










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkAgriscientia
On-line version ISSN 1668-298X
Agriscientia vol.24 no.1 Córdoba Jan./June 2007
Actividad radical, removilización y absorción de N en trigo en postantesis bajo dos sistemas de labranza y fertilización nitrogenada
Golik, S.I. y H. O. Chidichimo
S.I. Golik. Cerealicultura, Departamento de Tecnología Agropecuaria y Forestal. H. O. Chidichimo. Cerealicultura, Departamento de Tecnología Agropecuaria y Forestal. Facultad de Ciencias Agrarias y Forestales. UNLP. CC 31, 1900. La Plata, Argentina. CIC Pcia. Bs. As. silvinagolik@yahoo.com.ar
Resumen
El objetivo del trabajo fue analizar la actividad radical y su relación con la acumulación, removilización y absorción de N en trigo durante postantesis bajo dos sistemas de labranza y distintas dosis de fertilización nitrogenada. La mayor acumulación de N bajo la labranza convencional se tradujo en un mayor aporte de N al grano (10,36 g m- 2 en 2002 y 10,19 g m- 2 en 2003) y en un mayor rendimiento (485,78 g m- 2 en 2002 y 468,2 g m- 2 en 2003). La siembra directa no presentó diferencias con la labranza convencional en el N absorbido postantesis, pero sí en el removilizado. La fertilización aumentó el N acumulado en cada etapa fenológica, el N removilizado y el N absorbido postantesis. La materia seca radical disminuyó desde antesis hacia madurez. No obstante su tasa de absorción específica de N (SARn) se mantuvo, lo que sugiere que el N acumulado en el grano no sólo proviene del N removilizado sino también del absorbido postantesis. Durante el llenado de granos la MSR no mostró relación con la tasa de absorción de N, pero sí lo hizo su SARn (r= 0,99** y 0,97 ** para 2002 y 2003 respectivamente). Los cultivares presentaron un comportamiento diferencial frente a los distintos tratamientos analizados.
Palabras clave: Nitrógeno; Actividad radical; Absorción; Removilización.
Golik, S.I. and H. O. Chidichimo, 2007. Postanthesis root activity, removilisation and N absortion in wheat under two tillage systems and nitrogen fertilization. Agriscientia XXIV (1): 45-55
Summary
The objective of this work was to evaluate postanthesis root activity and N accumulation, removilisation and absorption under two tillage systems and three N fertilizer levels. The higher N accumulation under the conventional tillage resulted in a greater contribution of N to the grain (10,36 g m- 2 in 2002 and 10,19 g m- 2 in 2003) and a greater yield (485,78 g m- 2 in 2002 and 468,2 g m- 2 in 2003). No-tillage did not present differences with the conventional tillage in the postanthesis N absorbed, but in the remobilised N differences between two tillage systems were observed. The fertilization increased postanthesis N accumulated, N remobilised and N absorbed. Root dry matter decreased from anthesis towards maturity. However, its specific absortion rate of nitrogen (SARn) was remained but also increased during the last analyzed stage, which suggests that N accumulation in the grain not only came to the N remobilised but also postanthesis absorption was important. Root dry matter did not show relation with the absorption rate of nitrogen, but its SARn did. Cultivars presented differential behavior to the different analyzed treatments.
Key words: Nitrogen; Root activity; Absortion; Removilisation.
Fecha de recepción: 15/06/06; fecha de aceptación: 14/06/07
Abreviaturas: MSA, materia seca aérea; MSR, materia seca radical; CT, labranza convencional; SD, siembra directa; SARn, tasa de absorción específica de N; GS, estado de crecimiento; BB, Buck Biguá; BP, Buck Pronto; BBR, Buck Brasil; L, sistemas de labranza; F, tratamientos de fertilización; C, cultivares; Δ N, tasa de acumulación de N.
Introducción
El N se encuentra involucrado en todos los procesos metabólicos de las plantas. Su absorción y partición depende en gran medida de la disponibilidad y demanda durante todo el ciclo del cultivo.
Se conoce que el N absorbido previo a la antesis, en los cereales invernales, representa alrededor del 50 al 75% del N total en la planta a cosecha, y que en condiciones de alta fertilidad del suelo la absorción postantesis resulta igualmente importante porque está positivamente correlacionada con el contenido de N en el grano (Sarandón et al., 1997). Esta absorción se encuentra mucho más influenciada por factores ambientales como la temperatura, humedad y disponibilidad de agua y N (Papakosta & Gagianas, 1991) que la absorción previa a la antesis. En situaciones normales de campo es común que la disponibilidad de N a partir de floración resulte escasa. Si la restricción que impone el agotamiento del N del suelo se supera con ofertas mayores (por ejemplo con aportes por fertilización en dosis fraccionadas) se pueden observar aumentos en la absorción postfloración (Osaki et al., 1997; Delogu et al., 1998; Ruiz, 2001) puesto que el trigo conserva la capacidad de captar N después de antesis (Frederick & Bauer, 1999, Golik et al., 2004). En la medida que la restricción nitrogenada es menos marcada, la curva de acumulación de N sigue la marcha de la acumulación de la materia seca, permitiendo a su vez una mayor duración del área foliar en etapas más avanzadas del cultivo.
