido de materia orgánica en la capa arable del suelo y la consecuente pérdida de estabilidad estructural de los agregados que se relacionan con la formación de costras superficiales (sellado) dificultando el ingreso del agua de lluvia (infiltración disminuida y pérdidas de agua por escurrimiento superficial). Algunos estudios regionales señalan contenidos de materia orgánica del 1% en suelos con uso agrícola continuo (más de 50 años), respecto al contenido original de esos mismos suelos (~3,5%) bajo bosque de algarrobo (Cisneros et al., 1996; Moreno et al., 1996). En ese contexto resulta relevante valorar la contribución relativa de las fuentes de nitrógeno provenientes del suelo y de la atmósfera, vía simbiosis, como base para una estrategia de manejo orientada a la preservación del recurso natural en un esquema productivo.
El objetivo del presente estudio fue evaluar, bajo diferentes condiciones de fertilidad del suelo donde se cultiva maní en la provincia de Córdoba, la influencia de la inoculación de semillas de maní con distintas cepas de Bradyrhizobium sobre el rendimiento del cultivo y estimar la contribución relativa del nitrógeno del suelo y el proveniente de la fijación biológica.
Materiales y métodos
Bacterias y condiciones de crecimiento
Las bacterias ensayadas fueron mutantes espontáneas resistentes a antibióticos, derivadas de inoculantesrecomendados y de un aislamiento nativo: a) Bradyrhizobium sp. SEMIA 6144 inoculante recomendado por MIRCEN-Brasil (mutante espontánea: SEMIA 6144R resistente a estreptomicina, 40 μg mL-1), b) Bradyrhizobium sp. TAL 1000 inoculante recomendado por NifTAL-USA (mutante espontánea: TAL 1000R, resistente a kanamicina, 200 μg mL-1) y c) aislamiento nativo TT002 de la zona manisera de Córdoba (mutante espontánea: TT002R resistente a estreptomicina 200 μg ml-1). La capacidad de las mutantes espontáneas para formar nódulos en plantas de maní fue confirmada por ensayos de invernáculo.
Las cepas bacterianas fueron mantenidas en el medio extracto de levadura-manitol-agar (YEMA) a 4 °C (Vincent, 1970). La pureza se aseguró mediante siembras en el medio YEMA suplementado con rojo Congo y seleccionando las colonias uniformes. El cultivo primario se preparó a partir de una colonia aislada creciendo en 10 ml de medio YEM a 28 °C con agitación hasta alcanzar la fase de crecimiento ogarítmico. Estos cultivos fueron utilizados en la preparación de los inoculantes.
Preparación de los inoculantes
Las mutantes espontáneas resistentes a antibióticos fueron cultivadas en medio YEM hasta alcanzarla fase logarítmica tardía. Los cultivos fueron centrifugados a 10.000 rpm durante 10 minutos y las células suspendidas en 10 mL de medio YEM estéril. La suspensión bacteriana fue colocada en 20 g deturba estéril para conseguir una población de rizobios de 108 ufc g-1 (unidades formadoras de colonias por gramo de turba).
Los estudios experimentales a campo fueron conducidos durante los ciclos agrícolas 1997/98,1998/99 y 1999/00 (en adelante denominados 1998,1999 y 2000, respectivamente) en dos sitios correspondientes a otras tantas localidades de la provincia de Córdoba (Argentina). (i) Experimento 1998: se realizó en el campo experimental de la Universidad Nacional de Río Cuarto, ubicado en la zona rural de la ciudad del mismo nombre; (ii) experimentos 1999 y 2000: tuvieron lugar en un campo de producción agrícola comercial ubicado en la zona rural de General Deheza, 15 km al NO de esa localidad. El suelo del primero de los sitios experimentales fue clasificado como Hapludol típico familia textural limoarenoso(pH: 6,2; materia orgánica: 2,5%, contenido de N-NO3 al momento de la siembra del cultivo de maní: 133 kg ha-1). El suelo de la zona rural de General Deheza corresponde a un Haplustol típico familia textural limo-arenoso (pH: 6,0; materia orgánica:1%, contenido de N-NO3 al momento de la siembra del maní 47 y 25 kg ha-1, para los ciclos 1999 y 2000, respectivamente). No se hallaron registros de práctica de inoculación del cultivo de maní en ninguno de los sitios experimentales del estudio.
La lluvia registrada durante las estaciones de crecimiento del cultivo de maní fue de 797 mm en el ciclo 1998, 563 mm en 1999 y 532 mm en 2000. Estos valores se corresponden con registros promedios para el período noviembre-marzo de series históricas de Río Cuarto (619 mm, período 1974-93) y de General Deheza (587 mm, período 1953-91).
Determinación de la población de rizobios nativos en el suelo
El número más probable (NMP) de rizobios nodulantes de maní presentes en los suelos experimentales, previo a la siembra, fue determinado en los ciclos 1999 y 2000 usando el método de infección en planta. Para ello se tomaron muestras de suelo a una profundidad de 20 cm, las que se diluyeron en agua estéril y se emplearon para inocular semillas de maní sembradas a 1 cm de profundidad en recipientes con vermiculita. Las macetas fueron mantenidas en ambiente controlado (200 μE m-2 sec-1 de intensidad lumínica, ciclo día/noche de 16/8 h y temperatura de 28 °C y humedad relativa de 50%, ambas constantes) y regadas con agua destilada. A las 4 semanas, se determinó el número de plantas que habían formado nódulos y en cada una de ellas el número de nódulos por planta (Somasegaran & Hoben, 1994).
