










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkAgriscientia
versión On-line ISSN 1668-298X
Agriscientia vol.39 no.1 Córdoba jun. 2022
Artículos
Efecto de Adara-M2624 y otros siete portainjertos sobre daños y defectos de rutos de dos variedades de cereza (Prunus avium L.)
Effect of Adara-M2624 and other se-ven rootstocks on fruit damages and defects of two sweet cherry varieties.
E. Tersoglio 1 *
N. Setien 1
1 Tersoglio, E. (ORCID: 0000-0002-0869-4858) y Setien, N. (ORCID: 0000-0002-5870-3601): Instituto Nacional de Tecnología Agropecuaria (INTA), Dirección San Martín 3853, Luján de Cuyo, Mendoza, Argentina. CC3 CP 5507.
* Correspondencia a: tersoglio.eduardo@inta.gob.ar
DOI: 10.31047/1668.298x.v39.n1.28816
Fecha de recepción: 29/05/2020
fecha de aceptación: 12/11/2021
RESUMEN
Los portainjertos modifican el desempeño del árbol, adaptándose a diferentes condiciones edafo-climáticas. El diseño experimental fue de bloques completos al azar, con seis repeticiones. El porcentaje de daño se midió a través de los siguientes componentes: fruto partido, ausencia de pedúnculo y daño mecánico. El porcentaje de defecto se midió con los siguientes componentes: fruto con espolón, frutos dobles y sutura profunda. La adición de daño y de defecto constituyó la pérdida total. Las condiciones climáticas, variedad y portainjerto influyeron sobre todas las variables. Los portainjertos influyen sobre las pérdidas de las variedades Bing y Celeste. Los portainjertos que registraron la menor pérdida y, simultáneamente, el menor daño y defecto fueron SL 64, MxM 60 y Weiroot 13; mientras que CAB6P y Adara-Mariana 2624 redujeron los defectos, pero incrementaron los daños. Tanto GiSeLa®5 como GiSeLa®6 redujeron los daños, aunque GiSeLa®5 incrementó los defectos. Los portainjertos SL 64, MxM 60, Weirrot 13 y GiSeLa®6 redujeron agrietado, daño mecánico y sutura; CAB6P y Adara-Mariana 2624 redujeron sutura y espolón, pero incrementaron agrietado y daño mecánico; GiSeLa®5 redujo los tres tipos de daños e incrementó todos los tipos de defectos. Una adecuada selección del portainjerto disminuye pérdidas.
Palabras clave: cerezo, fruto partido, daños mecánicos, sutura profunda, espolón.
SUMMARY
Rootstocks modify tree performance by adapting to different soil and climatic conditions. The experimental design was a randomized complete block design, with six replications. The percentage of damage was measured by the following components: cracked fruit, stemless fruit and mechanical damage. The percentage of defect was measured by the following components: spur fruit, double fruit and deep suture. The addition of damage and defect constituted the total loss. Climatic conditions, variety and rootstock ¡nfluenced all variables. Rootstocks influenced the losses of Bing and Celeste varieties. The rootstocks that recorded the lowest loss and simultaneously, the lowest damage and defect were SL 64, MxM 60 and Weiroot 13, whereas CAB6P and Adara-Mariana 2624 reduced defects but increased damage. Both GiSeLa®5 and GiSeLa®6 reduced damage, although GiSeLa®5 increased defects. Rootstocks SL 64, MxM 60, Weirrot 13 and GiSeLa®6 reduced cracking, mechanical damage and suture; CAB6P and Adara-Mariana 2624 reduced suture and spur but increased cracking and mechanical damage; GiSeLa®5 reduced all three types of damage and increased all types of defects. Proper rootstock selection reduces losses.
Keywords: sweet cherry tree, cracked fruit, mechanical damage, deep suture, spur fruit.
INTRODUCCION
La calidad del fruto es el factor que más impulsa la decisión del comprador; incluso define el lugar de la adquisición y se vincula estrechamente con la variedad. La cantidad adquirida por el consumidor se incrementa en un 24 % cuando la calidad de la fruta es alta y ejerce una gran influencia sobre la fidelización del cliente (Wyman et al., 2019). La adquisición es impulsada principalmente, por la apariencia (Wyman et al., 2019) compuesta por los atributos de calidad: tamaño, color (Crisosto et al., 2003; Chiang et al., 2018) y estado del pedúnculo (Neilsen et al., 2010). Sin embargo, la satisfacción del comprador en el instante del consumo está gobernada por otros atributos como firmeza, contenido de sólidos solubles y su relación con la acidez (Crisosto et al., 2003). Según López-Ortega et al. (2016) los portainjertos influyen sobre esos atributos.
El portainjerto también influye en el comportamiento del árbol, mejorando su adaptación a diferentes condiciones ambientales, modificando la fisiología (Steiner et al., 2015), la absorción de nutrientes, la arquitectura del dosel, la ontogenia de órganos florales (Ljubojevic et al., 2016), la producción, la eficiencia productiva, la tolerancia a hipoxia (Gainza et al., 2015) y la eficiencia de cosecha (Ampatzidis y Whiting, 2013). También modifica ciertos atributos de calidad como el tamaño (Sansavini y Lugli, 2014; Gainza et al., 2015; López-Ortega et al., 2016), firmeza, contenido de sólidos solubles, acidez e índice de madurez (López-Ortega et al., 2016).
Sin embargo, la apariencia de la fruta comercializable se debe no sólo a la presencia de tales atributos sino, además, a la ausencia de daños y defectos. Por lo tanto, los frutos que no satisfacen el estándar de calidad deben eliminarse mediante selección en el empaque. Ello incrementa los costos de empaque, la huella de carbono (Rana et al., 2019) y las pérdidas económicas debido a la reducción de la cantidad comercializada.