Si bien la tasa máxima de crecimiento de las raíces se alcanza en floración (Siddique et al., 1990), y puede decrecer sustancialmente después de ésta, no implica que su actividad, en cuanto a la absorción de nutrientes, no se mantenga más allá de dicho estado fenológico. Aunque poco se conoce acerca del desarrollo y actividad radical bajo condiciones de campo durante el llenado de grano, es probable que la biomasa radical disminuya, sin embargo la tasa de acumulación linear de N en el grano proviene tanto de la absorción activa de N durante el llenado de grano como de la removilización de N de tejidos vegetativos. No obstante, esto podría depender de otros factores tales como el manejo del cultivo y los cultivares utilizados que pueden generar diferentes estrategias de crecimiento y funcionalidad de las raíces.
Se sabe que los tipos de labranzas modifican la dinámica del nitrógeno y en consecuencia inciden sobre la productividad de los cultivos. La siembra directa aparece como una alternativa factible en respuesta a la generalizada degradación de los suelos producida por la intensificación de la agricultura. Sin embargo, en numerosos trabajos se menciona, para un amplio rango de suelos, un menor crecimiento vegetativo de los cultivos bajo este sistema de labranza que se lo atribuye a una mayor compactación (Mahboubi et al., 1993; Chagas et al., 1994) o al menos a un endurecimiento superficial del suelo bajo este sistema de labranza (Taboada et al., 1998; Ferreras et al., 2001). En general la menor temperatura, junto con la mayor humedad y densidad aparente bajo siembra directa producen cambios en las condiciones del suelo que afectan el crecimiento radical, la absorción de nutrientes, el crecimiento aéreo y el rendimiento del cultivo (Qin et al., 2005).
A su vez, bajo sistemas sin laboreo la demanda de N de algunos cultivares puede no estar sincronizada con la disponibilidad en el suelo, y modificar la eficiencia en los procesos de acumulación y partición del N (Sarandón & Caldiz, 1990). Asimismo, y por esta razón, aquellos cultivares con un patrón de acumulación de materia seca y N más precoz se verán más perjudicados en siembra directa que los cultivares de menor precocidad.
El objetivo de este trabajo fue analizar la removilización, absorción y acumulación postantesis de nitrógeno y su relación con actividad radical en trigo durante estados avanzados del cultivo bajo dos sistemas de labranza y distintas dosis de fertilización nitrogenada.
Materiales y métodos
Se realizaron dos ensayos (2002 y 2003) en la Estación Experimental J.J. Hirschhorn (35° S, 58 ° O), La Plata, Facultad de Ciencias Agrarias y Forestales, UNLP, sobre un suelo Arguidol típico con ligeras limitaciones de drenaje interno. El análisis de las muestras arrojó los siguientes resultados a la siembra (0-20 cm): Reja: MO: 3,95 %, P asimilable (Bray Kurtz): 10,8 ppm, Nitratos (ppm): 41,07 Siembra Directa: MO: 4,31 %, P asimilable: 8,2 ppm, Nitratos (ppm): 23,4. MO: 4,21 %, N total: 0,21 %, NO3: 76,6 ppm, P asimilable (Bray Kurtz): 6,7 ppm.
Las fechas de siembra fueron: 31 de julio y 16 de julio respectivamente. Se sembraron tres cultivares de trigo pan, de ciclo similar, intermedio, que difieren en rendimiento, calidad y sanidad: Buck Pingo (BP), Buck Biguá (BB) y Buck Brasil (BBR), bajo dos sistemas de labranza: labranza convencional con arado de reja y vertedera (LC) y siembra directa (SD) en parcelas de 3,8 x 12 m de largo, a una densidad de 300 pl m- 2. Cabe destacar que estos sistemas se vienen realizando sobre el mismo lote desde hace 5 años. En cada sistema de labranza se aplicaron los siguientes tratamientos de fertilización: testigo sin el agregado de N (N0), 40 kg N ha- 1 en la siembra más 40 kg N ha- 1 en 2-3 nudos detectables (N80) y 80 kg N ha- 1 en la siembra más 80 kg N ha- 1 en 2-3 nudos detectables (N160) en forma de urea. A todas las parcelas se les aplicó 100 kg ha- 1 de superfosfato triple (0-46-0) en el momento de la siembra.
El ensayo quedó formado por dos tratamientos de labranza (LC y SD), 3 cultivares (BP, BB y BBR) y tres tratamientos de fertilización, según un diseño de parcelas divididas con tres repeticiones, donde las labranzas constituyeron la parcela principal, los cultivares las subparcelas y los tratamientos de los fertilización las subsubparcelas.
