










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgriscientia
versión On-line ISSN 1668-298X
Agriscientia vol.28 no.2 Córdoba jul./dic. 2011
ARTÍCULOS
Abundancia y riqueza específica de pulgones (Hemiptera: Aphididae) y sus parasitoides en diferentes genotipos y estados fenologicos del garbanzo
Mazzuferi, V. E.; A. Maidana, P. Fichetti, L. G. Hansen y D. S. Avalos
V. E. Mazzuferi, A. Maidana, P. Fichetti, L. G. Hansen y D. S. Avalos. Zoología Agrícola, Facultad de Cs. Agropecuarias-UNC. C.C. 509, 5000 Córdoba. Argentina. Correspondencia a V. E. Mazzuferi: vimazzu@agro.unc.edu.ar
RESUMEN
Los áfidos son insectos perjudiciales tanto por la succión de savia como por la transmisión de enfermedades. Los objetivos de este trabajo fueron conocer las especies de áfidos y sus parasitoides presentes en diferentes genotipos de garbanzo, calcular la incidencia del parasitismo e informar sobre las fluctuaciones poblacionales de pulgones y parasitoides en relación a los estados fenológicos del cultivo. El estudio se realizó durante 2006 y 2007 en San Marcos Sierras (Córdoba). Se evaluaron siete líneas y dos cultivares sembrados en un diseño en bloques completamente aleatorizados con tres repeticiones. Cada 15 días se colectaron manualmente áfidos y "momias" presentes en 10 plantas seleccionadas al azar, de cada genotipo y bloque. Se determinaron 13 especies de áfidos. En 2006 se observó mayor abundancia y riqueza de especies que en 2007. Las especies numéricamente dominantes fueron: Acyrthosiphon pisum y A. kondoi. Solamente cinco especies colonizaron el cultivo. El cultivar Chañaritos presentó la mayor abundancia de insectos en los dos años agrícolas, y los genotipos M60 y M75 mostraron la mayor riqueza de especies. En ambos años, los pulgones estuvieron presentes durante la etapa reproductiva del cultivo. Se registraron tres especies de parasitoides y se estimó un 35% de parasitismo total.
Palabras clave: Cicer arietinum; Genotipos; Acyrthosiphon pisum; Acyrthosiphon kondoi; Parasitoides.
Species richness and abundance of aphids (Hemiptera: Aphididae) and their parasitoids in different genotypes and at different growth stages of chickpea
SUMMARY
Aphids may damage plants by sucking plant juices or by transmitting diseases. The aim of this study was to obtain further information about aphid species and their parasitoids in different genotypes of chickpea, calculate the incidence of parasitism and report on fluctuating populations (from aphids and parasitoid) in relation to plant phenology. The study was undertaken during 2006 and 2007 in San Marcos Sierras (Cordoba). Seven lines and two cultivars of chickpea were evaluated. Crop plots were planted following a completely randomized design block with 3 replicates. Every two weeks different developmental stages of aphids and "mummies" found in 10 plants randomly selected from each genotype and block were collected manually. Thirteen species of aphids were identified. In 2006 there was greater abundance and species richness than in 2007. The numerically dominant species were Acyrthosiphon pisum and A. kondoi. Only 5 species colonized the crop. Chañaritos presented the greatest abundance of insects in both years and M60 and M75 genotypes showed the greatest species richness. In both years, aphids were present during the reproductive stage. There were three species of parasitoids and parasitism was estimated at about 35%.
Key words: Cicer arietinum; Genotypes; Acyrthosiphon pisum; Acyrthosiphon kondoi parasitoids.
Fecha de recepción: 09/03/11;
fecha de aceptación: 07/11/11
INTRODUCCIÓN
Los áfidos se citan entre las plagas más importantes de las regiones templadas (Davies, 1991). Su gran capacidad de multiplicación les permite alcanzar altas densidades poblacionales (Artigas Coch, 1994). Estos insectos no sólo succionan savia de hojas tiernas, pedúnculos de flores y frutos, sino que, además, son vectores de enfermedades virales.
Los pulgones citados sobre garbanzo, a nivel mundial, corresponden a las especies Aphis craccivora Koch., Acyrthosiphon pisum (Harris) (Sharma et al., 2007) y Myzus persicae Sulzer (Jones et al., 2008); el primero es señalado como importante vector de diferentes tipos de virus en la India (Kaiser et al., 1990) y África (Reed et al., 1986).