En cereza, los daños se clasifican como fruto partido (FP), daños mecánicos (DM) y ausencia de pedúnculo (AP) (InnovaChile, 2010). El daño FP se debe a un colapso localizado de la hipodermis, consecuencia de la absorción directa y pasiva de agua a través de las micro-fracturas de la membrana cuticular. Esto provoca un aumento del tamaño de las células epidérmicas que reduce la adhesión célula-célula. Posteriormente se produce la hipertrofia, debido a la fuerza impulsora del bajo potencial osmótico del fruto y, luego, la ruptura de células de la hipodermis que provoca el colapso de la epidermis y la formación de macro-grietas (Winkler, Brüggenwirth et al., 2016; Correia et al., 2018; Schumann y Knoche, 2020). Tal fenómeno induce principalmente grietas en la cavidad peduncular y en la cicatriz pistilar. Sin embargo, FP también es causado por exceso de agua de riego, ya que ocasiona un elevado flujo de agua hacia el fruto a través del sistema vascular vía simplasto - apoplasto que suele inducir grietas laterales (Measham et al., 2010; Measham et al., 2014). Para su control, se propone el uso de coberturas impermeables, que son confiables aunque costosas (Simon, 2006); el uso de variedades tolerantes, que es una alternativa económica, pero aun así se registran daños (Sansavini y Lugli, 2014); y la técnica de aplicaciones de soluciones de Calcio, que sin embargo es de baja reproducibilidad (Winkler y Knoche, 2019).
El Dm posee orígenes diversos. Entre estos, podemos mencionar cosecha inapropiada que ocasiona heridas, ausencia de pedúnculo, daños por impacto o por compresión que forman lesiones, como picaduras o pitting y magulladuras o bruising, ambas muy frecuentes durante la cosecha y postcosecha (Hussein et al., 2020), así como heridas causadas por pájaros (Anderson et al., 2013). Menos comunes son las lesiones superficiales producidas por patógenos (Lucero et al., 2015) y por raspado, rayado o russet que es una lesión cicatricial de la cutícula y de la epidermis debido a causas climáticas, virosis o daños por insectos (Martínez y Vilardell, 200).
La AP se debe, principalmente, a malas prácticas de cosecha, aunque también se vincula a variedades con menor fuerza de retención del pedúnculo (Wirch et al., 2009; Neilsen et al., 2010), portainjerto (Kappel y Lang, 2008), carga frutal (Bound et al., 2013), exceso de madurez por cosecha tardía y mayor dificultad de cosecha de doseles grandes (Ampatzidis y Whiting, 2013).
Los defectos se clasifican como: sutura profunda (SP) (Southwick and Uyemoto, 1999), fruto con espolón (EP) (Engin and Ünal, 2008a) y fruto doble (FD) (Engin et al., 2009). Se forman durante la post-cosecha del año anterior, a partir de la iniciación floral y durante todo el desarrollo de la yema floral. El inicio es, aproximadamente, 21 días después de finalizada la cosecha (Engin et al., 2009). El desarrollo de los verticilos florales es centrípeto, la formación de la yema floral inicia a fines de junio en el hemisferio norte (HN) y el desarrollo del gineceo se inicia tarde en agosto (HN) (Engin y Ünal, 2008a). Tanto FD como EP se originan tempranamente por una división extra del primordio pistilar, formando ovarios dobles que luego se fusionan y crecen (Engin et al., 2009). La formación de EP ocurre cuando uno de ellos es fecundado y el restante, de menor tamaño, es retenido (Engin y Ünal, 2008a) y la de FD cuando ambos pistilos son de tamaño similar y fecundados simultáneamente (Engin et al., 2009). Mientras que SP se forma por una cicatrización parcial del carpelo, tardíamente, alrededor del 23 de agosto (HN) (Engin y Ünal, 2008a; Engin et al., 2009). La formación de defectos es consecuencia de altas temperaturas durante la inducción y diferenciación floral (Southwick y Uyemoto, 1999; Engin y Ünal, 2008a; Engin et al., 2009; S0nstebya y Heideb, 2019). Los defectos aumentan cuando la temperatura de post-cosecha supera los 30 °C (Beppu y Kataoka, 1999).
Los antecedentes indican que portainjerto y variedad varían su desempeño según la zona, lo que destaca la importancia de la selección local (Ljubojevic et al., 2016; Sansavini and Lugli, 2014), especialmente cuando se usan portainjertos enanizantes (Fajt et al., 2014). La combinación portainjerto - variedad ideal es la que permite obtener alta producción de fruta sin daño o defecto, de tamaño comercial, de bajo costo de selección y de alta eficiencia en la gestión de cosecha.
El objetivo de este trabajo fue estudiar la influencia de ocho portainjertos sobre los daños, defectos y sobre sus componentes en dos variedades de cerezas, en las condiciones locales.
MATERIALES Y MÉTODOS
Condiciones del ensayo
El ensayo se realizó en la Estación Experimental Agropecuaria INTA, Mendoza, Argentina (33° 00' 13 S, 68° 51' 34 O, a 920 m s.n.m.), cuyo clima es continental árido y desértico, con una temperatura media anual de 17 °C y gran amplitud térmica. Los veranos son cálidos, con una precipitación media de 190 mm. La oferta promedio de frío invernal es de 1106 unidades de frío (Tersoglio y Naranjo, 2007). La profundidad del suelo, la textura y las características edáficas fueron descritas en Tersoglio y Setien (2016). El ensayo se inició en 2004, tuvo un marco de plantación de 3 m entre plantas y 5 m entre hileras, cultivado bajo malla antigranizo. El sistema de conducción fue de ejes múltiples con tres ejes por planta y 6 a 7 laterales cada uno. La lámina de riego se aplicó por goteo con una reposición fija de 7,6 mm cada vez que el potencial matriz del suelo alcanzó los 30-40 kPa.
Materiales vegetales
Las pérdidas se midieron durante la cosecha del sexto al octavo año después de la injertación (ADI). Los portainjertos fueron: Prunus mahaleb L. selección SL 64; los híbridos inter-específicos (Prunus mahaleb x Prunus avium) selecciones MxM 60 y MxM 14, Prunus cerasus L. selecciones CAB 6P y Weiroot 13 (W 13), los híbridos interespecíficos (Prunus cerasus x Prunus canescens) selecciones GiSeLa®5 (G 5) y GiSeLa®6 (G 6) y la combinación de Mariana 2624 (Prunus cerasifera Ehrh. x Prunus munsoniana W. Wight & Hedrick) y el filtro inter-compatible Prunus cerasifera var. Mirobalan selección Adara (Adara-M2624) (Tersoglio y Setien, 2016).
Las variedades utilizadas fueron: Bing clon OB260 de la Universidad de Oregon (EE.UU.), dosel de vigor medio-alto, auto-incompatible grupo III, tamaño de fruto, maduración y productividad intermedia, susceptible a la partidura; y Celeste de vigor medio, auto-fértil grupo IX, floración media, maduración temprana y muy susceptible a la partidura.