Durante antesis, cuaje, grano lechoso, grano pastoso y madurez, estados de crecimiento (GS) 65 (30/10 en 2002 y 23/10 en 2003) , GS 69 (12/11 en 2002 y 3/11 en 2003), GS 77 (22/11 en 2002 y 17/11 en 2003), GS 85 (5/12 en 2002 y 16/12 en 2003) y GS 95 (15/12 en 2002 y 2003), respectivamente (Zadoks et al., 1974), se obtuvo la biomasa aérea (MSA) a partir de la extracción de una superficie de 0,3 m2 (sectores de 0,5 m de tres surcos contiguos) y su posterior secado en estufa a 70 °C durante 48 hs.
La extracción de raíces, para las mismas etapas, se realizó con un barreno de 9,8 cm de diámetro y 10 cm de altura. En cada subsubparcela las muestras de suelo se tomaron a intervalos de 10 cm hasta los 40 cm de profundidad. Se tomaron 4 muestras en el surco y 4 en el entresurco. Las 8 muestras se juntaron en una única muestra. Las muestras se guardaron en frío a -18°C y se procesaron en laboratorio, lo mas pronto posible, mediante lavados sucesivos con agua y hexametafosfato de sodio como dispersante (Acciaresi et al., 2001), filtrado con mallas de 480 μm y separación manual de restos vegetales, hasta obtener la muestra de raíces limpias. Se secaron en estufa a 70 °C y se pesaron, obteniéndose la materia seca radical (MSR) en g m-2.
Se determinó el % de N por microKjeldahl, para las distintas etapas fenológicas consideradas previa división del material en paja y espiga y en paja, granza y granos en madurez. A partir del porcentaje de N y la materia seca correspondiente, se determinó el N acumulado en g m-2.. También se determinó: a) la absorción de N postfloración (g m-2.) como la diferencia en el contenido de N de la planta total a madurez y a floración, b) el N removilizado (g m-2.) como la diferencia entre el N de la planta entera a floración y el N contenido en la paja a madurez.
Para estimar la actividad radical en cada etapa fenólogica, se utilizó la tasa de absorción específica de N (SARn, mg N día- 1( g- 1 materia seca de raíz) propuesta por Osaki et al. (1995), calculada como la relación entre la tasa de acumulación de N y el peso seco promedio de las raíces, para cada período analizado.
Los resultados se procesaron con un análisis de varianza y las medias se compararon según la prueba de Tukey al 0,05 de probabilidades. Además se analizaron las posibles correlaciones entre las variables planteadas.
Debido a la presencia de efectos significativos de los años sobre la acumulación de N y a la presencia de interacciones con los años, se realizó un análisis por separado de cada año.
Resultados y discusión
Si bien las lluvias medias mensuales en marzo, abril y mayo, durante el año 2002, resultaron abundantes respecto a las medias históricas, las lluvias durante la implantación del cultivo fueron, en cuanto a cantidad y distribución, similares. En el año 2003 resultaron menores a las históricas, excepto para el mes de noviembre; no obstante atendieron los requerimientos del cultivo (Figura 1).

Figura 1. Datos climáticos
Acumulación, removilización y absorción de N postantesis
La acumulación de N, el N removilizado y el N absorbido postantesis, fueron afectados en forma diferente por los sistemas de labranza y los tratamientos de fertilización en las etapas fenológicas analizadas tanto en 2002 como en 2003.
Se conoce que la productividad del cultivo se encuentra fuertemente regulada por la acumulación de N, y que esta relación se mantiene estable frente a diversas condiciones ambientales. En este trabajo hubo una alta asociación entre el N acumulado y la MSA para los dos años analizados (Figura 2). Sin embargo, dicha asociación resultó mayor en 2002 (r=0,92** en 2002 y r=0,87** en 2003), lo que podría estar asociado a las abundantes lluvias registradas durante el mes de noviembre en 2003 (Figura 1) y a la pérdida de nitratos por lixiviación en dicho año. En 2002 la acumulación de N no presentó diferencias significativas por efecto de las labranzas en GS 65, GS 69 y GS 77. Sí hubo diferencias en GS 85 y GS 95. En 2003, el efecto de las labranzas sobre la acumulación de N fue significativo en GS 65, GS 69, GS 77 y en GS 95 (Tabla 1). La alta correlación encontrada, para ambos sistemas de labranza, entre el N acumulado en el cultivo y la biomasa aérea total (para LC, promedio de ambos años: r= 0,86** en GS 65, r= 0, 90** en GS 69, r= 0,89** en GS 77, r= 0,91** en GS 85, r= 0,91** en GS 95; para SD, promedio de ambos