Tratamiento de las semillas, siembra del cultivo y evaluaciones
Previo a la siembra, las semillas de maní del cultivar Tegua (tipo Virginia runner) se desinfectaron con funguicida Vitavax® (mezcla comercial de carboxiny thiram) aplicando una dosis de 250 mL porcada 100 kg de semillas. Se empleó este producto debido a que estudios previos mostraron que no afecta la interacción entre el cultivo de maní y bacteriasdel género Rhizobium, tanto nativas como aquellas provenientes de inoculantes (Fabra et al.,2000). La inoculación de las semillas se realizó aplicandola suspensión de las diferentes cepas enbase turba para alcanzar una densidad estimada de103 células por semilla.
Los tratamientos a evaluar consistieron en semillas inoculadas y no inoculadas (control) asignados a un diseño experimental de bloques al azar con tres repeticiones. Las fechas de siembra fueron: 28 de noviembre de 1997, 6 de noviembre de 1998 y 3 de diciembre de 1999, para los tres ciclos descritos anteriormente. La siembra se realizó en forma manual en hileras separadas a 0,7 m entre sí, con una densidad de 12 semillas por metro lineal de surco. En las etapas fenológicas R1 y R6 (Boote, 1992) se tomaron muestras de cada tratamiento y repetición, y se cosecharon todas las plantas presentes en una superficie de 1 m2, procurando la extracción de la mayor parte del sistema radical. Luego se removió el suelo adherido a las raíces mediante lavado con agua corriente para, posteriormente, separarlas de la parte aérea. Esta última se llevó a estufa de secado con circulación de aire forzado a 40 °C para la determinación de peso seco.
Los nódulos presentes fueron separados de las raíces para llevar a cabo las siguientes determinaciones: a) peso seco en estufa a 70 °C durante 72 h, b) actividad nitrogenasa y c) presencia de las cepas resistentes a antibióticos.
En la etapa R8 del cultivo (madurez de arrancado) se tomaron otras muestras de plantas presentes en 1 m2 por tratamiento y repetición, para determinar la producción de semillas y el rendimiento de maní confitería (peso de las semillas retenidas en zaranda de tajo con orificios oblongos de 7,5 mm de diámetro).
Actividad nitrogenasa
En las etapas R1 y R6 de los experimentos 1998 y 1999, se cuantificó la actividad nitrogenasa según la metodología propuesta por Hardy et al. (1973). Para ello, los nódulos se colocaron en un recipiente cerrado en el que se sustituyó el 10% de su atmósfera por acetileno y se incubó durante 1 hora a 28 °C. Al cabo de ese tiempo se procedió a la medición del etileno producido inyectando un volumen de 0,5 mL de muestra en un cromatógrafo de gases modelo Shimadzu serie GC-6A. Las condiciones del análisis fueron las siguientes: temperatura del detector (H2FID): 100 °C; temperatura del horno: 80 °C; columna de acero de 1m x 2mm empaquetada con Porapak T 80-100.
Ocupación de nódulos
Se tomaron aproximadamente 150 nódulos seleccionados al azar de cada tratamiento, los que fueron esterilizados superficialmente mediante el método de Vincent (1970). Posteriormente, los nódulos fueron macerados individualmente en condiciones asépticas con varilla de madera y resuspendidos en 300 μL de agua destilada estéril. Alícuotas de 10 μL del macerado se sembraron en placas de Petri conteniendo medio YEMA con la adición de los antibióticos empleados para la selección de los distintos mutantes rizobianos y sin ella (Somasegaran & Hoben, 1994). Los resultados se expresaron como porcentaje de nódulos ocupados por la cepa inoculada en relación con el número total de nódulos analizados para cada tratamiento.
Balance relativo de nitrógeno
Antes de la siembra del cultivo de maní y en cada oportunidad de muestreo de plantas (etapas R1, R6 y R8), se tomaron muestras de suelo entre 0-10, 10-20 y 20-40 cm de profundidad en cada sitio experimental y se analizaron para determinar el contenido de N-NO3 - usando el método de la reducción de cadmio (Lambert & Dubois, 1971), y los resultados se expresaron en kg N-NO3 -ha-1 a partir de los datos de densidad aparente de cada capa de suelo.
Paralelamente, porciones de órganos aéreos de las plantas muestreadas (tallos+hojas) se secaron a 40 °C hasta peso constante y molieron para la determinación del contenido de N por el método de micro Kjeldahl (Bremner & Mulvaney, 1982).
Con los datos de N-NO3 - del suelo y de contenido de N de los tejidos vegetales, se calculó el balance relativo de nitrógeno, la contribución de la fijación biológica (FBN) y la variación del N del suelo (VNS), según la metodología utilizada por Cholaky et al. (1986).
