










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgriscientia
versión On-line ISSN 1668-298X
Agriscientia vol.39 no.1 Córdoba jun. 2022
Artículos
Avance genético y estabilidad de cultivares de maíz (Zea mays L.) obtenidos por el método de los híbridos crípticos
Genetic advance and stability of maize (Zea mays L.) cultivars obtained by the cryptic hybrid method.
F. H. Canteros 1
R. F. Caro 1
1 Canteros, F. H. (ORCID: 0000-0003-1584-0048): Instituto Nacional de Tecnología Agropecuaria, Agencia de Extensión Rural, Monteros, Ruta 38 Km 758. Tucumán, Argentina, CP 4142. Universidad Nacional de Tucumán, Facultad de Agronomía y Zootecnia, Finca El Manantial", Florentino Ameghino S/N. El Manantial, Tucumán, Argentina, CP 4105. Caro, R. F. (ORCID: 0000-0002-8495-3700): Universidad Nacional de Tucumán, Facultad de Agronomía y Zootecnia, Finca El Manantial", Florentino Ameghino S/N. El Manantial, Tucumán, Argentina, CP 4105.
Correspondencia a: canteros.francisco@inta.gob.ar
Fecha de recepción: 29/06/2021
fecha de aceptación: 03/11/2021
DOI: 10.31047/1668.298x.v39.n1.33696
RESUMEN
El cultivo de maíz es importante en el subtrópico argentino. Los programas de mejoramiento apuntan a lograr métodos eficientes. El objetivo de este trabajo fue evaluar el potencial del método de híbridos crípticos. En el Experimento 1 se determinaron las ganancias genéticas logradas (GsL) en híbridos de primer ciclo (HC1) para rendimiento (REND), prolificidad (PRO), peso de mil semillas (P1000), porcentaje quebrado de tallos (QUE%), porcentaje acame de plantas (AC%), intervalo antesis-emisión de estigmas (IAEE), altura inserción de espiga (AE) y altura de planta (AP). En el Experimento 2, se evaluó la estabilidad de los HC1 más los testigos, a través del gráfico GGE biplot. Para ello, se evaluaron los HC1 en cuatro ambientes: campo experimental de INTA en Leales, Tucumán (LE); en Finca El Manantial de la Facultad de Agronomía y Zootecnia, Universidad Nacional de Tucumán (MA); en Los Altos, Catamarca (LA); y en El Palomar, Santiago del Estero (PA). Los análisis se realizaron con Info-Gen. La GsL fue elevada para REND 1841,3 kg.ha-1 y QUE% (-18,28 %). En el Experimento 2, el híbrido HC116 sobresalió por REND y estabilidad. Esta metodología tiene un buen potencial para mejorar el REND y otros caracteres de interés.
Palabras clave: subtrópico, mejoramiento, ganancias genéticas.
SUMMARY
Maize cultivation is important in the Argentinian subtropics. Breeding programs aim at attaining efficient methods. The objective of this work was to evaluate the potential of the cryptic hybrids method. In Experiment 1, the genetic gains achieved (GsL) in first-cycle hybrids (HC1) were determined for yield (YD), prolificity (PRO), weight of thousand seeds (W1000), stalk broken (Sb%), stalk lodging (SL%), anthesis-silking interval (ASI), ear height (EH) and plant height (PH). In Experiment 2, the stability of the HC1 and control hybrids was evaluated through the GGE biplot graph. For this purpose, the HC1 were evaluated in four environments: in Leales, the experimental field of INTA,in Tucumán (LE), in Finca El Manantial of the Faculty of Agronomy and Zootechnics, National University of Tucumán (MA), in Los Altos, Catamarca (LA) and in El Palomar, Santiago del Estero (PA). The analyses were carried out with the statistical package Info-Gen. The GsL was high for YD, 1841.3 kg.ha-1, SB% (-15.28 %). In Experiment 2, the hybrid HC116achieved the best YD and stability. It was found that this methodology has good potential to improve the YD and other characters.
Keywords: subtropics, breeding, genetic gains.
INTRODUCCION
El objetivo final que persigue un programa de mejoramiento es el aumento de los rendimientos, lo que lleva a los mejoradores a replantearse continuamente los métodos utilizados y, sobre todo, la eficiencia de estos a la hora de evaluar los costos y los resultados obtenidos.
Los híbridos (F1) son la primera generación de un cruzamiento entre plantas de diferentes genotipos. La heterosis -o vigor híbrido- es el fenómeno por el cual se produce un incremento de tamaño, vigor o productividad de un híbrido, en comparación con el valor promedio, o media, de sus progenitores.