Cosecha y muestreo
La cosecha se inició cuando un tercio de la producción alcanzó el grado 3 de la tabla de colores de cosecha de cereza de la EEA Mendoza (Instituto Nacional de Tecnología Agropecuaria [INTA], 2007), equivalente al código 4 de la tabla de colores del Centre Technique Interprofessionel des Fruits et Legumes [CTIFL], (2005). El muestreo fue sistemático, se extrajo el 10 % correspondiente a cada recipiente, aproximadamente 0,4 kg. Consecuentemente, el tamaño de muestra osciló entre 0 kg y 6 kg según producción de cada unidad experimental (UE). La cosecha se hizo en una o en dos pasadas dependiendo de la concentración de la madurez. Aunque se pesó toda la producción, solo se evaluó la primera pasada. De cada muestra, se extrajo al azar la sub-muestra, cuyo tamaño osciló entre 1,0 kg y 3,0 kg.
Variables analizadas
Desde el sexto hasta el octavo ADI, los frutos se clasificaron visualmente según presencia-ausencia de cada componente de daño o defecto. Tanto el porcentaje de daño (Daño) como el de defecto (Def) se calcularon como la correspondiente suma de FP, Dm y AP y de FD, EP y Sp respectivamente. La pérdida porcentual (Per) se calculó como la suma del porcentaje de daño y defecto.
Anualmente, los frutos de cada sub-muestra fueron clasificados según cada componente de daño y de defecto, luego se pesaron y se calculó el porcentaje. Se expresó como g de fruta de cada componente de daño o de defecto por cada 100 g de fruta cosechada.
Variables complementarias
Para mejorar la comprensión de las relaciones entre daños o defectos según condiciones ambientales, se utilizaron dos variables climáticas: precipitación (Pp) durante los 21 días previos a la cosecha (Measham et al., 2009) que se expresó en mm; y grados acumulados de crecimiento horario, con un umbral térmico de 30 °C (Beppu y Kataoka, 1999; Tersoglio y Naranjo, 2007) registrado en el período entre 21 días después de cosecha y el 28 de febrero (Engin y Ünal, 2008a), que se expresó como GDH > 30 °C.
Las variables volumen de dosel (VD) y altura de dosel (H) se midieron según Tersoglio y Setien (2016). Además, se calculó eficiencia productiva según volumen de dosel (EfV) y peso medio de fruto (PM). Se expresaron en m3, m, g.m-3 y g, respectivamente.
El diseño fue de bloques completos al azar, con seis repeticiones, con cuatro plantas por UE. Para el análisis de la varianza, se usó el programa estadístico InfoStat (Di Rienzo et al., 2018). La prueba de comparación múltiple de medias del ANAVA fue Scott & Knott (S&K) y el nivel de significancia fue p < 0,05. Los coeficientes de correlación utilizados fueron el de Pearson y Spearman según corresponda. Los modelos de regresión fueron seleccionados con TableCurve 2D (Wilkinson, 2002). Para medir el aporte de información de variables de clasificación se utilizó el método de comparación de regresiones anidadas (Mendenhall and Sincich, 1995).
RESULTADOS Y DISCUSIÓN Análisis de pérdida porcentual (Per)
La Per media fue 40,5 ± 22,9 %, que correspondió a fruta cosechada dañada y/o defectuosa. Grant et al. (2001) consideraron razonable una Per media del 20 %, lo que indica que los valores locales observados, sin considerar la falta de tamaño, duplican el valor propuesto.
La variabilidad se explicó por el modelo (R2Aj = 0,70; p < 0,0001), mediante los efectos bloque (p = 0,0451), ADI (p < 0,0001), variedad (p < 0,0001), portainjerto (p = 0,0263) e interacción variedad-portainjerto (p = 0,0073).
Dentro de las pérdidas está el daño por agrietado, que es de gran importancia en la mayoría de las zonas productoras (Correia et al., 2018). Mientras que Southwick et al. (1991) mencionaron la relevancia de los defectos sutura profunda, espolón y fruto doble en la producción de cerezas del valle central de California, EE. UU.
En el séptimo ADI, la Per se incrementó el 20,3 % respecto de la media, seguido por el sexto y octavo ADI con valores inferiores que la media (Tabla 1). La variedad Celeste registró un incremento del 14,4 %, respecto de Bing (Tabla 1). En tal sentido, Simón (2006) y Michailidis et al. (2020) citaron que existen diferencias de susceptibilidad a Fp entre variedades; mientras que Engin et al. (2009) mencionaron que existen variedades más susceptibles a experimentar determinados defectos que otras.

Tabla 1: Efecto del ADI, portainjerto, variedad e interacción sobre Per, Daño, Def, FP, DM, AP, SP y EP-FD
Los portainjertos SL 64, MxM 60, CAB 6P y W 13 registraron reducciones sobre la P media del 4,4 %, 3,4 %, 1,6 % y 1,8 %, respectivamente; mientras que MxM 14, G 5, G 6 o Adara-M2624 midieron incrementos sobre la media del 3,8 %, 2,9 %, 3,0 % y 1,6 %, respectivamente (Tabla 1). Según Correia et al. (2018), portainjerto es un factor que puede modificar la absorción de agua e influir en el agrietado de los frutos de un mismo cultivar.
Análisis de porcentaje de daño (Daño)
La media de D _ fue 26,4 ± 25,7 %, que correspondió al 65,2 % de las pérdidas totales.
Asoció con Pp (r = 0,87; p < 0,0001), con variedad (r = 0,25; p < 0,0001), pero solo Bing lo hizo con VD (r = 0,28; p = 0,0007). La variabilidad se explicó por el modelo (R2Aj = 0,85; p < 0,0001) mediante los efectos bloque (p = 0,0455), ADI (p < 0,0001), portainjerto (p = 0,0002), variedad (p < 0,0001) e interacción ADI-variedad (p < 0,0001), y ADI-portainjerto-variedad (p = 0,0007).
El séptimo ADI incrementó D - un 31 % respecto de la media. Esto como consecuencia de la mayor Pp que indujo mayor FP (Correia et al., 2018), pero también por la mayor cantidad de lesiones causadas por hongos (Lucero et al., 2015) (datos no mostrados). Mientras que el octavo y el sexto ADI, con bajas PP, registraron reducciones del 20 % y del 11 %, respectivamente (Tabla 1).