FBN = Nplanta (R8) - [(Nsuelo (S) + Nmin + Nlluvia) - Nsuelo (R8)](1)
VNS = [Nsuelo (R8) - ( Nsuelo (S) + Nmin )] + Nrastrojo (2)
Todos los valores se expresan en kg ha-1 y cada uno corresponde a:
Nplanta (R8) = contenido de nitrógeno de la planta en la etapa R8.
Nsuelo (S) = contenido de N-NO3 del suelo a la siembra (S).
Nmin = nitrógeno producido por mineralización de la materia orgánica. Para el cálculo de este componente fue necesario estimar previamente un factor de mineralización mediante la siguiente relación: (Ncultivo de referencia - Nsuelo) / Nmateria orgánica
Donde:
Ncultivo de referencia= es el contenido de N de un cultivo de referencia. En este caso se utilizó maíz sembrado en los mismos sitios experimentales del estudio.
Nsuelo= es el N del suelo calculado como la diferencia entre el contenido a la siembra y el contenido a la cosecha.
Nmateria orgánica= es el contenido de N de la materia orgánica calculado como el 5% de la materia orgánica del suelo.
El valor del factor de mineralización calculado fue de 0,02 y similar para los dos sitios experimentales.
Nlluvia = nitrógeno derivado de las lluvias registradas durante la estación de crecimiento del cultivo, estimado a razón de 2,5 ppm de nitrógeno por mm de lluvia (Harpaz, 1975).
Nsuelo (R8) = contenido de N-NO3 en el suelo a la etapa R8.
Nrastrojo= contenido de nitrógeno en el rastrojo del cultivo de maní después de la cosecha.
Finalmente, se calculó la eficiencia de utilización del N disponible del suelo como la relación: N aportado por el suelo al cultivo / (Nsuelo (S) + Nmin).
Análisis estadísticos
Los datos obtenidos del estudio fueron sometidos a análisis de varianza mediante el programa estadístico InfoStat y los promedios de cada tratamiento comparados según test de Duncan (P<0,05).
Resultados
Ciclo 1998
La condición de fertilidad del suelo de este experimento fue alta (ver descripción del campo experimental de la Universidad Nacional de Río Cuarto) y la cepa empleada como inoculante fue TT002R. El contenido de N-NO3- del suelo disponible al momento de la siembra del cultivo (133 kg ha-1) disminuyó sensiblemente durante la estación de crecimiento del maní, con un valor a la cosecha (etapa R8) de 30 kg ha-1, tanto en el tratamiento inoculado como en el testigo sin inocular. La medición realizada en la etapa R1 mostró un contenido de N-NO3- en el suelo superior en el tratamiento inoculado respecto del control, pero en etapas fenológicas posteriores ambos se equilibraron (Figura 1a). Por otra parte, no se detectaron diferencias significativas en la materia seca acumulada por las plantas y su contenido de N (Figuras 1b y 1c), en la producción de semillas (Figura 1d) y en el rendimiento de maní confitería (Figura 2); así como tampoco se evidenció en el peso seco de los nódulos y en la reducción de acetileno (Tabla 1).
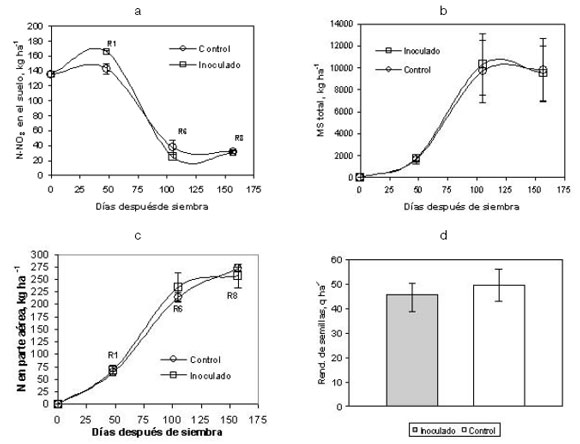
Figura 1. Ciclo 1998: (a) Variación del contenido de N-NO3 en el suelo entre la siembra y la cosecha del cultivo de maní; (b) materia seca acumulada por el cultivo de maní durante la estación de crecimiento; (c) nitrógeno acumulado en la parte aérea de la planta; (d) rendimiento de semillas en el tratamiento inoculado y el control sin inocular. Cada dato es el promedio de tres repeticiones por tratamiento y las líneas verticales indican la desviación estándar del promedio.

Figura 2. Efecto de la inoculación con las cepas TT002R (1998), SEMIA 6144R y TT002R (1999) y TAL 1000R (2000) sobre el rendimiento de maní confitería (porcentaje de semillas retenidas en una zaranda de tajo -orificio oblongo- de 7,5 mm). Cada dato es el promedio de tres repeticiones y las barras verticales indican la desviación estándar del promedio.
Tabla 1. Efecto de la inoculación con la cepa TT002R sobre el comportamiento simbiótico de maní en un suelo de alta fertilidad, ciclo 1998.