El objetivo principal de un programa de mejoramiento de maíz es la búsqueda de las mejores combinaciones de líneas con alta heterosis para la producción de híbridos. Para ello, los programas buscan tener, al menos, dos poblaciones que constituyan patrones o grupos heteróticos distintos que, al cruzarse, generen F1 que manifiesten vigor híbrido. El método tradicional se lleva a cabo a partir de dos variedades adaptadas de polinización libre, o de variedades sintéticas, que tengan heterosis al intercruzarlas.
El método de los híbridos crípticos fue propuesto por Villena (1965), Hallauer (1967), y Lonquist y Williams (1967), como una metodología para la obtención de híbridos superiores. Villena (1965) explica que esta metodología permite identificar, de forma temprana, las líneas cuyos genotipos en cruzas específicas expresan un alto grado de heterosis, y delinear métodos para su uso en un programa de mejoramiento genético.
Para seleccionar híbridos de esta forma, se necesitan poblaciones prolíficas, ya que el cruzamiento se realiza en simultáneo para obtener el híbrido, y la autofecundación para obtener líneas S1. Las líneas S1 tienen la mitad de la endogamia posible, y es todavía factible aprovechar la segregación que ocurre en cada una de ellas para llevar a cabo una selección intralíneas S1 mediante la misma técnica. Haber hecho la autofecundación en las dos plantas de cada cruza planta a planta, permite que los híbridos obtenidos de esa manera, puedan ser reproducidos posteriormente ya que, teóricamente, las frecuencias génicas de las poblaciones derivadas de las mazorcas autofecundadas deben ser similares (Marquéz Sánchez, 1988). Por lo tanto, el cruzamiento de plantas entre poblaciones derivadas de las mazorcas autofecundadas deberá ser similar al comportamiento de las cruzas originales planta a planta (Marquéz Sánchez, 1988).
Este método se basa en el aumento de la frecuencia de genes complementarios y la obtención de líneas con buena aptitud combinatoria específica (ACE) (Gomes Lopes et al., 2005; Botega Alves et al., 2012), que constituye el efecto más explotado en el mejoramiento para el rendimiento de maíz (Santos et al, 2007b) y permite la selección temprana de híbridos (Villena, 1965). Por otro lado, la prolificidad contribuye a la estabilidad de las poblaciones de maíz, y es una característica deseable en ambientes subóptimos (Prior y Russell, 1975), como los ambientes que caracterizan al subtrópico.
El objetivo de este trabajo fue evaluar el potencial del método de los híbridos crípticos, a través de la determinación del avance genético logrado de los híbridos desarrollados mediante el análisis del rendimiento, junto a otros caracteres de interés, y evaluar la estabilidad de los híbridos de primer ciclo de selección en cuatro ambientes.

Figura 1: Esquema de la técnica descripta para el método de los híbridos crípticos
MATERIALES Y MÉTODOS
Los grupos heteróticos denominados A y B se originaron de manera separada por el intercruzamiento de líneas parentales de diversos híbridos del Programa de Mejoramiento de Maíces Subtropicales (PMMS) del Instituto de Investigación Animal del Chaco Semiárido (IIACS), del Centro de Investigaciones Agropecuarias (CIAP) y del Instituto Nacional de Tecnología Agropecuaria (INTA). Previamente, estas líneas fueron convertidas a QPM (por el término en inglés Quality Protein Maize), a través del método de retrocruza. Se utilizaron, como padres donantes, las líneas de CIMMYT, CML 161 y CML 165 para cada grupo, respectivamente. Luego, a cada uno de los grupos, se incorporó resistencia a las imidazolinonas (RI). La selección para RI, se realizó usando un herbicida en doble dosis a base de imidazolinona, sobre las plantas de los dos grupos heteróticos.
Las líneas de cada grupo convertidas a RI, se usaron como macho balanceado (mezcla de cantidades iguales de semillas de cada una de las líneas). En la campaña 2013-2014, las semillas de F1 se sembraron en lotes de intercruzamientos aislados. En la campaña 2014-2015, las semillas F2 (semillas de plantas F1) se sembraron en dos lotes aislados. En cada uno de los lotes, se seleccionaron plantas de dos espigas usando la metodología de selección masal estratificada, descripta por Gardner (1961).
Las poblaciones formadas, denominadas variedad sintética A ciclo 0 (SYN0 A) y variedad sintética B ciclo 0 (SYN0B), se sembraron en la campaña 2015-2016 en surcos alternados de ambas poblaciones y se aplicó el método de los híbridos crípticos, autofecundando y cruzando las plantas selectas (Figura 1). A partir de ello, se formaron los híbridos de ciclo 0 (HC0), las S1 del grupo heterótico A (S1A) y S1 del grupo heterótico B (S1B). Las S1 fueron identificadas y guardadas en cámara de frío.