La variedad Celeste incrementó el D _ un 9,5 % ano respecto de Bing (Tabla 1). Sansavini y Lugli (2008) mencionaron que FP es el dano más frecuente y que existen diferencias de susceptibilidad entre variedades. También, los danos ocasionados por aves suelen ser altos en variedades tempranas, como Celeste, ya que estas disponen de menos fuentes alternativas de alimento (Mertz, 2016). Además, la fuerza de retención del pedúnculo varía con la variedad (Neilsen et al., 2010). La Figura 1 muestra un comportamiento interanual similar de ambas variedades; aunque el de Celeste registró mayor Dano que el de Bing excepto en el sexto ADI, que no se diferenció.
Los portainjertos SL 64, MxM 60, W 13, G 5 o G 6 registraron menor D _ que el resto con reducciones ano sobre la media del 3,5 %, 1,8 %, 1,7 %, 4,3 % y 0,1 %, respectivamente. Mientras que MxM 14, CAB 6P o Adara-M2624 registraron aumentos sobre la media del 1,8 %, 4,5 % y 4,8 %, respectivamente (Tabla 1). La selección del portainjerto es un factor que controla la absorción de agua que puede afectar D _ en los frutos (Simon et al., 2004; Correia et al.,

Figura 1: Interacción ADI-variedad sobre D0f y Dano. Test de S&K (a= 0,05) 2018). La interacción portainjerto-variedad no tuvo significancia (p = 0,1115).

Figura 2: Efecto interacción ADI-portainjerto-variedad según el Dano medio y efecto portainjerto-variedad del séptimo ADI según, Dano medio varietal. Test de S&K (a= 0,05)

Figura 3: Relación PP - FP según variedad
Análisis de porcentaje de fruto partido (FP)
La media fue 14,5 ± 18 %. Correia et al. (2018) y Michailidis et al. (2020) mencionaron que FP es un dano observado en la mayoría de las regiones productoras, con enormes pérdidas comerciales. Asoció con EfV (r = -0,18; p = 0,0024), semejante a lo mencionado por Measham et al. (2012) y por Bound et al. (2013). También lo hizo con PM (r = 0,22; p = 0,0002). Según Simon (2006) y Correia et al. (2018), a menor carga frutal le corresponde mayor PM que se asocia con mayor FP. También asoció con variedad (r = 0,36; p < 0,0001) y con DM (r = 0,53; p < 0,0001), pero no con AP (p = 0,0869). Contrariamente, asoció con magnitud negativa, con los tres defectos: SP (r = -0,40; p < 0,0001), Ep (r = -0,17; p = 0,0036) y Fd (r = -0,28; p < 0,0001). Asoció con P (r=0,74; p <0,0001) y se relacionó con un modelo lineal cuadrático (R2 = 0,62; p < 0,0001) en el que ambas variedades compartieron el mismo modelo. Sin embargo, la relación de cada variedad tuvo sus propios parámetros (p < 0,005), indicando que el comportamiento de Fp dependió de la variedad, tal como fue señalado por Measham et al. (2009).
La Figura 3 muestra que, para alcanzar la media de FP la variedad Bing requirió una Pp de 65 mm durante el período de cosecha, mientras que Celeste lo hizo sólo con 38 mm. Sin embargo, este resultado es contrario a lo citado por Simon (2006), indicando la importancia de la selección local de los materiales genéticos mejor adaptados, tal como lo mencionaron Ljubojevic et al. (2016).
La variabilidad de FP fue explicada por el modelo (R2Aj = 0,77; p < 0,0001) mediante bloque (p = 0,0372), ADI (p < 0,0001), variedad (p < 0,0001), portainjerto (p = 0,0235) e interacciones variedad-portainjerto (p = 0,0083), ADI-variedad (p < 0,0001) y ADI-variedad-portainjerto (p = 0,0003).
El FP del séptimo ADI se incrementó el 18,0 % respecto de la media, seguido por el octavo ADI con valores cercanos a la media y el del sexto ADI con valores inferiores a la media. La variedad Celeste registró un incremento del 11,0 °% respecto de Bing (Tabla 1). Correia et al. (2018) sugirieron que variedades de fruto de forma arriñonada, como Celeste, poseen cavidad peduncular más profunda, mantienen la piel húmeda por más tiempo después de una lluvia, absorben más agua y consecuentemente presentan mayor incidencia de Fp. Por lo tanto, cualquier estrategia de control debe considerar el efecto variedad (Measham et al., 2009).
La Figura 4 muestra que, en el sexto ADI, la variedad Bing con 18,2 mm de Pp y la variedad Celeste con 14,8 mm tuvieron FP cercano a cero. En el séptimo ADI, Bing con 86,4 mm de PP y Celeste con 83,8 mm tuvieron incrementos sobre la media del 10,6 °% y del 25,4 °%, respectivamente. Mientras que en el octavo ADI, Bing con 8,6 mm de PP redujo FP el 12,8 °% y Celeste con 24,4 mm lo incrementó el 3,8 °%, respecto de la media. Sin embargo, Simon (2006) resaltó que Celeste fue menos susceptible a FP que Bing, confirmando que es un daño difícil de predecir a campo, debido a numerosos factores que lo afectan. Entre ellos se citan tamaño del fruto y EfV (Measham et al., 2013), elasticidad de la epidermis, presencia de micro-fracturas en la membrana cuticular (Correia et al., 2018), contenido de sólidos solubles, firmeza (Simon, 2006), PP y estado fenológico (Measham et

Figura 4: Efecto interacción ADI-variedad y Pp sobre Fp. Test de S&K (a= 0,05) al., 2009), lámina y frecuencia de riego (Measham et al., 2014) y duración de la lluvia (Correia et al., 2018; Winkler et al., 2020), entre otros.
Los portainjertos CAB 6P y Adara-M2624 incrementaron FP en 3,8 °% y 2,1 °%, respectivamente; el resto de la serie no diferenció de la media (Tabla 1). Según Simon et al. (2004) y Correia et al. (2018), la selección del portainjerto es un factor que altera la absorción de agua e influye en el agrietamiento de los frutos. Michailidis et al. (2020) afirmaron que FP es consecuencia de la absorción directa de agua por la epidermis y de la absorción de agua por el sistema vascular del árbol. Además, Simón (2006) y Correia et al. (2018) señalaron que, para una misma variedad, los frutos grandes son más susceptibles que los pequeños. En coincidencia, el mayor PM registrado sobre CAB6P y Adara-M2624 podrían explicar el mayor FP (datos no mostrados).