Las poblaciones de estos insectos son afectadas por diversas especies de entomopatógenos, depredadores y parasitoides (Starý, 1976). Entre estos últimos se citan con frecuencia a los integrantes de la subfamilia Aphidiinae (Hymenoptera: Braconidae). El estudio de este grupo reviste gran interés por ser utilizados como agentes de control biológico (Tizado y Núñez Pérez, 1991). A nivel mundial, las principales especies que parasitan a los pulgones pertenecen a los géneros Aphidius y Diaeretiella (Michelena et al., 2004; Kos et al., 2008). En la India se cita a Trixys indicus (Subbarao & Sharma) y Lipolexix scutellarris Mackaur (Hymenoptera: Aphidiinae) como importantes parasitoides de Aphis craccivora en garbanzo (Singh & Tripathi, 1987).
En la Argentina, diferentes especies de Aphidiinae son citados como parasitoides de pulgones que se alimentan de fabáceas cultivadas y plantas hortícolas (Aragón e Imwinkelried, 1995; Berta et al., 2002; Andorno et al., 2007). No obstante, muy poco se conoce aún sobre este grupo de insectos en nuestro país (Berta et al., 2002).
Entre los métodos de control de plagas, además del control biológico y químico, el genético, con los genotipos resistentes, constituye una herramienta eficaz particularmente para el manejo de los insectos sucsopicadores. Las interacciones insectoplanta presentan una amplia gama de intensidades, que va desde las plantas que se comportan como el hospedante óptimo hasta aquellas que son totalmente inadecuadas (Maxwel y Jennings, 1984). Caracteres de la planta tales como arquitectura, densidad de tricomas, dureza y tamaño de las hojas, contenido químico y sustancias secundarias afectan la preferencia alimentaria de los insectos y los diferentes aspectos del desarrollo (Rossiter et al., 1988; Zangerl, 1990) disminuyendo también la habilidad para predadores y parasitoides (Fritz, 1995).
En la Argentina los estudios sobre la entomafauna asociada a garbanzo son recientes (Avalos et al., 2008). En relación a los pulgones, colectas realizadas sobre este cultivo permitieron identificar algunas especies, destacándose particularmente por su abundancia Acyrthosiphon pisum y Acyrthosiphon kondoi Shinji (Mazzuferi et al., 2009; Avalos et al., 2010) y dos especies de parasitoides: Aphidius ervi Haliday y A. colemani Viereck (Avalos et al., 2010). Sin embargo, aún no existen registros sobre las fluctuaciones poblacionales de los áfidos y sus parasitoides en relación a los estados fenológicos del cultivo, comportamiento de diferentes genotipos frente a sus infestaciones y eficiencia de parasitismo, entre otros.
A partir de trabajos de investigación para la mejora genética de este cultivo en la Argentina, se obtuvieron los cultivares Chañaritos S-156 y Norteño (Biderbost y Carreras, 1991; Carreras et al, 1997). Actualmente se evalúan líneas procedentes de distintos cruzamientos, a los fines de obtener nuevos cultivares que puedan adaptarse a diversas zonas según su respuesta a factores bióticos y abióticos y manejos culturales.
Desde este marco teórico se propone que la presencia y la abundancia de pulgones y sus parasitoides, difieren entre los diversos genotipos.
Los objetivos de este trabajo fueron ampliar la información sobre las especies de áfidos y sus parasitoides presentes en diferentes genotipos de garbanzo, calcular la incidencia del parasitismo e informar sobre las fluctuaciones poblacionales de pulgones y parasitoides en relación a los estados fenológicos del cultivo en el NO de la provincia de Córdoba.
MATERIALES Y MÉTODOS
Ubicación de parcelas experimentales El estudio se llevó a cabo en parcelas ubicadas en la localidad de San Marcos Sierras (Dpto. Cruz del Eje) en la provincia de Córdoba, durante los años 2006 y 2007.
Diseño de siembra
Se sembraron siete líneas (MxWR-315: RILs- 14 (M14); MxWR-315: RILs-19 (M19); MxWR-315: RILs-25 (M25); MxWR-315: RILs-59 (M59); MxWR- 315: RILs-60 (M60); JG-62xILC-72: RILs-64 (J64) y MxWR-315: RILs-75 (M75)) y dos cultivares (Chañaritos S-156 y Norteño) de garbanzo en un diseño en bloques completamente aleatorizados, con tres repeticiones. Cada parcela estuvo formada por cuatro surcos de 2,5 m de largo y 0,7 m entre ellos, donde se colocaron 10 semillas por metro. En ambas campañas, la siembra se realizó en la segunda quincena de mayo. El cultivo se manejó bajo prácticas agrícolas convencionales, sin la aplicación de insecticidas.
Recolección de insectos
Se realizaron muestreos quincenales desde la emergencia del cultivo hasta madurez de cosecha. En cada parcela, mediante observación directa y con la ayuda de un pincel, se recolectaron pulgones en diferentes estados de desarrollo, como así también los pulgones parasitados o "momias" presentes en 10 plantas tomadas al azar de cada genotipo.