La realización de este trabajo se llevó a cabo mediante dos experimentos. El Experimento 1, en el IIACS (CIAP, INTA). Los materiales seleccionados para la evaluación de los híbridos del ciclo 0 (HC0) fueron producto del cruzamiento de plantas SiA individuales y prolíficas (Figura 1) de las SYN0A y SYN0B. Se obtuvieron 42 HC0 y 84 S1 (Tabla 1), que fueron sembrados en un ensayo comparativo de rendimiento (ECR) el 21 de diciembre de 2016 (campaña 2016-2017). El primer año, se seleccionaron las 15 familias de mejor desempeño, a partir de los resultados de la prueba de límite de significancia estadística de Fisher (LSD). Esto representó una presión de selección del 35,71 %. En el ciclo de invierno de 2017 en invernáculo, los 22 híbridos de ciclo 1 (HC1) (Tabla 2) se formaron con las familias selectas.

Tabla 1: Códigos e híbridos HC0 utilizados en el ensayo, campaña 2016-2017, en IIACS – INTA
En el Experimento 1, para el estudio de la ganancia genética lograda (GsL), se compararon los resultados de las variables bajo estudio de los HC0 (campaña 2016-2017) y HC1 (campañas 2017-2018), sin considerar los testigos. Los ECR se llevaron a cabo en el campo experimental del IIACS, del CIAP, INTA (27° 03' S, 64° 15' O; 330 m s. n. m.), cuyas características edafoclimáticas se muestran en la Tabla 3.
El avance genético logrado (GsL) fue determinado mediante la fórmula: GsL = (xHC1 - xHC0), que es la diferencia de media de los híbridos HC1 de la temporada 2017-2018 (XHC1) y los híbridos HC0 de la temporada 2016-2017 (XHC0), a partir de lo propuesto por Vencovsky y Barriga (1992). La ganancia lograda en porcentaje (GsL%) fue calculada mediante la fórmula: (GsL / XHC0 Gral ).100. Los datos para el cálculo de GsL y GsL% se muestran en la Tabla 4.
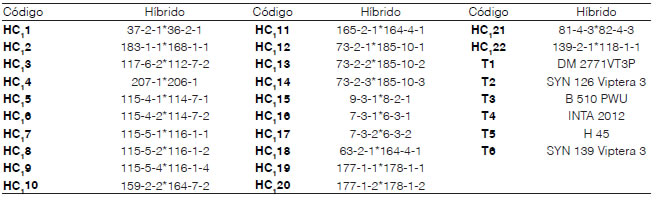
Tabla 2: Códigos e híbridos HC1 utilizados en el ensayo, campaña 2017-18 en IIACS – INTA
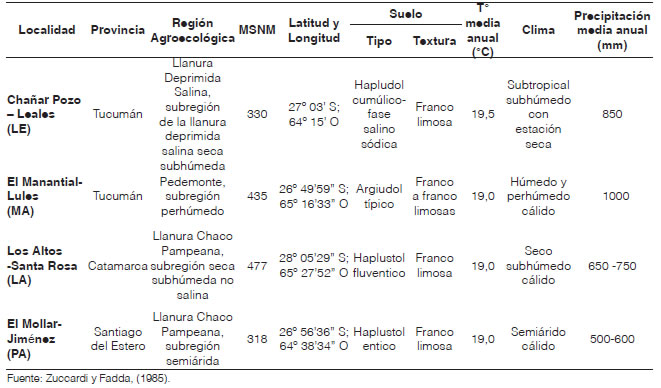
Tabla 3: Localidad, provincia, región agroecológica, msnm, latitud y longitud, suelo, temperatura, clima y precipitación de los sitios donde se realizaron los ensayos de los híbridos H
En el Experimento 2 se analizó la estabilidad de comportamiento de los HC1 en diferentes localidades. Para ello, se realizaron ECR de los 22 HC1 más los seis testigos (Tabla 2), llevados a cabo en la campaña 2017-2018, que fueron sembrados el 2 de enero de 2018 en el campo experimental del IIACS (CIAP-INTA) en Leales, provincia de Tucumán (27° 03' S, 64° 15' O; 330 m s. n. m.) (LE); el 4 de enero de 2018 en Finca El Manantial FAZ-UNT en Lules, provincia de Tucumán (26° 49' 59 S, 65° 16' 33 O; 435 m s. n. m.) (MA); el 10 de enero de 2018 en Los Altos, Santa Rosa, provincia de Catamarca (28° 05' 29 S, 65° 27' 52 O; 477 m s. n. m.) (LA); y el 16 de enero de 2018 en El Mollar Jiménez, provincia de Santiago del Estero (26° 56' 36 S, 64° 38' 34 O; 318 m s. n. m.) (PA). Las características edafoclimáticas de las localidades se describen en la Tabla 3. La parcela experimental para LE y MA fue de un surco de 5 m de longitud, espaciados a 0,70 m (3,5 m2), con 20 plantas distanciadas a 0,25 m, con una densidad de plantas ideal de 5,7 pl.m2. En tanto, en LA y PA, la parcela experimental fue de un surco de 5 m de longitud, espaciados a 0,52 m (2,6 m2), con 15 plantas distanciadas a 0,33 m, lo que da una densidad ideal de 5,7 pl.m2. La siembra, en todos los casos, se realizó