años: r=0,95** en GS 65, r= 0,90** en GS 69, r= 0,92** en GS 77, r=0,88** en GS 85, r= 0,93** en GS 95), coincide con resultados anteriores (Sarandón et al., 1997) y sugiere que la menor acumulación de N en SD (11,5 y 10,6 g m- 2 para LC y SD, respectivamente en el año 2002 y 10,9 y 9,5 g m- 2 para LC y SD, respectivamente en el año 2003) podría relacionarse con impedimentos en la acumulación de biomasa. Entre estos la mayor resistencia mecánica del suelo en SD (Ferreras et al., 2001), que coincide con la mayor densidad del suelo y la resistencia a la penetración encontrada en estas parcelas por Golik et al. (2004) y con el menor desarrollo radicular en SD respecto al desarrollo en LC (Sarandón et al., 1997). Alakukku & Elonen (1995) afirman que la absorción de N por el cultivo sería un parámetro sensible a la compactación del suelo y citan disminuciones en la acumulación de N entre un 4 y 9%, según el tipo de suelos. Por otra parte, la compactación puede retrasar la mineralización del N e incrementar las pérdidas de N gaseoso del suelo. A su vez, se ha citado que la desnitrificación bajo SD en los primeros 10 cm de suelo puede ser responsable de hasta el 55% de las diferencias en el contenido de nitratos con respecto a LC, en suelo similares al de este ensayo (Rodríguez & Giambiagi, 1995). La mayor acumulación de N bajo la labranza convencional se tradujo en un mayor aporte de N al grano (10,4 y 9,6 g m- 2 bajo LC y SD, respectivamente en el año 2002 y 10,2 y 8,6 g m- 2 bajo LC y SD, respectivamente en el año 2003). La SD si bien no presentó diferencias con la labranza convencional en el N absorbido postantesis, sí lo hizo en el N removilizado en el año 2003 (Tabla 2). Esto podría estar asociado a la menor disponibilidad temprana de dicho nutriente bajo el sistema no convencional de laboreo.
Tabla 1: Análisis de la varianza del N acumulado en 3 cultivares de trigo en 5 estados de crecimiento (GS) durante 2 años, bajo 2 sistemas de labranza tratamientos de fertilización
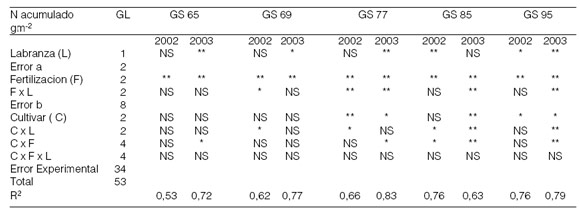

Figura 2. Acumulación de materia seca aérea (MSA) g m- 2 y N g m- 2, en 4 estadios de crecimiento (GS): 69: antesis, 77: cuaje, 85: grano lechoso y 95: madurez, para dos años 2002 y 2003.
Tabla 2: Análisis de la varianza del N grano, rendimiento, N removilizado y N absorbido postantesis en 3 cultivares de trigo durante 2 años, bajo 2 sistemas de labranza y 3 tratamientos de fertilización

Los tratamientos de fertilización presentaron efecto significativo, en 2002 y 2003, sobre la acumulación de N en todas las etapas consideradas (Tabla 2), incrementándose el N acumulado con dosis crecientes de N aplicado (6,9; 8,5 y 14,8 g m-2 para N0, N80 y N160, respectivamente en el año 2002 y 5,6; 11,2 y 13,8 g m-2 para N0, N80 y N160, respectivamente en el año 2003). El aporte del N removilizado hacia la espiga del acumulado en los tejidos vegetativos previo a la antesis y del absorbido postantesis se incrementó con el agregado de fertilizante, tanto en 2002 (3,5; 5,3 y 6,9 g m-2 bajo N0, N80 y N160, respectivamente para el N removilizado y 4,1; 6,6 y 8,2 g m-2 bajo N0, N80 y N160, respectivamente para el N absorbido postantesis) como en 2003 (2,33; 5,28 y 6,44 g m-2 bajo N0, N80 y N160, respectivamente para el N removilizado y 2,5; 5,6 y 7,6 g m-2 bajo N0, N80 y N160, respectivamente para el N absorbido postantesis). Los resultados del presente trabajo coinciden con los encontrados por Delogu et al. (1998) que hallaron un incremento en el aporte al grano tanto del N removilizado como del absorbido postantesis trabajando con aplicaciones de altas dosis de N (140 a 210 kg N ha-1) y no con los de Palta & Fillery (1993) y Golik et al. (2003), que encontraron que la aplicación de N en todos los tratamientos fertilizados provocó un incremento en la contribución a la espiga solamente del N removilizado y no de ambos, removilizado y absorbido postantesis. Probablemente esto sea debido a que las dosis aplicadas en estos últimos dos casos fueron similares y relativamente más baja que las aplicadas por Delogu et al. (1998) y en el presente trabajo. A su vez, hubo diferencias significativas para las distintas dosis aplicadas. El agregado de una dosis más alta en el último tratamiento permitió mantener una mayor disponibilidad de dicho nutriente para el cultivo, que se puso de manifiesto en la acumulación realizada por él.