Los pulgones se conservaron en alcohol al 70% para su posterior identificación, y las "momias" en tubos de Khan hasta la emergencia de los himenópteros parasitoides cuyos adultos fueron debidamente montados y rotulados para su identificación.
Procesamiento y análisis de datos
Sobre la base del material colectado, se identificaron los áfidos en el laboratorio de Zoología Agrícola de la Facultad de Ciencias Agropecuarias de la U.N.C mediante claves (Remaudiére y Seco Fernandez, 1990; Delfino, 1991; Artigas Coch, 1994; Ortego, 1994); a su vez, los parasitoides fueron determinados en el Instituto M. Lillo. En ambas campañas agrícolas, los ejemplares que no pudieron ser correctamente determinados se agruparon como no identificados. Se calculó la proporción relativa aportada por cada especie y se analizaron sus fluctuaciones temporales. Se evaluó la abundancia de pulgones presentes en los diferentes genotipos, y se establecieron las asociaciones entre las diferentes especies de áfidos y parasitoides con las líneas y los cultivares sembrados.
El porcentaje de parasitismo se calculó a partir de la siguiente fórmula:
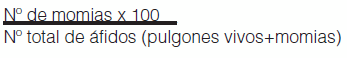
Para comparar la abundancia de áfidos entre diferentes genotipos se realizó una regresión de Poisson. Se analizó cada año por separado. Se utilizó el programa estadístico INFOSTAT (versión 2008).
RESULTADOS
Pulgones
Considerando ambas campañas agrícolas se recolectaron en total 206 pulgones (153 ejemplares vivos + 53 momias). Se registraron 13 especies pertenecientes a 10 géneros (Tabla 1). Cabe destacar que sólo se observaron ninfas y ejemplares ápteros de cinco especies (Tabla 1).
Tabla 1. Especies de pulgones (Hemiptera: Aphididae) recolectadas durante 2006 y 2007 en San Marcos Sierras (Córdoba), las que colonizaron el cultivo y clasificación según rango de hospedantes.

Del total de especies registradas solamente Acyrthosiphon kondoi, A. pisum y Therioaphis trifolii se hallaron en ambas campañas agrícolas (Tabla 1).
Campaña 2006
De los 98 ejemplares de áfidos recolectados en esta campaña, 66 fueron identificados y clasificados en 11 especies pertenecientes a 8 géneros (Tabla 1). El género Acyrthosiphon representó la mitad del total de los ejemplares colectados y la especie más abundante fue A. pisum (Figura 1).
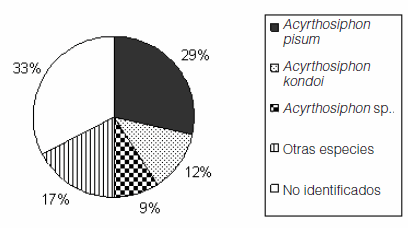
Figura 1. Abundancia relativa de las especies de pulgones registrada sobre garbanzo durante 2006 en San Marcos Sierras (Córdoba).
Campaña 2007
En este año se registraron solamente 55 ejemplares de pulgones, 41 de los cuales fueron clasificados en cinco especies pertenecientes a cuatro géneros (Tabla 1). El género Acyrthosiphon representó alrededor del 60% del total de los ejemplares colectados y A. kondoi fue la especie numéricamente dominante (Figura 2).

Figura 1. Abundancia relativa de las especies de pulgones registrada sobre garbanzo durante 2007 en San Marcos Sierras (Córdoba).
Parasitoides
Los datos corresponden a la campaña 2006, ya que en 2007 no se registraron parasitoides. Se recolectaron en total 53 momias de pulgones. En algunos casos éstas se encontraban vacías, indicando que los parasitoides ya habían emergido. De las acondicionadas en laboratorio se criaron en total 16 ejemplares correspondientes a 3 especies que pertenecen a 2 géneros de la familia Braconidae (Aphidiinae) (Tabla 2).
Tabla 2. Especies de parasitoides de pulgones registradas en garbanzo en San Marcos Sierras (Córdoba) y clasificación según rango de hospedantes.

El género Aphidius fue el mejor representado numéricamente y la especie A. ervi la más numerosa (Figura 3). Si bien no pudieron identificarse todas las especies de áfidos, de las 16 momias acondicionadas, pudo establecerse que la asociación más frecuente correspondió a Acyrthosiphon spp. - Aphidius ervi.
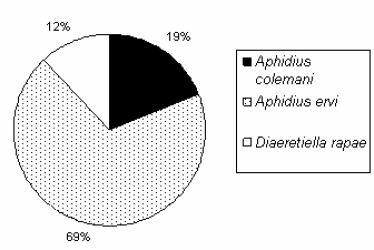
Figura 3. Abundancia relativa de las especies de parasitoides registrada durante 2006 en San Marcos Sierras (Córdoba).