El FP de la variedad Bing no diferenció portainjerto, mostrando la importancia del efecto simple variedad. Sin embargo, la variedad Celeste, más susceptible, diferenció portainjertos con valores superiores a los de Bing. Sobre SL 64, MxM 60 o W 13 registró valores cercanos a la media; mientras que sobre MxM 14, G 5, G 6 o Adara-M2624 se incrementó el 7,4 %, 6,6 %, 5,2 % y 5,9 % sobre la media, respectivamente. El mayor incremento se observó sobre CAB 6P con 13,9 % sobre la media (Tabla 1). La evidencia indica que FP diferenció portainjerto solo cuando la variedad fue muy susceptible. Measham et al. (2010 y 2014) indicaron que el flujo neto de savia xilemática del pedúnculo es alto después de una lluvia y es impulsado, principalmente, por el potencial de turgencia causado por la absorción de agua osmótica, desde el simplasto hacia el apoplasto del fruto. Por su parte, la tensión por transpiración del fruto tuvo poca importancia, aunque la funcionalidad xilemática del pedúnculo disminuyó hacia la madurez, reduciendo la tensión que finalmente se anula y aísla el fruto (Winkler, Peschel et al., 2016), e indica que la influencia del portainjerto se reduce hacia la madurez. Measham et al. (2010) encontraron que riegos excesivos se asocian a una expansión celular y consecuente aumento del volumen del fruto, que induce grietas grandes y profundas sobre las mejillas; mientras que las apicales (cicatriz pistilar y cavidad peduncular) se asocian al contacto con agua libre de lluvia. Por ello, Measham et al. (2014) consideraron que la selección del portainjerto constituye una estrategia de interés. Por otro lado, Winkler et al. (2020) señalaron que FP es función principalmente de la duración de la humedad y del área mojada del fruto, siendo la cavidad peduncular (efecto cavidad) y cicatriz pistilar (efecto gota) las zonas más frecuentemente afectadas.
La interacción ADI-portainjerto-variedad de la Figura 5 muestra que Bing sobre todos los portainjertos y que Celeste sobre SL 64, MxM 60, W 13, G 5 o G 6 registraron FP menor o igual a la media, en dos de los tres ADI.
En el séptimo ADI (Figura 5), la media de FP de Bing fue 25,1 % y sobre SL 64, MxM 14 o G 5 registró valores menores que la media varietal, con disminuciones del 4,1 %, 3,5 % y 5,4 %, respectivamente. La media de Celeste fue 39,9 %, pero sobre SL 64, MxM 60, W 13 o Adara-M2624 se redujo 9,9 %, 6,5 %, 5,5 % y 12,6 %, sobre la media varietal.
La información indica que variedades susceptibles a FP mejoran su desempeño sobre los portainjertos SL 64, MxM 60 o W 13.
Análisis de porcentaje daño mecánico (DM)
La media de DM fue 7,4 ± 8,8 %. Asoció con FP (r = 0,53; p < 0,0001), con AP (r = 0,19; p = 0,0066) y con variedad (r = 0,35; p < 0,0001). Aunque con menor magnitud que FP, también asoció con Pp (r = 0,69; p < 0,0001) y se relacionó con un modelo lineal cuadrático (R2 = 0,52; p < 0,0001), indicando que PP durante la maduración, además de causar FP también incrementa DM. La Figura 6 muestra diferencias varietales, ya que para alcanzar el DM medio, Bing requirió 61 mm; mientras que Celeste lo hizo con solo 41 mm, indicando la mayor susceptibilidad de esta última.
No asoció con EfV (p = 0,9907) y con PM (p = 0,8692) y con VD (p = 0,8949). Consecuentemente, los datos sugieren que una mayor EfV, PM y menor VD no se asocian con mayor DM. Según Gainza et al. (2015), la reducción de los costos de cosecha se logra reduciendo el VD. Según Koumanov et al. (2018) y Ampatzidis y Whiting (2013) también se logra mediante la selección de una adecuada arquitectura de dosel.
La variabilidad de DM se explicó con el modelo (R2Aj = 0,74; p < 0,0001) mediante los efectos ADI (p <0,0001), portainjerto (p = 0,0001), variedad

Figura 5: Efecto interacción ADI-portainjerto-variedad sobre Fp. Test de S&K (a= 0,05)

Figura 6: Relación Pp-DM según variedad
En las condiciones del séptimo ADI, se incrementó DM un 8,6 % sobre la media; mientras que en el sexto y octavo ADI disminuyó 3,8 % y 4,8 %, respectivamente. Esto indica que una alta PP durante la maduración incrementa simultáneamente FP y DM.
Al igual que FP la variedad Celeste registró mayor DM que Bing con un incremento del 3,9 % (Tabla 1). El mayor DM de Celeste pudo deberse a su pedúnculo particularmente corto, que dificulta la cosecha e induce tomar los frutos con los dedos, situación que incrementaría daño por magulladuras, tal como lo expuso Hussein et al. (2020). La interacción ADI-variedad (p = 0,1095) y variedad-portainjerto (p = 0,4298) no tuvieron significancia. El portainjerto Adara-M2624 incrementó DM 3,8 % sobre la media; el resto no se diferenció (Tabla 1). El portainjerto Adara-M2624 fue más precoz que el resto (datos no mostrados), consistente con lo observado por López-Ortega et al. (2016). Ello sugiere mayor frecuencia de frutos muy maduros y, consecuentemente, mayor frecuencia de DM por magulladuras (Hussein et al., 2020). Además, la mayor precocidad de cosecha indujo mayor daño por aves, tal como señala Mertz (2016).
La Figura 7 muestra el comportamiento de las combinaciones variedad-portainjerto, durante el séptimo ADI. La media de Bing fue 13,5 % y la de Celeste fue 18,3 %. Cuando Bing se injertó sobre MxM 60, MxM 14, W 13 o G 5 y Celeste sobre MxM 14, CAB 6P o G 5 se registraron valores de DM menores a las correspondientes medias varietales. Tanto Bing como Celeste sobre Adara-M2624 registraron incrementos del 13,2 % y 7,2 % respecto de las correspondientes medias varietales. En relación con CAB6P, ambas variedades no diferenciaron, pero mostraron un comportamiento inverso, siendo Bing la que registró el mayor DM.