Respecto a la contribución relativa de las fuentes de aprovisionamiento de nitrógeno al cultivo, no se detectaron diferencias entre tratamientos (Figura 3a). El suelo aportó, en promedio, 65% del nitrógeno total acumulado por las plantas (264 kg ha-1), la FBN 27% y el 8% restante provino de las lluvias registradas durante la estación de crecimiento.

Figura 3. Nitrógeno total extraído por el cultivo y la contribución de las fuentes de aprovisionamiento del nutriente a la planta de maní en los tratamientos inoculados y en el control sin inocular para los ciclos: a) 1998, b) 1999 y c) 2000. Cada dato es el promedio de tres repeticiones.
La cantidad de nitrógeno extraída por el cultivo (264 kg ha-1, en promedio) reflejó la situación de alta fertilidad del suelo, caracterizada por un elevado contenido de N-NO3- en el momento de la siembra (133 kg ha-1) y el aporte de la mineralización (~70 kg ha-1) y la disponibilidad del nutriente para las plantas favorecida por las buenas condiciones hidrológicas del ciclo (797 mm de lluvia). Aunque no se detectó un efecto significativo de la cepa adicionada como inoculante respecto de las cepas nativas (tratamiento control), se destaca el aporte de la asociación simbiótica (en promedio, 27% del N acumulado por las plantas). Otro aspecto destacable fue que a pesar de la buena disponibilidad de N en el suelo, del aporte por mineralización y de la restitución del nutriente al suelo desde el rastrojo después de la cosecha (57 kg ha-1), el suelo quedó con un balance de N negativo. Esto se corresponde con el requerimiento de N que tiene el maní, señalado en la bibliografía (~ 50 a 60 kg de N tn-1 de grano producido {Gascho, 1992}), y es la razón por la que, generalmente, este cultivo deja el suelo en condiciones deficitarias aun considerando el aporte de la fijación simbiótica como una fuente alternativa de provisión del nutriente.
Las buenas condiciones hidrológicas ya mencionadas favorecieron tanto el desarrollo radical del cultivo como la absorción de N. Una evidencia indirecta de esos efectos lo constituye la eficiencia de utilización del N disponible del suelo que, en promedio, fue del 85%.
Ciclo 1999
Contrariamente a lo señalado para el ciclo anterior, en 1999 la condición de fertilidad del suelo en el experimento realizado en la zona rural de General Deheza fue notoriamente inferior (25 kg ha-1 de NNO 3 al momento de la siembra), a lo que se agregó una muy reducida población nativa de rizobiosnodulantes de maní en el suelo (<50 células/g de suelo). Las cepas de inoculante usadas en este experimento fueron SEMIA 6144R y TT002R. La Figura 4a muestra la variación del contenido de N del suelo durante el ciclo del cultivo para el tratamiento inoculado y el control. No se detectaron efectos significativos de la inoculación sobre la materia seca producida por las plantas, el N acumulado en la parte aérea del cultivo y el rendimiento de semillas (Figuras 4b, 4c y 4d).

Figura 4. Ciclo 1999: (a) Variación del contenido de N-NO3 en el suelo entre la siembra y la cosecha del cultivo de maní; (b) materia seca acumulada por el cultivo de maní durante la estación de crecimiento; (c) nitrógeno acumulado en la parte aérea de la planta; (d) rendimiento de semillas en los tratamientos inoculados y en el control sin inocular. Cada dato es el promedio de tres repeticiones por tratamiento y las líneas verticales indican la desviación estándar del promedio.
La ocupación de los nódulos con las cepas inoculadas fue muy baja (SEMIA 6144R 7% y TT002R 8%) y, aunque no hubo diferencias en los parámetros simbióticos determinados entre plantas provenientes de semillas inoculadas y sin inocular (Tabla 2), se observó un leve incremento en el rendimiento de maní confitería en los tratamientos inoculados (Figura 2), que en promedio alcanzó un 82,4% mientras que en el control sin inocular fue de 74,8%.
Tabla 2. Efecto de la inoculación con cepas SEMIA 6144R y TT002R sobre el comportamiento simbiótico de maní en un suelo de baja fertilidad, ciclo 1999.

La cantidad promedio de nitrógeno extraído por el cultivo al final del ciclo fue de 141 kg ha-1 (equivalenteal 53% del mismo valor cuantificado en el ciclo anterior). Dicha cantidad reflejó la condición deprimida de fertilidad del suelo, la que también se evidenció en el menor aporte de la mineralización (39 kg ha-1) estimada durante la estación de crecimiento (casi la mitad de la calculada en 1998). Paraesta condición de baja fertilidad del suelo, el balance relativo de nitrógeno arrojó un aporte promedio por FBN de 59% (superior al observado en el ciclo 1998), aporte que fue ligeramente superior en el tratamiento inoculado (61%) respecto del control sin inocular (57%). Por su parte, el suelo contribuyó, en promedio, con el 31% del N acumulado en las plantas, y las lluvias con el 10% restante (Figura 3b).
Similar a lo señalado en el ciclo 1998, al momento de la cosecha del maní el suelo presentó déficit de nitrógeno, aunque su magnitud fue marcadamente inferior en este caso, correspondiéndose con una menor eficiencia de utilización del N disponible en el suelo (68%, en promedio).