Fluctuación de la abundancia de pulgones y porcentaje de parasitismo
Campaña 2006
Los primeros ejemplares de pulgones se registraron a mediados del invierno (primeros días de agosto), coincidiendo con la etapa de comienzos de floración del cultivo, y se los recolectó hasta la primavera (fines de setiembre) durante el período de floración-fructificación. El pico de abundancia ocurrió en la segunda quincena de agosto coincidiendo con la etapa de floración (Figura 4). Las tres especies más abundantes halladas durante esta campaña fueron Acyrthosiphon kondoi, Acyrthosiphon sp. y A. pisum. Las dos primeras especies se registraron con similar abundancia en la segunda quincena de agosto, coincidiendo con la etapa de floración del cultivo. A. pisum se presentó en bajo número en agosto; la máxima abundancia se observó a mediados de septiembre, coincidiendo con la etapa de floración-fructificación, y declinó posteriormente hasta desaparecer un mes después (Figura 5).

Figura 4. Fluctuación de la abundancia de pulgones y porcentaje de parasitismo en diferentes estados fenológicos del garbanzo en 2006, en San Marcos Sierras (Córdoba).

Figura 5. Fluctuación poblacional de las principales especies de áfidos asociadas a garbanzo, durante 2006, en San Marcos Sierras (Córdoba).
Los parasitoides se hallaron presentes durante todo el período en que se encontraron los áfidos. En general, cuando el porcentaje de parasitismo fue alto la abundancia de pulgones fue baja y viceversa. El porcentaje de parasitismo fue máximo a comienzos de agosto y septiembre. Hacia fines de septiembre la población de pulgones declinó y también el porcentaje de parasitismo (Figura 4). Durante esta campaña se registró un 35% de parasitismo total.
Campaña 2007
Los primeros ejemplares de áfidos comenzaron a encontrarse a partir del mes de septiembre. En la primera semana se observó la máxima abundancia de insectos, pico que coincidió con el estado fenológico de floración (Figura 6).

Figura 6. Fluctuación de la abundancia de pulgones en diferentes estados fenológicos del garbanzo en 2007 en San Marcos Sierras (Córdoba).
Comparando entre campañas agrícolas se observó que el pico de abundancia de áfidos ocurrió en agosto en el año 2006 y un mes después en 2007 (Figuras 4 y 6).
Las principales especies halladas fueron Acyrthosiphon kondoi, A. pisum y Aphis craccivora. La primera mostró el máximo de abundancia en la primera semana de septiembre y continuó registrándose en bajo número sobre el cultivo hasta fines del mes de octubre. Las dos últimas especies solamente se presentaron en la primera semana de septiembre, y estos picos poblacionales coincidieron con la etapa de floración del cultivo (Figura 7).
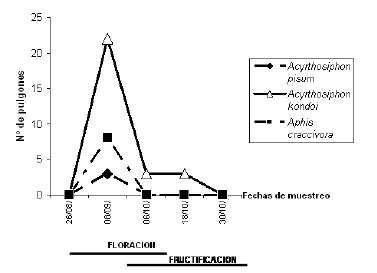
Figura 7. Fluctuación poblacional de las principales especies de áfidos asociadas a garbanzo, durante 2007, en San Marcos Sierras (Córdoba).
Comparación de abundancia de pulgones entre los diferentes genotipos
Al analizar la abundancia de pulgones presentes en los diferentes genotipos en la campaña 2006, la regresión de Poisson arrojó diferencias significativas fundamentalmente al tomar al cultivar Chañaritos como referencia. Si bien pudo observarse que todos los genotipos presentaron menor número de afidos que el cultivar, sólo se establecieron diferencias estadísticamente significativas con los materiales M14 (p<0,0010), M19 (p<0,0006), M25 (p<0,0086) y M59 (p< 0,00141). Para los restantes genotipos los valores hallados fueron: Norteño (p<0,0742), M75 (p<0,5795), M60 (p<0,3190), J64 (p<0,3973). En el análisis de la campaña 2007, si bien el cultivar Chañaritos mostró mayor abundancia de áfidos que los restantes genotipos, no se establecieron diferencias estadísticamente significativas.
Riqueza de especies y asociación con los genotipos
Pulgones
La mayor riqueza específica de pulgones se registró durante la campaña agrícola 2006, ya que de las 13 especies encontradas en este estudio, 11 estuvieron presentes en este año (Tabla 1). Teniendo en cuenta ambas campañas agrícolas, los genotipos M60 y M75 reunieron el mayor número de especies de pulgones, puesto que se hallaron asociados a siete y seis respectivamente. En los restantes genotipos se encontraron sólo dos o tres especies, salvo el genotipo M25 que estuvo asociado a cuatro (Tabla 3). Acyrthosiphon pisum y A. kondoi mostraron el mayor rango de hospedantes al encontrarse presentes en siete de los nueve genotipos estudiados. Le siguieron en orden de importancia Acyrthosiphon sp. y Aphis craccivora en tres genotipos; R. padi, especie que no coloniza el cultivo, se registró en cuatro (Tabla 3).