Entre los cultivares no se observaron diferencias durante GS 65 y GS 69 tanto en 2002 como en 2003, en la acumulación de N (Tabla 1). A partir de allí sí presentaron diferencias significativas (a excepción en GS 85 en el año 2002), siendo BBR el que presentó menor acumulación en GS 77 en ambos años analizados respecto a los otros dos cultivares (11,4; 11,8 y 9,8 g m-2 para BP, BB y BBR respectivamente, en el año 2002 y 9,6; 9,8 y 8,4 g m-2 para BP, BB y BBR respectivamente, en el año 2003) y en GS 85 en 2003 (11,8; 10,3 y 9,5 g m-2 para BP, BB y BBR respectivamente) y en GS 95 en 2002 respecto a BP (15; 13,7 y 14 g m-2 para BP, BB y BBR respectivamente). Esto tuvo una alta asociación con el N en grano (r=0,97** para ambos años). No obstante los cultivares no presentaron diferencias significativas en el N removilizado y en el absorbido postantesis en 2002 ni en 2003 (Tabla 2).
Interacciones: Se considera que existe un mayor déficit de N en el suelo en SD que en los sistemas de labranza convencional (Doran, 1980). Por lo tanto, debería observarse una mayor respuesta a la fertilización nitrogenada en SD que en LC (Darroch & Fowler, 1990). Cuando hubo interacciones, sin embargo, la respuesta a la fertilización nitrogenada en este ensayo fue menor, sugiriendo la existencia de otros factores condicionantes, además del probable déficit de N en el suelo (Figura 3a). Los cultivares tuvieron un comportamiento diferencial ante los sistemas de labranza y los tratamientos de fertilización en lo relacionado con la acumulación y partición del N (Figura 3b). Se ha considerado que aquellos cultivares con mayores tasas de crecimiento, es decir, más precoces en cuanto a la acumulación del N total, deberían verse más perjudicados en SD que en LC. Los datos de este ensayo señalan que BB fue el que mejor respondió frente a la labranza conservacionista y a la aplicación de dosis creciente de N. BP y BBR se vieron perjudicados en condiciones de SD, indicando una menor adaptación respecto a BB a dichas condiciones.
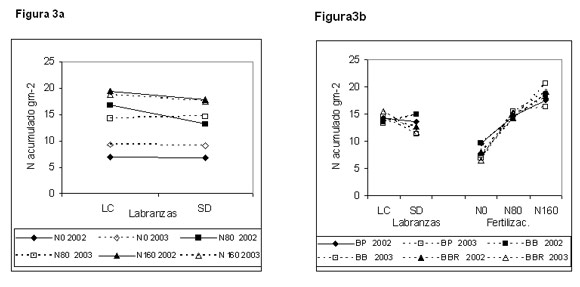
Figura 3. Interacciónes para el N acumulado g m-2 (promedio para los estados de crecimiento con diferencia estadísticamente significativa, para dos años : 2002 y 2003). Labranza (LC y SD) x fertilización (N0, N80 y N160) (Figura 3a); labranza (LC y SD) x cultivar (BP, BB y BBR) y fertilización (N0, N80 y N160) x cultivar (BP, BB y BBR) (Figura 3 b).
Respecto al N en grano y al N acumulado postantesis se observaron las mismas respuestas, para las interacciones entre cultivares y labranzas, y entre cultivares y tratamientos de fertilización (Figuras 4 y 5), que para el N acumulado, lo que indica la fuerte asociación existente entre dichos parámetros (r=0,97** para N en grano y N total acumulado para ambos años; r= 0,76** en el 2002 y r= 0,81** en 2003 para N en grano y N absorbido postantesis). No hubo interacción entre labranza y fertilización para ninguno de los dos parámetros analizados.

Figura 4. Interacciones para N acumulado postantesis g m-2 (promedio para los estados de crecimiento con diferencia estadísticamente significativa, para dos años 2002 y 2003). Labranza (LC y SD) x cultivar (BP, BB y BBR) y fertilización (N0, N80 y N160) x cultivar (BP, BB y BBR) .

Figura 5. Interacciones para N en grano g m-2 (promedio para los estados de crecimiento con diferencia estadísticamente significativa, para dos años 2002 y 2003). Labranza (LC y SD) x cultivar (BP, BB y BBR) y fertilización (N0, N80 y N160) x cultivar (BP, BB y BBR) .