con bastón sembrador, 3 semillas por golpe y luego, se raleó a una planta en estado fenológico V2-V3 (Ritchie y Hanway, 1982). Los ECR se dispusieron en un diseño de bloque completo al azar (DBCA) con dos repeticiones. Se fertilizó con 50 kg.ha-1 de fosfato di amónico (18-46-0) en la siembra, y con 200 kg.ha-1 de urea (46 % N) en el estado fenológico de V6 (Ritchie y Hanway, 1982). Se usaron como testigos seis híbridos comerciales, que se repitieron en las dos campañas. Cuatro de ellos son híbridos simples de semilleros comerciales, recomendados para el noroeste argentino (NOA), DM 2771 VT3P, OGM (Don Mario Semillas), SYN126 Viptera 3 y SYN 139 Viptera 3, tropicales, OGM's (Syngenta Argentina), B510 PWU, OGM (Brevant Semillas), y dos del PMMS, H45 e INTA 2012, tropicales convencionales, sin manipulación genética.
Las malezas fueron controladas con aplicaciones de herbicida total (2 l.ha-1 de glifosato 48 %) previo a la siembra y un herbicida pre-emergente (3 l.ha-1 de atrazina), y carpidas manuales durante el resto del ciclo del cultivo. Los insectos (cogollero) fueron controlados según los niveles de infestación (dos aplicaciones de deltametrina en dosis de 75 cm3.ha-1). Los ECR fueron conducidos en secano.

Tabla 4: Valores del promedio general de los híbridos y de ganancia genética
En el Experimento 1 se determinó:
-Rendimiento ajustado a 14,5 % de humedad del grano (REND). Se obtuvo cosechando a mano la parcela y luego, se procedió a la trilla de las espigas con máquina estacionaria. Se pesó en balanza electrónica (Kretz, origen Argentina). Se midió dos veces la humedad de los granos de cada parcela con humedímetro (DIKEY-John MULTI-GRAIN, origen EE. UU.), se promediaron los valores y se llevaron todas las muestras a 14,5 % de humedad, por último, se transformaron a kg.ha-1.
-Prolificidad (PRO): el número de espigas por planta se determinó en cosecha, dividiendo el número espigas cosechadas por el número de plantas de la parcela útil.
-El peso de 1000 semillas (P1000), se determinó a partir de alícuotas de 500 granos, extraídas de cada una de las muestras cosechadas, secadas hasta peso constante en estufa a 65 °C y pesadas en balanza electrónica (modelo Adventure, marca OHAUS, origen EE. UU.), con una capacidad máxima de 3100 gr, precisión 0,01 gr.
-Porcentaje de quebrado de tallos (QUE%): se contó el número de plantas que presentaban quebrado de tallo por debajo de la espiga principal. Los datos se tomaron dos semanas antes de la cosecha y fue evaluada en porcentaje, con respecto al total de plantas de la parcela.
-Porcentaje de acame de plantas (AC%): se contó el número de plantas que presentaban una inclinación mayor de 30° con respecto a la vertical por parcela. Los datos se tomaron dos semanas antes de la cosecha y fue evaluada en porcentaje, con respecto al total de plantas de la parcela.
-Intervalo antesis, emisión de estigmas (IAEE): en 10 plantas marcadas de cada parcela, se determinó la cantidad de días desde la siembra a 50 % de plantas, con panojas emitiendo polen (VT), y espigas con estigmas visibles (R1), de acuerdo a la clave de Ritchie y Hanway (1982). Luego, se sacó la diferencia de días mediante la fórmula IAEE = R1-VT.
-Altura de inserción de espiga (AE): en cinco plantas de cada parcela, se midió la distancia en m, desde la base de la planta hasta el nudo de inserción de la espiga principal y se sacó el promedio.