Tabla 3. Especies de pulgones asociadas a los diferentes genotipos de garbanzo durante 2006 (X) y 2007 (![]() ) en San Marcos Sierras, Córdoba.
) en San Marcos Sierras, Córdoba.

Parasitoides
El genotipo M60 fue el único que reunió a dos especies de parasitoides de las tres registradas en la campaña 2006; en los demás genotipos sólo se encontró una especie (Tabla 4). Aphidius ervi parasitó pulgones presentes en seis de los nueve genotipos estudiados. Le siguieron en orden de importancia A. colemani, que se registró en dos, y Diaeretiella rapae, sólo en un genotipo (Tabla 4).
Tabla 4. Especies de parasitoides asociadas a pulgones presentes en los diferentes genotipos de garbanzo durante 2006 en San Marcos Sierras, Córdoba.

DISCUSIÓN
Se encontraron en total 13 especies de pulgones asociadas a garbanzo. A nivel mundial solamente tres de ellas han sido citadas sobre este cultivo: Acyrthosiphon pisum, Aphis craccivora (Kaiser et al., 1990; Sharma et al., 2007) y Myzus persicae (Jones et al., 2008). Un número superior de áfidos y mayor riqueza de especies se encontraron en la campaña 2006. En 2007 recién comenzaron a observarse ejemplares de pulgones un mes después que en la campaña anterior. Tanto la abundancia como la diversidad de especies pueden estar afectadas por las condiciones climáticas particulares de los diferentes años agrícolas. Existen evidencias de que las diferentes especies de áfidos requieren determinadas condiciones de temperatura para reproducirse (Artigas Coch, 1994; Hodgson, 2007). Las condiciones adversas del invierno de 2007 pueden haber retardado la aparición y el desarrollo de las colonias de pulgones e inhibido la presencia de algunas especies menos adaptadas a las bajas temperaturas. Según información registrada en varias provincias argentinas, incluyendo a Córdoba, el 9 de julio de 2007 se produjo una intensa nevada a la que se sumaron temperaturas frías durante los meses de agosto y septiembre. Esta combinación de condiciones puso en evidencia que el invierno de 2007 resultó mucho más frío que el de 2006 (http://blog.nuestroclima.com).
También las diferentes condiciones climáticas de los años de estudio pueden haber influido en la presencia de la especie más abundante encontrada en cada campaña agrícola. En 2006 se destacó Acyrthosiphon pisum, en cambio en 2007 dominó en número A. kondoi. Según Artigas Coch (1994), las infestaciones de A. pisum se ven favorecidas por inviernos suaves. Por debajo de los 12ºC esta especie tiene un desarrollo muy lento y las hembras pueden permanecer en estado juvenil hasta 6 semanas y debajo de 5º C no se reproducen. En cambio, A. kondoi se adapta mejor a los inviernos fríos produciendo ninfas a partir de 3ºC.
Por otra parte, solamente se encontraron hembras ápteras y ninfas de Acyrthosiphon kondoi, A. pisum, Aphis craccivora y Terioaphis trifolli. De acuerdo a la literatura consultada, se conoce que los pulgones tienen ciclos biológicos relativamente complejos y presentan individuos con diferentes formas, que cumplen distintas funciones. Cuando las condiciones climáticas del invierno son compatibles con la supervivencia de los pulgones y éstos cuentan con recursos alimenticios disponibles, realizan un ciclo anholociclico (Barbagallo et al., 1998). En estos ciclos, las hembras ápteras vivíparas son las encargadas de multiplicar una colonia de áfidos. Las formas aladas, en cambio, se ocupan de colonizar nuevos hospedantes (Agarwala, 2007). Considerando estas características, puede inferirse que las cuatro especies citadas son capaces de colonizar el cultivo. Dichas especies son también las que se encuentran en otras fabáceas y particularmente en alfalfa con mayor frecuencia (Delfino, 1991; Aragón e Imwinkelried, 1995; Cordo et al., 2004). Las restantes, de las que sólo se recolectaron individuos alados (formas migrantes), probablemente se hallaban realizando vuelos de reconocimiento y picaduras de prueba al momento de tomar las muestras.