Ciclo 2000
Este experimento se condujo en el mismo sitio experimental del ciclo anterior (campo de producción comercial ubicado a 15 km al NO de la localidad de Gral. Deheza). Se reprodujo así una condición de baja fertilidad del suelo, reflejada en un contenido de N-NO3- de 47 kg ha-1 al momento de la siembra; en este caso la cepa utilizada como inoculante fue TAL 1000R. La Figura 5a muestra la variación del contenido de N en el suelo desde la siembra hasta la cosecha del maní. Aunque la ocupación de los nódulos con la cepa introducida fue alta (80% en las etapas R1 y R6), las diferencias observadas entre el tratamiento inoculado y el control en la materia seca producida por el cultivo, el nitrógeno acumulado en las plantas de maní, el rendimiento de semillas y los parámetros de la actividad simbiótica, no fueron estadísticamente significativos (Figura 5b, 5c, 5d y Tabla 3).

Figura 5. Ciclo 2000: (a) Variación del contenido de N-NO3 en el suelo entre la siembra y la cosecha del cultivo de maní; (b) materia seca acumulada por el cultivo de maní durante la estación de crecimiento; (c) nitrógeno acumulado en la parte aérea de la planta; (d) rendimiento de semillas en los tratamientos inoculado y en el control sin inocular. Cada dato es el promedio de tres repeticiones por tratamiento y las líneas verticales indican la desviación estándar del promedio.
Tabla 3. Efecto de la inoculación con la cepa TAL 1000R sobre el comportamiento simbiótico de maní en un suelo de baja fertilidad, ciclo 2000.

En el año 2000 no se detectaron diferencias estadísticamente significativas en el rendimiento de maní confitería, en forma similar a lo sucedido en 1998 (Figura 2). Sin embargo, se reprodujo la tendencia a favor del tratamiento inoculado respecto al control como se había detectado en 1999.
Figura 2. Efecto de la inoculación con las cepas TT002R (1998), SEMIA 6144R y TT002R (1999) y TAL 1000R (2000) sobre el rendimiento de maní confitería (porcentaje de semillas retenidas en una zaranda de tajo -orificio oblongo- de 7,5 mm). Cada dato es el promedio de tres repeticiones y las barras verticales indican la desviación estándar del promedio.
El N extraído por el cultivo fue, en promedio de ambos tratamientos, 189 kg ha-1, lo que equivale al 72% de lo acumulado en 1998 y refleja la condición deprimida de fertilidad del suelo cuyo contenido al momento de la siembra fue de 47 kg ha-1 en los primeros 40 cm del perfil. El aporte por mineralización también fue bajo totalizando ~39 kg ha-1.
En cuanto a la contribución relativa de las fuentes de aprovisionamiento de N al cultivo, los resultados se asemejan a los descritos para el ciclo anterior. La FBN aportó entre 57 y 59 % del N acumulado por el cultivo, correspondiéndole entre 33 y 36% al proveniente del suelo y 7-8% a las lluvias (Figura 3c). Como en los años anteriores, el suelo quedó con déficit de N al momento de la cosecha, y la eficiencia de utilización del N disponible tuvo un valor cercano a la de 1999 (75%).
La tabla 4 muestra una síntesis del balance de nitrógeno para los tres experimentos. Resultó evidente la relación entre el contenido de N en el suelo al momento de la siembra, el nivel del nutriente extraído por el cultivo y la eficiencia de utilización del N. Se corresponde, además, con una condición hidrológica favorable (797 mm de lluvia durante el ciclo) que aseguró ausencia de limitaciones para el crecimiento radical y la absorción del N. Hubo también un aporte superior del proceso de mineralización de la fracción orgánica (2,5%), que en conjunto explican la proporción mayor del suministro de N al cultivo proveniente de suelo (65%) y sólo 27% de la FBN. En contraste, los datos de los experimentos 1999 y 2000 muestran una condición de fertilidad del suelo deprimida (25 y 47 kg ha-1 N-NO3 a la siembra, respectivamente) en correspondencia con un menor aporte por mineralización de la materia orgánica (1%) y, en conjunto, una contribución relativa menor al N extraído por el cultivo desde el suelo reflejada en una menor eficiencia de utilización de ese N. Contrariamente al experimento 1998, se destaca en estos dos casos el mayor aporte relativo proveniente de la simbiosis (58,5% , en promedio). No obstante, la cantidad de N extraído por el cultivo fue menor, poniendo en evidencia restricciones a la nutrición mineral expresadas también en el rendimiento de frutos (~ 2 tn ha-1 contra ~ 4,5 tn ha-1 en 1998).
Tabla 4. Balance relativo de nitrógeno de los tres experimentos. Los valores están en unidades de kg ha-1.