Biomasa y actividad radical
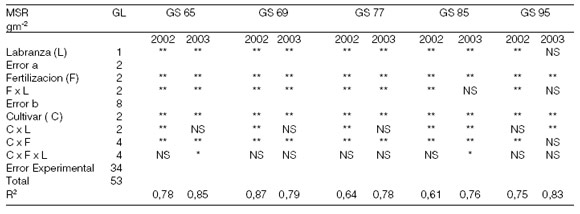
Se encontraron diferencias estadísticamente significativas en la producción de biomasa radical para los sistemas de labranza como para los tratamientos de fertilización y los cultivares, tanto en 2002 como en 2003 (Tabla 3). Resultó mayor bajo labranza convencional (promedio de los cinco estados fenológicos considerados: 183 y 164,76 g m-2 para LC y SD, respectivamente en el año 2002, y 181,2 y 140,7 g m-2 para LC y SD, respectivamente en el año 2003) y en los tratamientos con aplicación de N (promedio para los cinco estados fenológicos analizados: 144,2; 177,5 y 199,9 g m-2 para N0, N80 y N160, respectivamente en el año 2002, y 116,5; 160,7 y 205,6 g m-2 para N0, N80 y N160, respectivamente en el año 2003). Entre los cultivares, BP y BB presentaron menor MSR que BBR, principalmente durante GS 65, GS 69 y GS 77; a partir de allí hubo un comportamiento más dispar entre los cultivares (promedio para los cinco estados fenológicos analizados: 168,1; 174,8 y 178,8 g m-2 para BP, BB y BBR, respectivamente en el año 2002 y 147,8; 141,5 y 193,3 g m- 2 para BP, BB y BBR, respectivamente en el año 2003).
Tabla 3: Análisis de la varianza de la MSR en 3 cultivares de trigo en 5 estados de crecimiento (GS) durante 2 años, bajo 2 sistemas de labranza y 3 tratamientos de fertilización.
La SD provocó una menor producción de raíces en relación a la LC. A su vez, con fertilización nitrogenada la SD no alcanzó el rendimiento de la labranza convencional (455,8 y 441,4 g m-2 para LC y SD, respectivamente en el año 2002, y 468,2 y 399,3 g m-2 para LC y SD, respectivamente en el año 2003), lo que indicaría que además del N puede haber otros factores que limitarían el rendimiento del trigo. Esto pudo estar asociado a la mayor densidad aparente y resistencia a la penetración encontrada para este sistema de labranza por Mahboubi et al. (1993), Chagas et al. (1994) y Golik et al. (2004). Qin et al. (2005) encontraron una mayor densidad del suelo en SD que en LC, entre los 0 y 25 cm de profundidad, que explicaría la distinta respuesta, observada por ellos, en la distribución del sistema radical del maíz bajo dichos sistemas de laboreo.
La MSR, promedio para cada etapa, presentó su mayor valor en GS 65, para luego disminuir durante GS 69 y GS 77, presentar un aumento durante GS 85 y mostrar su menor valor durante GS 95 en el 2002. En 2003 también presentó su mayor valor en GS 65, disminuyendo progresivamente hacia la madurez del cultivo, a excepción de un pequeño aumento durante GS 85 (Figura 6). Si bien con el agregado de N mejoró su producción, en términos generales disminuyó desde antesis hacia madurez. Los aumentos de materia seca radical observados en algunos estados fenológicos entre GS 65 y GS 95, pueden ser atribuidos a una producción tardía de macollos asociados a las condiciones ambientales dadas durante floración y el llenado de granos, tal lo sugerido por Abayomi and Wright (1999). Dicha producción tardía de macollos fue observada pero no cuantificada debido a su escasa contribución en el rendimiento del cultivo. No obstante su SARn se mantuvo e incluso aumentó durante la última etapa analizada (GS 95) (Figura 6), lo que sugiere que el N acumulado en el grano por el cultivo no sólo proviene del N removilizado sino también del absorbido postantesis. Esto coincide con lo sugerido por Osaki et al. (1997) y Ruiz (2001) y queda corroborado por la correlación positiva hallada entre la SARn y el N absorbido postantesis (r=0,55** y 0,76** para 2002 y 2003, respectivamente), e indica que la actividad radical es un proceso que puede mantenerse durante estados avanzados del cultivo bajo buenas condiciones de fertilidad del suelo. La SARn presentó diferencias significativas para los sistemas de labranzas (Tabla 4), en GS 95 de 2002 (0,00017 y 0,0021 mg N día-1 g de raíz-1 para LC y SD, respectivamente) y en GS 69 (0,0003 y 0,0012 mg N día-1 g de raíz-1 para LC y SD, respectivamente), GS 77 (0,0004 y 0,0007 mg N día-1g de raíz-1 para LC y SD, respectivamente), GS 85 (0,0003 y 0,0011 mg N día-1 g de raíz-1 para LC y SD, respectivamente) de 2003. Esto tuvo una alta relación con la menor producción de MSR logrado bajo SD, en dichos estados fenológicos. En general la SARn presentó diferencias significativas para los tratamientos de fertilización (Tabla 4), siendo mayor con el agregado de fertilizante (promedio de los cinco estadios fenológicos considerados: 0,0006; 0,0007 y 0,001 mg N día-1 g de raíz-1 para N0, N80 y N160, respectivamente en 2002 y 0,0006; 0,0018 y 0,002 mg N día-1 g de raíz-1 para N0, N80 y N160, respectivamente en 2003). Entre los cultivares BB fue el que presentó mayor SARn, considerando la totalidad de los estados analizados (0,0009; 0,001 y 0,0006 mg N día-1 g de raíz-1 para BP, BB y BBR, respectivamente en el año 2002, y 0,001; 0,0022 y 0,0019 mg N día-1 g de raíz-1 para BP, BB y BBR, respectivamente en el año 2003).