Resulta relevante señalar la asociación áfidosgarbanzo ya que la mayoría de ellos son importantes vectores de enfermedades virales, con diferentes modalidades de transmisión. Así, las cuatro especies de pulgones que colonizaron el cultivo en este estudio se hallan citadas como transmisoras de virus en fabáceas (Hodgson, 2007), y Aphis craccivora, en particular, es un efectivo transmisor de diversas enfermedades víricas a garbanzo en la India (Kaiser et al., 1990) y África (Reed et al., 1986). Las formas migrantes son capaces de transmitir virus de manera no persistente. Jones & Coutts (1996) comprobaron que el virus del mosaico de las cucurbitáceas (CMV) es transmitido, a plantas de garbanzo, por diferentes especies de áfidos migrantes, que no se establecen en el cultivo. A su vez, Jones et al. (2008) encontraron que, a pesar de la ausencia de colonización de los áfidos, muchos genotipos se infectan rápidamente con CMV en condiciones de campo.
Tanto por presentarse en ambas campañas agrícolas como por su abundancia y capacidad de colonizar el cultivo, puede señalarse a Acyrthosiphon pisum y A. kondoi como las especies más importantes en garbanzo. En este sentido, estos datos difieren de lo observado en la India, donde la principal especie de pulgón por su frecuencia de aparición es Aphis craccivora (Sharma et al., 2007).
Las especies que parasitaron los áfidos fueron Aphidius colemani, A. ervi y Diaeretiella rapae. Todas se encontraron durante la campaña 2006. Aphidius ervi fue la especie dominante, lo que podría explicarse por el predominio de ejemplares de Acyrthosiphon spp. encontrados en este estudio ya que Zuazúa et al. (2003) señalan que A. ervi parasita con preferencia a A. pisum. La especie A. ervi fue introducida en la Argentina en 1973 y 1978 para el control de A. pisum y A. kondoi sobre cultivos de leguminosas (Berta et al., 2002).
La ausencia de parasitoides en 2007 puede deberse a la menor abundancia de hospedadores como consecuencia de las condiciones climáticas adversas que se presentaron en este año, según se expuso en párrafos anteriores. La cita de Diaeretiella rapae, mencionada en este estudio, sumada a las especies Aphidius ervi y A. colemani ya publicadas por este equipo de investigación (Avalos et al., 2010), constituyen los primeros registros en el sistema garbanzo-pulgones-parasitoides en la Argentina.
Los picos de abundancia de las dos principales especies, Acyrthosiphon pisum (en 2006) y A. kondoi (en 2007) ocurrieron durante las etapas reproductivas del cultivo, fundamentalmente durante la floración. Respecto a A. pisum, lo observado en este estudio se corresponde con lo registrado por Sharma et al., (2007) quienes establecen que esta especie se alimenta de flores y vainas en garbanzo.
Si bien en esta investigación los pulgones no alcanzaron densidades poblacionales elevadas, como es frecuente observar en estos insectos (Artigas Coch, 1984), hubo diferencias entre los distintos genotipos, tanto en lo relacionado a la abundancia cuanto a la riqueza de especies asociadas. Algunos autores sostienen que el garbanzo presenta pocos insectos porque las plantas poseen pelos glandulares y el cultivo se realiza en invierno (Van Der Maesen, 1972; De Miguel Gordillo, 1991). En relación a los pulgones, se ha registrado que algunas variedades de garbanzo que poseen abundantes tricomas no son colonizadas por los áfidos, o que éstos se alimentan exitosamente en plantas, o áreas de la planta, donde los tricomas no se encuentran (Edwards, 2001; Sharma et al., 2007).
En este estudio, dos de las tres especies de parasitoides encontradas se hallaron asociadas a pulgones presentes en el genotipo M60; coincidentemente esta línea y la M75 exhibieron la mayor riqueza de especies de áfidos, pero no la mayor abundancia. Se halla documentado que además de la presencia de tricomas, caracteres de la planta tales como arquitectura, dureza y tamaño de las hojas, contenido químico y sustancias secundarias afectan la preferencia alimentaria de los insectos y diferentes aspectos de su desarrollo (Rossiter et al., 1988; Zangerl, 1990) disminuyendo también la habilidad para predadores y parasitoides (Fritz, 1995).
En la campaña 2006, al incremento de los pulgones en agosto le siguió una respuesta numérica de los parasitoides en septiembre, lo que puede indicar que estos microhimenópteros contribuyen a la regulación de las poblaciones de áfidos en garbanzo. En este sentido, los resultados de esta investigación son consistentes con lo observado en otros sistemas que incluyen áfidos y parasitoides en árboles cultivados (Zuparko & Dahlsten, 1994) y huertas de Brassicacea (Duchovskiene & Raudonis, 2008).
A partir de los resultados encontrados, se abren nuevos interrogantes que plantean futuros estudios sobre las diferentes causas (morfológicas, físicas o químicas) que influyen en los procesos de selección y éxito en la colonización o no de los diversos genotipos.
AGRADECIMIENTOS
Los autores agradecen al Ing. E. Orecchia por permitir efectuar los ensayos en su establecimiento; a la Dra. C. Berta (Instituto M. Lillo, Tucumán) por identificar los parasitoides y a la SECyT (U.N.C), por subsidiar esta investigación.