Discusión
Desde el punto de vista de la simbiosis, el maní es considerado un cultivo promiscuo (poco específico), ya que establece relación simbiótica con rizobios de diferentes géneros, capaces de infectar a un grupo diverso de leguminosas (Alwi et al., 1989; Taurian et al., 2002). Sin embargo, no todos los microsimbiontes de maní son igualmente efectivos en la fijación de nitrógeno con esta leguminosa. En condiciones de campo, las cepas nativas bien adaptadas a las condiciones edáficas y climáticas del lugar son, en general, de baja capacidad de fijación y/o alta habilidad de nodulación (infectividad), pero pueden competir y dominar a otras cepas altamente eficientes, seleccionadas en condiciones controladas de laboratorio (Martensson et al., 1985). En ese sentido, Thies et al. (1991) informaron que poblaciones indígenas de Rhizobium en concentraciones tan bajas como 50 células por gramo de suelo, fueron capaces de reducir o eliminar la respuesta a la inoculación. Una situación similar se evidenció en el ciclo 1999 de este estudio, donde poblaciones indígenas, en concentración menor a la indicada por Thies et al. (1991), redujeron sensiblemente la ocupación de nódulos por las cepas inoculadas, con lo que demostraron ser más competitivas para formar nódulos que las cepas introducidas (SEMIA 6144R y TT002R), a juzgar por el escaso número de nódulos ocupados por éstas. Por su parte, la cepa TAL 1000R (usada en el experimento del ciclo 2000) pareciera ser más competitiva que las anteriores (SEMIA 6144R y TT002R) bajo la misma condición de fertilidad. Sin embargo, al comparar la contribución relativa de la FBN al nitrógeno total acumulado en la planta en el tratamiento inoculado respecto del control (Figura 3c), la eficiencia de dicha cepa en la captura de N2 resultó similar a la de las cepas nativas.
El principal destino de la producción argentina de maní es la industria alimenticia, por lo que la calidad del producto es un aspecto relevante. Uno de los parámetros para valorar esa calidad es el rendimiento de maní "confitería" (proporción de semillas retenidas en una zaranda de 7,5 mm) asociado con el tamaño de las semillas y su uniformidad. Los resultados de este estudio mostraron que la inoculación con las cepas seleccionadas no mejoró la producción total de semillas en ninguno de los tres ciclos experimentales. No obstante, el rendimiento de maní "confitería" fue 6, 10 y 11% superior en respuesta a la inoculación con cepas seleccionadas en los tres ciclos estudiados, respectivamente. Esas diferencias no fueron estadísticamente significativas; sin embargo, el hecho de haberse presentado en las tres situaciones experimentales involucrando diferentes niveles de fertilidad y condiciones meteorológicas, es una evidencia empírica que soportaría la realización de estudios adicionales para confirmar o desechar ese posible efecto.
La ausencia de efectos significativos sobre la producción de semillas hallada en este estudio es concordante con resultados de otros estudios citados en la literatura. Del mismo modo estos resultados coinciden con antecedentes que reflejan el carácter complementario entre las fuentes de aprovisionamiento de nitrógeno al cultivo (fijado biológicamente y absorbido del suelo) y los cambios relativos que ambas experimentan según la condición de fertilidad del suelo. La absorción de nitrógeno del suelo tiene un costo energético para la planta menor que el requerido para fijarlo biológicamente (Shabaev et al., 1996). Efectivamente, las leguminosas poseen un mecanismo de regulación que limita la fijación biológica de N2 cuando la planta puede satisfacer su demanda desde otra fuente (el suelo, en este caso). En consecuencia, es posible suponer que en condiciones de baja disponibilidad de nitrógeno en el suelo (como la correspondiente a los experimentos de 1999 y 2000), la proporción del nutriente proporcionada por la fijación biológica sería mayor y lo inverso ocurriría en la condición de alta fertilidad. Los resultados obtenidos confirmaron esa presunción, ya que la contribución de la FBN al cultivo de maní representó 59 y 58% del nitrógeno total acumulado en las plantas en los ciclos 1999 y 2000, respectivamente; a su vez, la correspondiente al ciclo 1998 (~27% en promedio) fue significativamente inferior debido a la condición de alta fertilidad inicial que presentó el suelo (~133 kg ha-1 de N-NO3-).
El efecto inhibidor que un alto contenido de N en el suelo tiene sobre la FBN, es un dato conocido en la literatura para un rango de especies leguminosas. Así, Reddy et al. (1998) observaron en el norte de Zambia una disminución relativa de la proporción de N derivado de la FBN en el cultivo de maní, debido a un incremento en el N del suelo. Pero no se registran datos similares de la contribución relativa de este proceso a la nutrición del cultivo de maní en la región central de la provincia de Córdoba.
Existen diferentes métodos para estimar el balance relativo de nitrógeno en condiciones controladas (dilución del isótopo 15N, variedades no nodulantes, entre otros). Sin embargo, para experimentos conducidos en condiciones de campo, como los implementados en este trabajo, el método usado es una alternativa válida para fines comparativos ya que tiene en cuenta las condiciones edáficas y de manejo del cultivo (Cholaky et al, 1986). Fue así que se procedió a monitorear la dinámica del nutriente en el suelo y su acumulación en el cultivo durante el ciclo ontogénico, asumiendo que las pérdidas por volatilización fueron imperceptibles durante el curso de los experimentos. De igual modo, se desestimó la pérdida por percolación profunda en la medida que no se registraron eventos de lluvias de alta intensidad que pudieran haber "lavado" el nutriente por debajo del 1,2 m de profundidad. No obstante las limitaciones del método, a los fines del presente trabajo, disponer de datos cuantitativos del balance relativo de N resultó de valor porque permitió la comparación entre las situaciones experimentales ensayadas.