Figura 6. Materia seca radical (MSR) g m-2 y tasa de absorción específica de N (SARn) mg N día-1 g-1 raíz en 5 estados de crecimiento (GS), años 2002 y 2003.
Tabla 4: Análisis de la varianza de la tasa de absorción específica de N (SARn) en 3 cultivares de trigo en 4 estados de crecimiento (GS) durante 2 años, bajo 2 sistemas de labranza y 3 tratamientos de fertilización.

Durante el llenado de granos la MSR no mostró relación con la tasa de absorción de N, pero sí lo hizo su SARn (r=0,99** y 0,97 para 2002 y 2003, respectivamente). Esto coincide con lo hallado por Shinano et al. (1994), respecto a que la tasa de absorción de N está relacionada con SARn y con la tasa de crecimiento del cultivo y no con la MSR, y por Osaki et al. (1995), quienes consideran que la tasa de acumulación de N en trigo se encuentra estrechamente relacionada con SARn, mientras que la MSR permanece constante o decrece con el tiempo.
Interacciones: en los cultivares actuales se asume que la absorción de N postfloración puede permanecer alta, por lo tanto se considera que la habilidad de las raíces para absorber N después de antesis es un factor importante que controla la productividad de los cultivos. Cuando hubo interacciones, en general se observó una menor respuesta a la fertilización, en cuanto a la producción de MSR, bajo SD que bajo LC (Figura 7a). Entre los cultivares, BB fue el que presentó menor MSR bajo SD. BBR fue el de mayor MSR en SD y el de mayor respuesta frente al agregado de N. BP mostró un comportamiento intermedio (Figura 7b).

Figura 7. Interacciones (promedio para los estados de crecimiento con diferencia estadísticamente significativa) para la materia seca radical (MSR, en g m-2), para dos años 2002 y 2003. Labranza (LC y SD) x fertilización (N0, N80 y N160) (Figura 7a) ; labranza (LC y SD) x cultivar (BP, BB y BBR) y fertilización (N0, N80 y N160) x cultivar (BP, BB y BBR) (Figura 7b) .
Respecto a la tasa de absorción específica de N, BB fue el que presentó mayor actividad específica bajo SD, asociado a su mayor acumulación de N y a su menor MSR bajo este sistema conservacionista de labranza. Asimismo fue el que presentó mejor respuesta frente a la aplicación de dosis crecientes de fertilizante. BBR resultó el cultivar más afectado en cuanto a su actividad especifica bajo SD. BP mostró un comportamiento intermedio (Figura 8). No hubo interacción para los sistemas de labranza y los tratamientos de fertilización.

Figura 8. Interacciones para la tasa de absorción específica de N mg N día -1 g raíz -1 (promedio para los estados de crecimiento con diferencia estadísticamente significativa, para dos años 2002 y 2003). Labranza (LC y SD) x cultivar (BP, BB y BBR) y fertilización (N0,N80 y N160) x cultivar (BP, BB y BBR).
Conclusiones
Existieron diferencias en el N acumulado y en el removilizado entre sistemas de labranzas, que se tradujeron en el contenido de N en el grano y en el rendimiento del cultivo.
La fertilización aumentó el N acumulado en cada etapa fenológica, el N removilizado y el N absorbido postantesis.
Si bien la materia seca radical disminuyó durante el llenado de granos, su SARn no, lo que indica que la actividad radical es un proceso que puede mantenerse durante estados avanzados del cultivo. Esto sugiere que el N acumulado en el grano no sólo proviene del N removilizado sino también del absorbido postantesis. Durante el llenado de granos la MSR no mostró relación con la tasa de absorción de N, pero sí lo hizo su SARn.
Entre los cultivares, BB fue el que presentó mayor actividad específica bajo SD, asociado a su mayor acumulación de N y a su menor MSR bajo este sistema de labranza. BBR fue el de mayor SARn bajo LC y el mas afectado en su actividad bajo SD. BP mostró un comportamiento intermedio respecto a los otros dos cultivares.