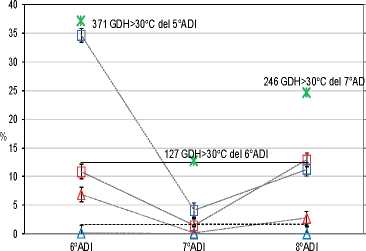
Figura 7. Efecto interacción variedad-portainjerto sobre el DM del séptimo ADI. Test de S&K (a= 0,05)
Análisis de porcentaje de ausencia de pedúnculo (AP)
La AP registró una media de 4,7 ± 6,7 %. Asoció con VD (r = 0,33; p < 0,0001) y con H (r = 0,42; p < 0,0001), consistente con lo mencionado por Koumanov et al. (2018), quienes sugirieron que el tamaño del dosel influye sobre la AP. Además, asoció con Pp (r = 0,44; p < 0,0001) y con variedad (r = 0,48; p < 0,0001), tal como lo reportaron Wirch et al. (2009) y Neilsen et al. (2010).
La variabilidad de AP se explicó con el modelo (R2Aj = 0,77; P < 0,0001) por los efectos bloque (p = 0,001), ADI (P < 0,0001), portainjerto (p < 0,0001), variedad (P < 0,0001) e interacción portainjerto-variedad (p = 0,0026), ADI-variedad (p < 0,0001), bloque-variedad (p = 0,0052).
El comportamiento interanual de AP fue similar al de FP y DM. La mayor incidencia fue en el séptimo ADI, con un incremento del 4,3 % sobre la media (Tabla 1). Neilsen et al. (2010) señalaron que la reducción de la fuerza de retención del pedúnculo y el consecuente aumento de AP se debe, entre otros, a la mayor madurez del fruto. Por lo tanto, el incremento del séptimo ADI podría deberse al retraso de la cosecha, consecuencia de las condiciones climáticas.
La variedad Bing registró un incremento del
5.3 % respecto de Celeste (Tabla 1). La menor AP correspondió a G 5, seguido por SL 64, G 6 y Adara-M2624 con reducciones de la media del
3.4 % y del 1,1 %, 0,3 % y 1,2 %, respectivamente. Los de mayor incidencia fueron MxM 60, MxM 14, CAB 6P o W 13, con incrementos sobre la media 0,6 %, 3,0 %, 1,0 % y 1,2 %, respectivamente (Tabla 1). Así, se confirma lo observado por Kappel y Lang (2008), quienes reportaron la influencia de portainjerto sobre la retención del pedúnculo. La AP de la variedad Celeste no diferenció portainjerto, registrando valores menores que la media; y, por lo tanto, se considera una variedad tolerante a este defecto. Mientras que Bing solo sobre G 5 redujo la AP el 2,5 %; no obstante, sobre MxM 14 la incrementó el 8,4 % respecto de la media. Luego, Bing sobre MxM 60, CAB 6P o W 13 incrementó el 3,2 %, 4,6 % y 4,2 % sobre la media, respectivamente. El resto tuvo valores cercanos a la media (Tabla 1). Nuevamente la evidencia indica que AP diferenció portainjerto solo cuando la variedad fue susceptible. Las variedades susceptibles a AP mejoran sobre G 5, aunque también lo hacen sobre G 6, SL 64 o Adara-M2624.
Análisis de porcentaje de defecto (Def)
La media de Def fue 14,1 ± 16,9 %, que correspondió al 34,8 % de las pérdidas totales. Southwick et al. (1991) indicaron que, en algunos años, los defectos pueden alcanzar hasta el 80 % de los frutos. El desarrollo defectuoso de los frutos se produce durante la formación de las yemas florales debido a que son muy sensibles a las altas temperaturas (Villar et al., 2020). En tales condiciones las variedades pueden registrar un determinado defecto o varios (Usenik et al., 2005; Engin et al., 2009).
Asoció con GDH > 30 °C (r = 0,60; p < 0,0001) y con VD (r = -0,36; p < 0,0001). Steiner et al. (2015) encontraron que las yemas florales de doseles pequeños, sobre portainjertos enanizantes, están más expuestas a la luz y experimentan mayor temperatura que las de los doseles grandes. Este incremento de la temperatura induce la formación de frutos defectuosos (Beppu y Kataoka, 1999; Engin et al., 2009; S0nstebya y Heideb, 2019).
También asoció con EfV (r = 0,31; p < 0,001) y con PM (r = -0,32; p < 0,0001). Los frutos más pequeños pueden experimentar, más frecuentemente, una cicatrización carpelar incompleta (Engin y Ünal, 2008a; Engin et al., 2009). Engin y Ünal (2008b) encontraron que la proporción de gineceos defectuosos disminuye en el período que va de yema floral otoñal hasta la fecundación en primavera, indicando que hay una reducción natural de la frecuencia de ovarios deformes. Pero, en condiciones muy favorables, aumenta la fecundación de gineceos deformes. Y, consecuentemente, la formación de frutos defectuosos, incrementando la EfV y reduciendo el que (p < 0,0001), ADI (p < 0,0001), portainjerto (p < 0,0001), variedad (p < 0,0001), variedad-portainjerto (p = 0,0088), ADI-variedad (p < 0,0001) y ADI-portainjerto (p = 0,0029).
El sexto ADI registró un incremento del 13,8 % sobre la media, seguido por el octavo y el séptimo con reducciones del 3,1 % y 10,7 % sobre la media, respectivamente (Tabla 1). La variedad Celeste incrementó Def un 4,9 % respecto de Bing (Tabla 1). Además, la Figura 1 muestra que el comportamiento interanual de Def difiere con la variedad. Tanto Bing como Celeste diferenciaron todos los ADI. En el sexto y octavo ADI, Bing superó la media, y en el séptimo ADI fue menor.
La variedad Celeste superó la media en el sexto, mientras que en el séptimo y octavo fue menor. Southwick y Uyemoto (1999), Engin y Ünal (2008a) y Engin et al. (2009) coincidieron en que los defectos resultan de la interacción entre el control genético y las condiciones ambientales en la postcosecha de año anterior.