-Altura de planta (AP): en cinco plantas de cada parcela, se midió la distancia en m, desde la base de la planta hasta el extremo superior de la panoja y se sacó el promedio.
Debido a que los datos de REND, PRO y P1000 de los HC1 de la campaña 2017-2018, fueron superiores en un 31,51 %, 14,78 % y 8,36 % respecto a los de la campaña 2016-2017, para la determinación del avance genético logrado (GsL), estas variables fueron corregidas sobre la base de la diferencia de los valores promedios de los testigos en ambas campañas. Esto es similar a lo reportado por Hallauer (1967).
En el Experimento 2, solo se determinó REND, como se describe arriba.
Los análisis de ANOVA, LSD de Fisher y análisis de estabilidad y comportamiento de los híbridos a través del gráfico GGE biplot (Figura 2), se hicieron con el paquete estadístico Info-Gen (Balzarini y Di Rienzo, 2004).
RESULTADOS Y DISCUSIÓN
A partir de los datos del Experimento 1, la GsL para REND fue de 1841,30 kg.ha-1 y la GsL% 25,48 % (Tabla 4), valor mucho mayor a la reportada por otros autores que usaron la misma metodología, como es el caso de Coors (1999), quien reportó valores de entre 4,5 % y 6,2 %.ciclo-1. Por su parte, Raposo et al. (2004) y Reis et al. (2009) reportaron valores de 7,9 % y 5,7 %, respectivamente. Eyherabide y Hallauer (1991) reportaron ganancias por ciclo de 6,5 % en poblaciones templadas. Keeratinijakal y Lamkey (1993a) reportaron 6,5 %.ciclo-1, mientras que Santos et al. (2007b) obtuvieron un promedio de 7,2 %.ciclo-1.
Para la PRO, la GsL fue de 0,08 y GsL% de 6,5 % (Tabla 4). Este valor fue más bajo que el reportado por Souza Jr. y Pinto (2000) que fue de 11,8 %, pero mayor que el 2,62 % reportado por Santos et al. (2007a). Como se muestra en la Tabla 4, el promedio para este carácter en los HC0 es de 1,23 espigas.planta-1, que es alto y se encuentra fijado en las poblaciones que forman los grupos heteróticos, que dieron origen a los híbridos evaluados en este trabajo.
En P1000 la GsL fue de -40,13 y GsL% -13,03 (Tabla 4), esto representa una disminución de 40,13 gr, y es contrario a lo reportado por da Cunha et al. (2012), quienes reportaron un incremento de 0,58.
La GsL del QUE% fue de -15,28 (Tabla 4), es decir, muy por debajo del -1,64 % reportado por Schnicker y Lamkey (1993), del -1,04 % obtenido por Keeratinijakal y Lamkey (1993b), del -7,6 % por Menz Rademacher et al. (1999), del -1,65 % en condiciones de sequía y -1,17 % en condiciones de secano, informado por Badu-Apraku et al. (2018). Esta gran diferencia en los resultados, con respecto a los reportados por otros autores, puede deberse a la presión de selección aplicada en los HC0, que pasaron al próximo ciclo (HC1) sobre este carácter.

Figura 2: GGE biplot.Identificación de los mejores híbridos en cada localidad.Los puntos amarillos representan localidades, los azules los híbridos HC1.
La GsL para AC%, fue de +5,66 % (Tabla 4). Esta difiere de lo reportado por Schnicker y Lamkey (1993), -0,23 %, y Menz Radermacher et al. (1999), -11,0 %, también de lo reportado por Badu-Apraku et al. (2018), en condiciones de sequía y en condiciones de secano, que encontraron valores de -1,05 y -1.09 %, respectivamente.
El IAEE tuvo una tuvo una GsL -0,22 días y GsL % de -36,07 % (Tabla4). El valor encontrado es superior al reportado por Badu-Apraku et al. (2018) para dos condiciones ambientales diferentes, que fue de -0,52 % en condiciones de sequía y -0,45 % en secano. Este resultado es positivo ya que la reducción de este carácter le confiere mayor tolerancia al estrés por sequía a los cultivares (Bolaños y Edmeades, 1996)
La AE tuvo una GsLde -0,13 cm y GsL % de -9,42 %(Tabla 4). La disminución de AE fue mucho mayor que la reportada por Santos et al. (2007a), que fue de -3,17 %.
Para AP la GsL-0,03 cm y GsL% -1,06 % (Tabla 4). Este valor de GsL % fue similar al reportado por Santos et al. (2007a), de -1,81 %.