BIBLIOGRAFÍA
1. Agarwala, B. K. 2007. Phenotypic plasticity in aphids (Homoptera: Insecta): Components of variation and causative factors. Current Science 93 (3): 308-313. [ Links ]
2. Andorno, A. V., S. N. López y E. N. Botto. 2007. Asociaciones áfido-parasitoide (Hemiptera: Aphididae; Hymenoptera: Braconidae, Aphidiinae) en cultivos hortícolas orgánicos en Los Cardales, Buenos Aires, Argentina. Rev. Soc. Entomol. Arg. 66 (1-2): 171-175. [ Links ]
3. Aragón, J. R. y J. M. Imwinkelried. 1995. Plagas de la alfalfa. En: Hijano, E. y A. Navarro (eds.), La alfalfa en la Argentina, INTA Subprograma Alfalfa, pp. 81-104. [ Links ]
4. Artigas Coch, J. N. 1994. Familia Aphididae. En: Entomología Económica, Insectos de Interés Agrícola, Forestal, Médico y Veterinaria. Ediciones Universidad de Concepción, Concepción, Chile, pp. 481-629. [ Links ]
5. Avalos, S.; P. Fichetti; V. Mazzuferi; C. Berta y J. Carreras. 2008. Entomofauna asociada a diferentes genotipos de garbanzo en la región central de Argentina. En: Resúmenes XXXI Congreso Argentino de Horticultura. Mar del Plata, Argentina, pp.395. [ Links ]
6. Avalos, S.; V. Mazzuferi; P. Fichetti; C. Berta y J. Carreras. 2010. Entomofauna asociada a garbanzo en el noroeste de Córdoba (Argentina). Horticultura Argentina 29 (70): 5-11. [ Links ]
7. Barbagallo, S.; P. Cravedi; E. Pasqualini y J. Patti. 1998. Pulgones de los principales cultivos forestales. Mundi- Prensa. Bayer. [ Links ]
8. Berta D. C.; M. V. Colomo y N. E. Ovruski. 2002. Interrelaciones entre los áfidos colonizadores del tomate y sus himenópteros parasitoides en Tucumán (Argentina). Bol. San. Veg. Plagas 28: 67-77. [ Links ]
9. Biderbost, E. B. y J. Carreras. 1991. Chañaritos S-156 nuevo cultivar de garbanzo (Cicer arietinum L.). Agriscientia 8: 15-18. [ Links ]
10. Carreras, J.; S. García; N. Gray; N. Collavino; V. Failde; C. Pastrana y E. Biderbost. 1997. Norteño: un cultivar de garbanzo para exportación. En: Actas del XX Congreso Argentino de Horticultura., Bahía Blanca, Argentina, pp. 126. [ Links ]
11. Cordo, H. A.; L. Logarzo; K. Braun y O. Di Iorio. 2004. Catálogo de insectos fitófagos de la Argentina y sus plantas asociadas. Sociedad Entomológica Argentina Ediciones. Buenos Aires. [ Links ]
12. Davies, R. J. 1991. Introducción a la Entomología. Mundi- Prensa, Madrid. [ Links ]
13. Delfino, M. A. 1991. Reconocimiento de los áfidos (Homoptera: Aphidoidea) encontrados en cultivos de alfalfa (Medicago sativa) en la Argentina. Rev. Fac. Agron. FCA y Forest. UNLP. 66 y 67. [ Links ]
14. De Miguel Gordillo, E. 1991. El garbanzo. Una alternativa para el secano. Mundi-Prensa, Madrid. [ Links ]
15. Duchovskiene, L. and L. Raudonis. 2008. Seasonal abundance of Brevicoryne brassicae L. and Diaeretiella rapae (M'Instosh) under different cabbage growing systems. Ekologija 54 (4): 260-264. [ Links ]
16. Edwards, O. R. 2001. Interspecific and intraspecific variation in the performance of three pest aphid species on five grain legume hosts. Entomol. Exp. Appl. 100: 21-30. [ Links ]
17. Fritz, R. S. 1995. Direct and indirect effects of plant genetic variation on enemy impact. Ecol. Entomol. 20: 18-26. [ Links ]
18. Hodgson, E. W. 2007. Aphids in alfalfa. Utah University Extension and Utah Plant Pest Diagnostic Laboratory, 5 pp. [ Links ]
19. InfoStat 2008. Manual del Usuario. Grupo InfoStat, FCA, Universidad Nacional de Córdoba. Edit. Brujas Argentina. [ Links ]
20. Jones, R. A. and B. A. Coutts. 1996. Alfalfa mosaic and cucumber mosaic virus infection in chickpea and lentil: incidence and seed transmission. Ann. Appl. Biol. 129: 491-506. [ Links ]