Los portainjertos SL 64, MxM 60, CAB 6P, W 13 o Adara-M2624 redujeron Def en un 1,0 %, 1,6 %, 6.0 %, 0,2 % y 3,3 % de la media, respectivamente. Mientras que MxM 14 o G 6 registraron el 2.0 % y el 2,9 % de incremento sobre la media, respectivamente, siendo el de G 5 el mayor con 7, 2 % (Tabla 1).
La variedad Bing sobre MxM 14 o G 5 incrementó Def en un 1,4 % y 1,7 % sobre la media, respectivamente; mientras que los portainjertos restantes lo redujeron entre 1,1 % y 8,7 %. La variedad Celeste sobre SL 64, MxM 14, W 13 o Adara-M2624 registró valores cercanos a la media, pero sobre G5 o G6 registró incrementos sobre la media del 12,6 % y 6,9 %, respectivamente (Tabla 1). Ambas variedades sobre MxM 60 o CAB 6P formaron un grupo homogéneo, registrando el menor Def e indicando que ambos portainjertos presentan estabilidad al cambio varietal, ya que no diferenciaron entre variedades (Tabla 1).
Southwick y Uyemoto (1999) observaron que el estrés hídrico durante la post-cosecha del año anterior indujo Def. Steiner et al. (2015) encontraron que la regulación de la conductancia estomática difiere entre portainjertos, incluso con un adecuado suministro hídrico. En cerezos, la máxima conductancia estomática se alcanza con temperaturas foliares de 30 °C a 40 °C, pero entre 40 °C y 45 °C disminuye rápidamente. Este proceso de regulación es más rápido en portainjertos enanizantes que en vigorosos, indicando que los restrictivos suministran menos agua a las hojas, por su sistema hidráulico más débil. En tal sentido, Olmstead et al. (2006) observaron que la unión en la zona injerto-portainjerto del enanizante G5 suele ser defectuosa, aumentando la tensión hidráulica y reduciendo el crecimiento. En consecuencia, las yemas de doseles pequeños tienen mayor exposición a la radiación solar y alcanzan más rápidamente el rango crítico de temperatura (Steiner et al., 2015). En tal sentido, el portainjerto más enanizante de la serie, el G5, indujo el mayor Def medio. Sin embargo, el semi-enanizante W13 mostró valores bajos (Tabla 1). Además, la interacción ADI-portainjerto (Figura 8) mostró que G 5 registró menor Def que la media sólo en uno de los tres ADI, mientras que W 13 lo hizo en dos de los tres ADI. Los datos sugieren que, en zonas cálidas, existen pocos portainjertos que reducen el
VD y además disminuyen Def. Entre los portainjertos que registraron el menor nivel de Per sólo SL 64, MxM 60 y W 13 mostraron simultáneamente bajo D , y bajo D - con una relación D - -D , de 1,8; 2,0 y 1,8 veces, respectivamente (Tabla 1). Mientras que CAB 6P y Adara-M2624 registraron alto Daño y bajo Def, con una relación de 3,8 y 2,9 veces. Los G 5 y G 6 midieron el menor D - , pero G 5 registró el

Figura 8: Efecto interacción ADI-portainjerto sobre Der Test de S&K (a = 0,05)
Análisis de porcentaje de sutura profunda (SP)
La SP media fue 12,5 ± 16,2 % con datos que oscilaron entre 0 % y 85 %, indicando que, además de ser el defecto de mayor incidencia, en condiciones desfavorables puede alcanzar gran importancia. Asoció con VD (r = -0,33; p < 0,0001), con H (r = -0,41; p < 0,0001), con ADI (r = 0,38; p < 0,0001), con GDH > 30 °C del año precedente (r = 0,54; p < 0,0001), con EfV (r = 0,33; p < 0,0001) y con PM (r = -0,32; p < 0,0001).
La SP relacionó con GDH > 30 °C y VD, mediante un modelo simple lineal (Raj2 = 0,52; p < 0,0001) a través de la ecuación
Sp =A + (b * GDH > 30° C2) + (c * VDct°,5), cuyos parámetros fueron A = 12,018; b = 1,6278E-04; c = -2,9205716. Este modelo indica que, al duplicar GDH > 30 °C de 150 a 300 unidades, manteniendo fijo el VDct en 16,8 m3 que corresponde al máximo volumen disponible (Tersoglio y Setien, 2016), la SP aumentó de 3,7 °% a 14,7 °%, un incremento del 11,0 °%. Pero, al reducir el VD a la mitad, manteniendo fija la cantidad de calor, en 300 GDH > 30 °C, la SP aumentó de 14,7 °% a 18,2 °%, un incremento de 3,5 °%. Esto indica que SP es más sensible al incremento de GDH > 30 °C que a la reducción de VD.
Asoció con Pp (r = -0,43; p < 0,0001), la magnitud negativa puede deberse a que frutos con SP en presencia de agua libre tienden a desarrollar FP con mayor facilidad y frecuencia. Esto se debe a la ausencia de una correcta cicatrización carpelar que interrumpe la formación de la epidermis y de la membrana cuticular (Engin et al., 2009). En casos extremos la sutura puede extenderse hasta el endocarpio (Southwick et al., 1991). La falta de membrana cuticular permite que las células del parénquima de la pulpa, estructuralmente más débiles y con un potencial osmótico más negativo, contacten y absorban agua más rápidamente. Ello ocasiona un rápido agrietamiento siguiendo el modelo de la cremallera a lo largo de la sutura (Winkler, Peschel et al., 2016). Por el contrario, Winkler et al., (2020) observaron que en frutos sanos hay baja frecuencia de FP sobre la sutura, ya que la membrana cuticular existe y las gotas escurren rápidamente. Por ello, en condiciones de clima lluvioso, frutos con el defecto SP tienden a desarrollar el daño FP.
El modelo (R2Aj = 0,77; p < 0,0001) explicó la variabilidad de SP por los efectos bloque (p < 0,0001), ADI (p < 0,0001), portainjerto (p = 0,0001), variedad (p < 0,0001) e interacción portainjerto-variedad (p < 0,0001), ADI-variedad (p < 0,0001), ADI-portainjerto-variedad (p = 0,007), bloque-portainjerto (p = 0,0214).