Bibliografía
1. Abayomi, Y.A. and D. Wright, 1999. Effects of water stress on growth and yield of spring wheat (Triticum aestivum L) cultivars. Tropical Agriculture 76 (2): 120-125. [ Links ]
2. Acciaresi H.A.; H.O. Chidichimo and S.J. Sarandón, 2001. Traits related to competitive ability of wheat (Triticum aestivum) varieties against Italian Ryegrass (Lolium multiflorum). Biological Agriculture and Horticulture 19: 275- 286. [ Links ]
3. Alakukku, L. and P. Elonen. 1995. Longterm effects of a single compaction by heavy field traffic on yield and nitrogen uptake of annual crops. Soil & Tillage Research 36: 141-152. [ Links ]
4. Chagas, C.I.; H.J. Marelli y O.J. Santanatoglia, 1994. Propiedades físicas y contenido hídrico de un Argiudol típico bajo tres sistemas de labranza. Ciencia del Suelo 12: 11-16. [ Links ]
5. Darroch, B.A. and D.B. Fowler. 1990. Dry matter production and nitrogen accumulation in no- till winter wheat. Canadian Journal of Plant Science 70: 461-472. [ Links ]
6. Delogu, G; L. Cattivelli, N. Pecchioni, D. De Falcis, T. Maggiore and A.M. Stanca, 1998. Uptake and agronomic efficiency of nitrogen in winter barley and winter wheat. European Journal of Agronomy 9: 11-20. [ Links ]
7. Doran, J.W.; 1980. Soil microbial and biochemical changes associated with reduced tillage. Soil Science Society of America Journal 44: 765-771. [ Links ]
8. Ferreras, L.A.; J.J. De Battista, A. Ausilio y C. Pecorari, 2001. Parámetros físicos del suelo en condiciones no perturbadas y bajo laboreo. Pesquisa Agropecuaria Brasileira. 36 (1): 161-170. [ Links ]
9. Frederick, J.R. and P.J. Bauer, 1999. Ecology and Physiology of Yield Determination. In Wheat. Editors: Emilio H. Satorre & Gustavo A. Slafer. Chapter 3, pp. 45-65. [ Links ]
10. Golik, S.I.; H.O. Chidichimo, D. Pérez y L. Pane, 2003. Acumulación, removilización, absorción postantesis y eficiencia de utilización de n en trigo bajo diferentes labranzas y fertilizaciones. Pesquisa Agropecuaria Brasileira, 38 (5): 619-626. [ Links ]
11. Golik, S.I.; J.I. Zanelli, A. Oliveira y H.O. Chidichimo, 2004. Biomasa y removilización y absorción de N en trigo en postantesis. XXV Reunión Argentina de Fisiología Vegetal, Santa Rosa, La Pampa: 70. [ Links ]
12. Mahboubi, A.A.; R. Lal and N.R. Faussey, 1993. Twentyeight years of tillage effects on two soils in Ohio. Soil Science Society of America Journal.57: 506-512. [ Links ]
13. Osaki, M.; T. Shinano, M. Matsumoto, J. Ushiki, M.M. Shinano, M. Urayama, and T. Tadano, 1995. Productivity of high- yielding crops V. Root growth and root activity. Soil Sci. Plant Nutrition 41 (4): 635-647. [ Links ]
14. Osaki, M.; T. Shinano, M. Matsumoto, T. Zheng and T. Tadano, 1997. A root-shoot interaction hypotesis for high productivity of field crops. Plant Nutrition -for sustainable food production & environment: 669-674. [ Links ]
15. Palta, J.A. and R.P. Fillery, 1993. Postanthesis remobilisation and losses of nitrogen in wheat in relation to applied nitrogen. Plant and Soil 155/156: 179-181. [ Links ]
16. Papakosta, D.K. and A.A. Gagianas, 1991. Nitrogen and dry matter accumulation, remobilization, and losses of Mediterranean wheat during grain filing. Agron. Journal 83: 864-870. [ Links ]
17. Qin, R.; P. Stamp and W. Richner, 2005. Impact of tillage and banded starter fertilizer on maize root growth in the top 25 centimeters of the soil. Agron. Journal 97: 674- 683. [ Links ]
18. Rodríguez, M.B. and N. Giambiagi, 1995. Denitrification in tillage and no tillage pampean soils: relationships among soil water, available carbon, and nitrate ans nitrous oxide production. Communications in Soils Science Plant Analysis 26: 3205-3220. [ Links ]
19. Ruiz, R.A, 2001.Trigo, cuaderno de actualización técnica 63, (3): 24- 33. [ Links ]
20. Sarandón, S.J. and D.O. Caldiz, 1990. Effects of varying nitrogen supply at different growth stages on nitrogen uptake and nitrogen partitioning efficiency in two wheat cultivars. Fertilizer Research, Dordrecht, v. 22: 21-27. [ Links ]
21. Sarandón, S.J.; S.I. Golik y H.O. Chidichimo, 1997. Acumulación y partición del N en dos cultivares de trigo pan ante distintos momentos de fertilización nitrogenada bajo siembra directa y labranza convencional. Rev. Fac. Agr. La Plata, 102 (2): 175-186. [ Links ]
22. Siddique, K.H.M.; R.K. Belford and D. Tennant, 1990. Root:shoot ratios of old and modern, tall and semi dwarf wheats in a mediterranean environment. Plant and Soil 121: 89-98. [ Links ]
23. Taboada, M.A.; F.G. Micucci, D.J. Cosentino and R.S. Lavado, 1998. Comparison of compaction induced by conventional and cero tillage in two soils of the Rolling Pampa of Argentina. Soil & Tillage Research 49: 57-63. [ Links ]
24. Zadoks, J.C.; T.T. Chang and C.F. Konzak, 1974. A decimal code for the growth stages of cereals. Weed Research 14: 415- 421. [ Links ]