A pesar de las diferencias en la contribución relativa del nitrógeno del suelo y del aire al balance nutricional del cultivo para las dos condiciones de fertilidad ensayadas, el N acumulado por las plantas crecidas en el suelo con alta concentración del nutriente fue superior al de las que lo hicieron en el sitio con disponibilidad limitante de N. Pero en ambos casos fueron compatibles con el nivel de rendimiento de semillas alcanzado. Este resultado puede ser atribuido a la interacción más favorable entre disponibilidad del nutriente y condiciones hidrológicas registradas en el primero de los sitios experimentales, que pudieron facilitar el desarrollo radical de las plantas de maní y el proceso de absorción del nutriente por el cultivo.
Aunque existen reportes en la bibliografía, los resultados obtenidos en este estudio constituyen una evidencia empírica de la contribución que la fijación simbiótica de nitrógeno hace a la economía de la nutrición nitrogenada del maní, y la manera en que se magnifica en condiciones de un suelo con baja fertilidad o de escasa disponibilidad del nutriente.
Considerando la pérdida progresiva de la productividad de los suelos de la región central de Córdoba durante las dos últimas décadas (departamentos Tercero Arriba, mitad norte de Río Cuarto y este de Juárez Celman), debida, entre otras causas, a la creciente presión de uso agrícola de esas tierras (Bricchi et al., 1996; Cisneros et al., 1998; Giayetto et al., 1998); los resultados expuestos sustentan la hipótesis sobre la posibilidad de obtener rendimientos rentables de maní o soja en sistemas de agricultura continua, sin que ello signifique un equilibrio en el balance nutricional (extraccionesaportes) que continua siendo negativo. Parte de la respuesta es la captura de N que se produce mediante la FBN y su contribución significativa al balance nutricional, en particular cuando se presentan años hidrológicamente normales para la región en cantidad y distribución de lluvias. Sin embargo, como lo muestran estos mismos datos, el aporte de esa fuente alternativa no es suficiente para mantener en equilibrio el balance de N; en consecuencia, resulta necesario considerar la implementación de otras prácticas de manejo destinadas a recuperar el déficit de N del suelo que se viene produciendo.
En tal sentido, la continuidad de estudios orientados a desarrollar inoculantes para este cultivo y la implementación de prácticas de manejo que favorezcan una agricultura sustentable (labranza reducida, rotación de cultivos, por ejemplo), aparecen claramente justificadas. Más aún, al considerar la actual localización del área cultivada con maní en la región sur de la provincia de Córdoba, sobre suelos aptos para el cultivo por su textura arenosa pero de menor desarrollo, baja estabilidad y reducido contenido de materia orgánica donde una producción sustentable sólo es compatible con una estrategia que contemple la preservación del recurso. La inoculación es una herramienta tecnológica apta para esa finalidad y, en general, de un costo comparativamente menor al de otras prácticas como la fertilización.
Agradecimientos
Este trabajo fue subsidiado por la Secretaría de Ciencia y Técnica de la Universidad Nacional de Río Cuarto, el CONICET y la ANPCyT.
Bibliografía
1. Alwi, N.; J.C. Wynne, J.O. Rawlings, T.J. Schneeweis and G.H. Elkan, 1989. Symbiotic relationship between Bradyrhizobium strains and peanut. Crop Science. 29: 50-54. [ Links ]
2. Boote, K., 1992. Growth stages of peanut (Arachis hypogaea L.). Peanut Science. 9: 35-40. [ Links ]
3. Bremner, J.M. and C.S. Mulvaney, 1982. Total nitrogen. Methods of soil analysis. ASA and SSSA, A.L Page, Ed. Madisosn WI. pp.595-624. [ Links ]
4. Bricchi, E.; E. Bonadeo, J.M. Cisneros, J. Marcos, V. Reboyras y W. Hulsken, 1996. Efecto de las labranzas sobre el perfil cultural y la distribución de raíces de maní en un Haplustol típico de Córdoba. XV Congreso Argentino de la Ciencia del Suelo (AACS). Santa Rosa, La Pampa. pp. 197 y 198. [ Links ]
5. Cisneros, J.M.; C. Cholaky, E. Bricchi, J.J. Cantero y O. Giayetto, 1996. Evaluación del efecto del uso sobre las propiedades físicas de un Haplustol típico del centro de Córdoba. XV Congreso Argentino de la Ciencia del Suelo (AACS). Santa Rosa, La Pampa. pp. 23 y 24. [ Links ]