21. Jones, R. A.; B. A. Coutts; L. J. Latham and S. J.Mckirdy. 2008. Cucumber mosaic virus infection of chickpea stands: temporal and spatial patterns of spread and yield-limiting potential. Plant Pathol. 57: 842-853. [ Links ]
22. Kaiser, W. J.; A. M. Ghanekar; Y. L. Nene; B. S. Rao and V. Anjaiah. 1990. Viral diseases of chickpea. Chickpea in the Nineties. En: II International Workshop Chickpea Improvement, Hyderabad, India, pp. 139-142. [ Links ]
23. Kos, K.; . Tomanovic; O. Petrovic-Obradovic; . Laznik; M. Vidrih and S. Trdan. 2008. Aphids (Aphididae) and their parasitoids in selected vegetable ecosystems in Slovenia. Acta Agric. Slov. 91: 15-22. [ Links ]
24. Maxwell, F. G. y P. R. Jennings. 1984. Mejoramiento de plantas resistentes a insectos. Edit. Limusa, México, D. F. [ Links ]
25. Mazzuferi, V.; A. Maidana; S. Avalos; P. Fichetti y J. Carreras. 2009. Los pulgones (Hemiptera: Aphididae) presentes en genotipos diferentes de garbanzo. En: Resúmenes XXXII Congreso Argentino de Horticultura. Salta, Argentina. p. 358. [ Links ]
26. Michelena J. M.; P. González y E. Soler. 2004. Parasitoides afidiinos (Hymenoptera, Braconidae, Aphidiinae) de pulgones de cultivos agrícolas en la Comunidad Valenciana. Bol. San. Veg. Plagas 30:317-326. [ Links ]
27. Nuestro clima. 2007. http://blog.nuestroclima.com. [Consulta: 4 de noviembre 2007] [ Links ]
28. Ortego, J. 1994. Curso sobre trampeo e identificación de pulgones vectores de virus de la papa. A.E.R. INTA, Malargüe, Mendoza, 88 pp. [ Links ]
29. Reed, W.; C. Cardona; S. Sithanantham and S. Lateef. 1986. Chickpea insect pest and their control. En: Saxena M. C. & K. B. Singh (eds.), The chickpea, CAB International-Wallingford, U. K., pp. 283-285. [ Links ]
30. Remaudiére, G. y M. Seco Fernández. 1990. Claves para ayudar al reconocimiento de alados de pulgones trampeados en la Región Mediterránea. Universidad de León, León, 205 pp. [ Links ]
31. Rossiter, M.; J. C. Schultz and T. Baldwin. 1988. Relationships among defoliation, red oak phenolics, and gypsy moth growth and reproduction. Ecology 69: 267-277. [ Links ]
32. Sharma, H.C.; C. L. Gowda; P. C. Stevenson; T. J. Ridsdill-Smith; S. L. Clement; G. V. Ranga Rao; J. Romeis; M. Miles and M. El Bouhssini. 2007. Host plant resistance and insect pest management in chickpea. En: Yadav, S. S. (ed.), Chickpea Breeding and Management, CAB International, Wallingford, pp. 520-537. [ Links ]
33. Singh, R. and N. Tripathi. 1987. Record of parasitoids from Tarai belt of eastern Uttar Pradesh. Journal of Aphidology 1: 89-92 [ Links ]
34. Starý, P. 1976. Aphid parasites (Hymenoptera, Aphidiidae) of the Mediterranean Area. Academia Nakladatelstvi Ceskoslovenske Akademie Ved. Praha, Czech Republic. [ Links ]
35. Tizado, E. J. y E. Núñez Pérez. 1991. Aportación al conocimiento en España de los parasitoides de pulgones de la subfamilia Aphidiinae (Hym. Braconidae). Bol. San. Veg. Plagas 17:545-554. [ Links ]
36. Van Der Maesen, L. J. 1972. Cicer L., A Monograph of the genus with special reference to the chickpea (Cicer arietinum L.), its ecology and cultivation. Comunication of the Agricultural University, Wageningen 72 (10), Netherland. [ Links ]
37. Zangerl, A. 1990. Furanocoumarin induction in wild parsnip: evidence for an induced defence against herbivores. Ecology 71: 1926-1932. [ Links ]
38. Zuazúa F.; J. E. Araya y M. A. Guerrero. 2003. Efectos letales de insecticidas sobre Aphidius ervi Haliday (Hymenoptera: Aphidiidae), parasitoide de Acyrthosiphon pisum (Harris) (Homoptera: Aphididae). Bol. San. Veg. Plagas 29: 299-307. [ Links ]
39. Zuparko, R. and D. Dahlsten. 1994. Host plant resistance and biological control for linden aphids. J. Arboric. 20 (5): 278-281. [ Links ]