El ANOVA para REND, conjunto de los híbridos HC1, en las cuatro localidades del Experimento 2 se puede ver en la Tabla 5. Se observa que hubo significancia entre los G (Híbridos), E (Localidades), pero no así en la interacción genotipo ambiente (GEI). Esto es similar a lo reportado por Oyervides et al. (1993), Biasutti et al. (2004) y Alcocer et al. (2005), quienes trabajando con pasturas en nuestra región tampoco encontraron GEI.
Al no existir interacción GEI, puede inferirse que las diferencias entre los híbridos se mantienen en los diferentes ambientes, o que dichos ambientes no son discriminantes para estas variables. Es por ello que se realizó un análisis de LSD (Tabla 6), como fue sugerido por Mariotti (1994).
El análisis LSD conjunto de todos los híbridos en las cuatro localidades (Tabla 6) muestra al híbrido HC116 como el de mayor REND, superando a los testigos T4 y T6. En tanto, HC112, HC113, HC15, HC11, HC17 superan al testigo T3 y HC13, HC121 y HC118, HC14, HC115, HC16 superan a los testigos T2 y T5. Todos estos se encuentran por encima del promedio, por lo cual 12 de los 22 HC0 superan, al menos, a dos testigos y se encuentran por encima del promedio.
Del ANOVA (Tabla 5), se desprende que los E constituyen la fuente de variación más importante, en relación a la contribución de los G y GEI sobre la variabilidad total. Por ello, Cornelius et al. (1996), Crossa y Cornelius (1997) y Crossa, Cornelius y Yan (2002), proponen el modelo regresión por sitio (SREG).
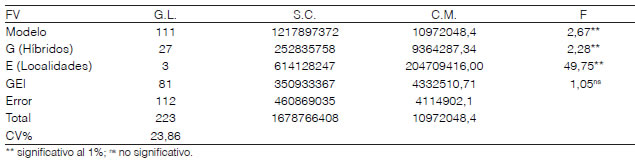
Tabla 5: Análisis de la varianza para REND en cuatro localidades

Tabla 6: Prueba de LSD, Alfa 0,05 para REND de los híbridos HC1
En el gráfico GGE biplot (Figura 2), se observa que los componentes principales CP1 explican (47,1 %) y CP2 (25,2 %) y, en conjunto, explican 72,3 % de la variabilidad total G y G + E.
Según la propuesta de interpretación de Yan y Hunt (2002), se puede observar en la Figura 2 que se diferencian tres mega ambientes formados por MA, LE, y el tercer ambiente formado por las localidades de LA y PA. El ambiente MA resultó el más discriminante.
En tanto el híbrido HC116, se encuentra en el vértice del polígono, y presenta el mejor valor en el eje CP1. Además de un valor cercano a cero en el eje CP2, lo que indica que este híbrido es el más productivo, con una buena adaptación y estabilidad para REND en los ambientes bajo evaluación. Por otro lado, se encontraron híbridos sitio-específicos para REND: HC11, HC116, HC13 y HC16 para MA; HC113 y HCJ8 para LE; HC17, HC115, HC15 y HC121 para LA; HC15, HC112 y HC17, para PA.
CONCLUSIONES
En cuanto a las GsL para REND, fue relativamente alta, comparada con lo reportado por otros autores que usaron la misma metodología. Esto puede deberse a que estamos en el primer ciclo de selección y es de esperar que la GsL para REND disminuya con el avance de los ciclos. Además, se trabajó con poblaciones mejoradas a priori para REND. Por otro lado, se cumplió con la finalidad de esta metodología, que es la selección temprana de las familias con heterosis.
En cuanto a PRO, la GsL fue baja. Esto puede deberse al hecho de que las poblaciones que dan origen a las líneas progenitoras de híbridos tienen este carácter fijado, ya que para aplicar esta metodología son necesarias las plantas prolíficas.
El QUE% tuvo un decrecimiento de la GsL, lo que representa una tendencia positiva, en tanto para AC% tuvo un leve aumento. Esto último, aunque indeseable, no es tan relevante, ya que la espiga no cae al suelo y, por lo tanto, no se genera una pérdida de REND.
Se consiguió una disminución de AE y AP, que es lo que se busca en los híbridos modernos.
Pudo identificarse el híbrido HC116, de amplia adaptación y estabilidad de REND, además de híbridos con adaptación sitio-específico. También, cinco híbridos, HC112, HC113, HC15, HC11 y HC17, tuvieron rendimientos superiores a cuatro de los seis testigos en el análisis conjunto de REND. Por otra parte, se identificaron tres mega ambientes, MA, LE, y un tercero formado por las localidades de LA y PA.