6. Cisneros, J.M.; C. Cholaky, O. Giayetto, E. Bricchi, J. Marcos y G. Cerioni, 1998. Influencia de diferentes sistemas de laboreo sobre la resistencia mecánica de suelos del área manisera. XVI Congreso Argentino de la Ciencia del Suelo (AACS). Villa Carlos Paz, Córdoba. pp. 253-254. [ Links ]
7. Cholaky, L.; O. Giayetto, E.C. Neumann y S. Cavaignac, 1983. Respuesta del maní (Arachis hypogaea L.) a la inoculación al suelo con Rhizobium spp. Rev. UNRC. 3 (2): 173-179. [ Links ]
8. Cholaky, L.; A. Cantero Gutiérrez, O. Giayetto, E. Bonadeo y E.C. Neumann 1986. Fertilización nitrogenada y modelos de siembra en soja de hábito indeterminado: efectos sobre la ontogénesis, nodulación, concentración de nitrógeno, acumulación y distribución de la materia seca, morfología, componentes del rendimiento, rendimientos biológico y económico, índices de cosecha y balance relativo de nitrógeno. Rev. UNRC. 6 (2): 133-166. [ Links ]
9. Fabra, A.; O.Giayetto, G. Cerioni, and S. Castro, 2000. Interaction in the rhizosphere among peanut microsymbionts and a biocontrol agent. FYTON 68: 31-37. [ Links ]
10. Gascho, G.J., 1992. Groundnut (Peanut) Chapter 5.2, in IFA World fertilizer Use manual. Halliday, D.J.; M.E. Trenkel and W. Wichmann, Eds. International Fertilizer Industry Association, Paris. [ Links ]
11. Giayetto, O.; G. Cerioni, W.E. Asnal y J.M. Cisneros, 1998. Rotaciones y labranzas en sistemas agrícolas del centro de Córdoba. Efectos sobre la productividad de cultivos oleaginosos. III Reunión Nacional de Oleaginosas. Bahía Blanca, Buenos Aires. pp. 145-146. [ Links ]
12. Hardy, R.; R. Burns and T. Holsten, 1973. Application of the acetylene reduction assay for the measurement of nitrogen fixation. Soil Biology and Biochemistry 5: 47-81. [ Links ]
13. Harpaz, Y., 1975. Simulation of the nitrogen balance in semi arid regions. Thesis for the Degree Doctor of Philosophy, Hebrew University, Jerusalen, Israel. [ Links ]
14. Lambert, R.J. and R.J. Dubois, 1971. Spectrophotometric determination of nitrate in the presence of chloride. Analytical Chemistry 43: 955-957. [ Links ]
15. Luyindula, N. and R.Weaver, 1989. Nitrogen partitioning in cowpea as influenced by rhizobial strain and mineral nitrogen. Agronomy Journal 81: 593-596. [ Links ]
16. Martensson, A.M.; L.Brutti and H. Ljunggren, 1985. Competition between strains of Bradyrhizobium japonicum for nodulation of soybeans at different nitrogen fertilizer levels. Plant Soil 117:219-226. [ Links ]
17. Moreno, I.; C. Cholaky, M. Lesser y J. Marcos, 1996. Efecto de la labranza sobre el contenido de carbono orgánico y su implicancia en la estabilidad estructural. Actas Resúmenes del XV Congreso Argentino de la Ciencia del Suelo (AACS). Santa Rosa, La Pampa-Argentina: 26a-26b. [ Links ]
18. Reddy, V.M.; J.W. Tanner, R.C. Roy and J.M. Elliot, 1981. The effects of irrigation, inoculants and fertilizer nitrogen on peanuts (Arachis hypogaea L.) II. Yield. Peanut Science 8: 125-128. [ Links ]
19. Reddy, G.B.; A. Mapiki, and B.R. Singh, 1998. Effect of residual fertilizer N, lime and Bradyrhizobium inoculum on groundnut yield, N uptake and N2 fixation. Acta Agriculture Scandinavia. Sect. B Soil Plant. 48:91-99. [ Links ]
20. Shabaev, V.P.; V. Smolin, N.A. Provorov and B.V. Simarov, 1996. The influence of various Rhizobium meliloti strains on lucerne mass and nitrogen accumulation on the mineral nitrogen background. Biology Bulletin. 23:291-296. [ Links ]
21. Simon, T.; S. Kálalova and K.Petrazik, 1996. Identification of Rhizobium strains and evaluation of their competitiveness. Folia Microbiologica 41:65-72. [ Links ]
22. Somasegaran, P. and H.J. Hoben, 1994. Quantifying the growth of rhizobia. Handbook for Rhizobia. Springer Laboratory. New York, Inc., USA. pp. 47-57. [ Links ]
23. Stalker, H., 1997. Peanut (Arachis hypogaea L.). Field Crops Research. 53: 205-217. [ Links ]
24. Taurian, T.; O.M. Aguilar and A. Fabra, 2002. Characterization of nodulating peanut rhizobia isolated from a native soil population in Córdoba, Argentina. Symbiosis 33: 59-72 [ Links ]
25. Thies, J.E.; P.W. Singleton and B.B. Bohlool, 1991. Influence of the size of indigenous rhizobial populations on establishment and symbiotic performance of introduced rhizobia on field-grown legumes. Applied and Environmental Microbiology 57 :19-28. [ Links ]
26. Vincent, J., 1970. A manual for the practical study of root nodule bacteria, in IBP Handbook Nº 15, Blackwell Scientific Publication, Oxford. [ Links ]