BIBLIOGRAFÍA
Alcocer, G., Pérez, P. G., García Posse, F. R. y Devani, M. R. (2005). Determinación de estabilidad de la producción varietal de Chloris gayana Kunth en Tucumán y zonas de influencia. Revista Industrial y Agrícola de Tucumán, 82(1-2), 55-58. http://www.scielo.org.ar/scielo.php?pid = S1851-30182005000100007&script=sci_arttext&tlng=pt
Balzarini, M. y Di Rienzo, J. (2004). Info-Gen (versión 2004) [Software]. Córdoba, Argentina: Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba. https://www.info-gen.com.ar/
Badu-Apraku, B., Talabi, A. O., Ifie, B. E., Chabi, Y. C., Obeng-Antwi, K., Haruna, A. y Asiedu, R. (2018). Gains in grain yield of extra-early maize during three breeding periods under drought and rainfed conditions. Crop Science, 58(6), 2399-2412. https:// doi.org/10.2135/cropsci2018.03.0168
Biasutti, C. A., Peiretti, D. A., Nazar, M. C. y Alemanno, G. A. (2004). Respuesta a la selección masal por prolificidad en maíz en diferentes ambientes. AgriS-cientia, 21(2). https://doi.org/10.31047/1668.298x.v21. n2.2666
Bolaños, J. y Edmeades, G. O. (1996). The importance of the anthesis-silking interval in breeding for drught tolerance in tropical maize. Field Crops Research, 48(1), 65-80. https://doi:10.1016/0378-4290(96)00036-6
Botega Alves, N., Pereira Nascimento J., Pereira C.H., Van Rooijen, M., Rui, M. C. y Souza, J. C. (26 - 30 de agosto de 2012). Avaliagao de híbridos crípticos de milho. Documento presentado en XXIX Congresso Nacional de Milho e Sorgo. Águas de Liondóia, Sao Paulo, Brasil.
Coors, J. G. (1999). Selection methodology and heterosis. EnJames G. Coors y Shivaji Pandey (Eds.), The Genetics and Exploitation of Heterosis In Crops (225-245). The American Society of Agronomy, Inc. y Crop Science Society of America, Inc. https://doi. org/10.2134/1999.geneticsandexploitation.c21
Cornelius, P. L., Crossa, J. y Seyedsadr, M. S. (1996). Statistical tests and estimators of multiplicative models for genotype-by-environment interaction. Genotype by Environment Interaction (No. 631.523 KAN. CIMMYT.).
Crossa, J. y Cornelius, P. L. (1997). Sites regression and shifted multiplicative model clustering of cultivar trial sites under heterogeneity of error variances. Crop Science, 37(2), 406-415. https://doi.org/10.2135/crop sci1997.0011183X003700020017x Crossa, J., Cornelius, P. L. y Yan, W. (2002). Biplots of linear bilinear models for studying crossover genotypex environment interaction. Crop Science, 42(2), 619633. https://doi.org/10.2135/cropsci2002.6190 da Cunha, K. S., Pereira, M. G., Gongalves, L. S. A., Berilli, A. P. C. G., de Oliveira, E. D., Ramos, H. C. C. y do Amaral Júnior, A. T. (2012). Full-sib reciprocal recurrent selection in the maize populations cimmyt and piranao. Genetics and Molecular Research, 11(3), 3398-3408. http://dx.doi.org/10.4238/2012.September.25.8
Eyherabide G. H. y Hallauer A. R. (1991). Reciprocal full-sib recurrent selection in maize: I direct and indirect responses. Crop Science, 31(4), 952-959. https://doi. org/10.2135/cropsci1991.0011183X003100040023x Gardner C. O. (1961). An evaluation of effects of mass selection and seed irradiation with thermal neutrons on yield of corn. Crop Science, 1(4), 241-245. https://doi. org/10.2135/cropsci1961.0011183X000100040004x Gomes Lopes, M. T., Soriano Viana, J. M. y Lopes, R. (2005). Capacidade produtiva de híbridos de famílias endogamicas de milho (Zea mays L.) obtido pelo método dos híbridos crípticos. Revista Ceres, 52(303), 613-632. Universidad Federal de Vigosa. Minas Gerais. Brasil. https://locus.ufv. br//handle/123456789/13082 Hallauer, A. R. (1967). Development of single cross hybrids from two eared maize populations 1. Crop Science, 7(3), 192-195. https://doi.org/10.2135/crops ci1967.0011183X000700030007x Keeratinijakal, V. y Lamkey, K. R. (1993a). Responses to reciprocal recurrent selection in BSS and BCSB1 maize populations. Crop Science, 33(1), 73-77. https://doi.org/10.2135/cropsci1993.0011183x003300 010012x
Keeratinijakal, V. y Lamkey, K. R.(1993b). Genetic Effects Associated with reciprocal Recurrent Selection in BSSS and BSCB1 Maize Populations. Crop Science, 33(1), 78-82. https://doi.org/10.2135/cropsci1993.001 1183x003300010013x
Lonnquist, J. H. y Williams, N. E. (1967). Development of maize hybrids through selection among full-sib families 1. Crop Science, 7(4), 369-370. https://doi. org/10.2135/cropsci1967.0011183X000700040028x Mariotti, J. (1994). La interacción genotipo-ambiente, su significado e importancia en el mejoramiento genético y en la ecuación de cultivares. Serie Monográfica N° 1. INTA-CRTS.
Marquéz Sánchez, F. (1988). Genotecnia Vegetal. Métodos, Teoría, Resultados. Tomo II. AGT Editor.
Menz Rademacher, M. A., Hallauer, A. R. y Russell, W. A. (1999). Comparative response of two reciprocal recurrent selection methods in BS21 and BS22 maize populations. Crop Science, 39(1), 89-97. https://doi. org/10.2135/cropsci1999.0011183X00390001014x
Oyervides, A., Mariaca, J., De Leon, H. y Reyes, M. (1993). Estimación de parámetros genéticos en una población de maíz tropical. Agronomía mesoamericana. 4(1), 30-35. https://d¡alnet.un¡r¡oja. es/servlet/articulo?codigo=5542800
Prior, C. L. y Russell, W. A. (1975). Yield performance of nonproliflc and proliflc maize hybrids at six plant densities 1. Crop Science, 15(4), 482-486. https://doi. org/10.2135/cropsci1975.0011183X001500040010x
Raposo, F V., Patto Ramalho, M. A. P y Ribeiro, P H. E. (2004). Alterations in heterosis of maize populations derived from single-cross hybrids after reciprocal recurrent selection. Crop Breeding an Applied Biotechnology 4(1), 74-80. http://www.alice.cnptia. embrapa.br/alice/handle/doc/487790
Reis, M. C. D., Souza, J. C. D., Ramalho, M. A. P, Guedes, F L. y Santos, P H. A. D. (2009). Progresso genético com a selegáo recorrente recíproca para híbridos interpopulacionais de milho. Pesquisa Agropecuária Brasileira, 44, 1667-1672. https://doi.org/10.1590/S0100-204X2009001200016
Ritchie, S. W. y Hanway, J. J. (1982).How a Corn Plant Develops (Special Report N° 48). lowa State University of Science and Technology, Cooperative Extension Service.
Santos, F S., Amaral Junior, A. T., Freitas Junior, S. P, Rangel, R. M. y Pereira, M. G. (2007a). Predigáo de ganhos genético por índices de selegáo na populagáo de milho-pipoca UNB-2U sob selegáo recorrente. Bragantia, 66(3), 389-396. https://doi.org/10.1590/ S0006-87052007000300004
Santos, M. F., Cámara, T M. M., Moro, G. V., Costa, E. F. N. y De Souza, C. L. (2007b). Responses to selection and changes in combining ability after three cycles of a modified reciprocal recurrent selection in maize. Euphytica, 157(1), 185-194. https://doi. org/10.1007/s10681-007-9410-x
Schnicker, B. J. y Lamkey, K. R. (1993). Interpopulation genetic variance after reciprocal recurrent selection in BSSS and BSCB1 maize populations. Crop Science, 33(1), 90-95. https://doi.org/10.2135/cropsci1993.001 1183X003300010016x
Souza Jr., C. L. y Pinto, R. M. C. (2000). Response to a short-term reciprocal recurrent selection procedure in maize. Maydica, 45(1), 21-28.
Vencovsky, R. y Barriga, P (1992). Genética biométrica aplicada ao fitomelhoramento. Ribeirao Preto: Sociedade Brasileira de Genética, p. 469.
Villena, W. (1965). Selección de líneas para habilidad combinatoria general y específica en el primer ciclo de selección recurrente recíproca en poblaciones de Eto Blanco y Colima 14. En Proc. 11 Reunión Anual PCCMCA, 31-34. Ciudad de Panamá, Panamá.
Yan, W. y Hunt, L. A. (2002). Biplot analysis of diallel data. Crop science, 42(1), 21-30, 21-30. https://doi. org/10.2135/cropsci2002.2100a
Zuccardi, R. B. y Fadda, G. S. (1985). Bosquejo agroecológico de la provincia de Tucumán. Miscelánean° 86. Universidad Nacional de Tucumán, Facultad de Agronomía y Zootecnia: San Miguel de Tucumán, Argentina.