El sexto ADI registró la mayor SP, con un incremento del 12,1 % respecto de la media; mientras que en el séptimo y en el octavo se redujo el 9,2 °% y el 2,9 °%, respectivamente (Tabla 1). Ambas variedades fueron susceptibles a experimentar SP; sin embargo, Celeste duplicó a Bing, con un incremento del 8,2 °% (Tabla 1). Southwick y Uyemoto (1999) reportaron el origen genético de estos defectos. La Figura 9 muestra que SP es influido por los GDH > 30 °C del año anterior según la variedad (Beppu y Kataoka, 1999). Con una oferta de calor durante el quinto ADI de 371 GDH > 30 °C, la variedad Celeste registró en el sexto ADI un incremento del 22 % sobre la media que correspondió a 2,8 veces la media; mientras que Bing registró una reducción sólo del 1,7 %, que correspondió a 0,9 veces la media. Con los 127 GDH > 30 °C registrados durante el sexto ADI, la SP del séptimo ADI de Bing y de Celeste no se diferenciaron y ambas tuvieron valores menores que la media, con reducciones del 11,0 % y 8,4 %, respectivamente. Con los 246 GDH > 30 °C registrados durante el séptimo ADI, la SP del octavo ADI de ambas variedades registraron valores que no diferenciaron de la media. La información indica que, con 127 o 246 GDH > 30 °C, no diferenciaron variedad y tuvieron baja a media incidencia de Sp. Sin embargo, entre 246 GDH > 30 °C y 371 GDH > 30 °C la variedad Celeste triplicó la media, mientras que Bing no la superó. Esto sugiere que Celeste experimentó SP cuando la oferta de calor excedió cierto umbral ubicado entre 246 y 371 GDH > 30 °C (Engin et al., 2009).

Figura 9: Interacción ADI-variedad sobre Sp y Ep-Fd según GDH > 30 °C del año precedente. Test de S&K (a= 0,05)
Los portainjertos MxM 14 o G 5 incrementaron SP 2,6 % y 5,6 % sobre la media, respectivamente; el resto formó un grupo homogéneo con menor SP (Tabla 1). La variedad Bing sobre G 6 o Adara-M2624 redujo SP el 8,9 % y 7,9 %, respectivamente; el resto tuvo valores cercanos a la media, mientras que la variedad Celeste sobre G 5 registró la mayor SP incrementando un 14,3 % sobre la media (Tabla 1). Ambas variedades sobre MxM 60 o CAB 6P formaron un grupo homogéneo con valores cercanos a la media, indicando que ambos portainjertos son estables al cambio varietal (Tabla 1). La evidencia indicó que SP diferenció combinaciones portainjerto-variedad, debido a que ambas variedades fueron susceptibles.
La relación SP-GDH > 30 °C ajustó con un modelo lineal cúbico (R2 = 0,31; p < 0,0001). Pero, al incorporar la información de los grupos homogéneos de variedad-portainjerto, se obtuvo un modelo lineal cúbico (Raj2 = 0,57; p < 0,0001) que mejoró la calidad de ajuste (p < 0,005). Los cuatro grupos se relacionaron con el mismo modelo, pero cada uno tuvo sus propios parámetros. La Figura 10 muestra que Bing sobre G6 o Adara-M2624 no alcanzó la SP media, aún con una alta oferta de calor de 371 GDH > 30 °C. Mientras que Bing sobre el resto de los portainjertos o Celeste sobre MxM 60 o CAB 6P alcanzaron la media con 320 GDH > 30 °C. La variedad Celeste sobre G 5 lo hizo con solo 201 GDH > 30 °C y el resto de los portainjertos con 255 GDH > 30 °C.
La Figura 11 muestra que, en el sexto ADI, la SP media de Bing midió 10,8 % y la de Celeste 34,7 %. La variedad Bing sobre G 6 o Adara-M2624 redujo la Sp un 7,6 % y 6,8 % sobre la media de la variedad, respectivamente; mientras que sobre MxM 14 la incrementó un 9,6 %. La variedad Celeste sobre CAB 6P o Adara-M2624 redujo la Sp un 14,7 % y 2,9 % sobre la media varietal, respectivamente; pero sobre MxM 14, W 13, G 5 o G 6 la incrementó en 5,4 %, 6,5 %, 21,2 % y 14,7 %, respectivamente.
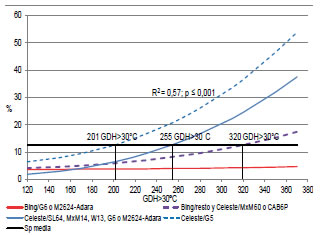
Figura 10: Relación GDH > 30 °C - Sp según grupo homogéneo de portainjerto-variedad

Figura 11: Efecto portainjerto-variedad sobre Sp del sexto ADI. Test de S&K (a= 0,05)
Análisis de porcentaje de espolón y dobles (Ep-Fd)
La media de EP-FD fue 1,67 ± 6,7 % lo que indica gran variabilidad de los datos. Sin embargo, la media de FD fue 0,36 ± 0,95 %, pero la de Ep fue
1,3 ± 5,8 % con datos que oscilaron entre 0 % y 90 %, indicando que en condiciones desfavorables puede alcanzar gran incidencia.
Ep asoció con FD (r = 0,32; p < 0,0001); sin embargo, SP no asoció con Ep (p = 0,3347) o con Fd (p = 0,6635). Esto indica que, aunque los tres defectos comparten una causa común, que es una elevada cantidad de calor durante la diferenciación floral, el porcentaje de un tipo de daño difiere según el momento en que se registra la alta temperatura, sea al inicio (EP o Fd) o al final de la temporada durante la cicatrización carpelar (SP). El efecto variedad asoció con E (r = -0,58; p = 0,0002) y con Fd (r = -0,43; p < 0,0001).
La variabilidad de EP-Fd se explicó con el modelo (R2a = 0,71; p = 0,0004) mediante los efectos ADI (p < 0,0001), portainjerto (p = 0,0026), variedad (p < 0,0001) e interacciones portainjerto-variedad (p = 0,0044), ADI-variedad (p < 0,0001) y ADI-portainjerto-variedad (p = 0,0173).
El EP-Fd del sexto ADI duplicó la media con un incremento del 1,9 %; mientras que en el séptimo y el octavo se registraron reducciones del 1,6 % y 0,3 %, respectivamente, con un comportamiento interanual semejante al de SP (Tabla 1). En la variedad Bing, considerada susceptible, registró valores 41 veces mayor que de Celeste (Tabla 1). La Figura 9 muestra que, durante los tres ADI, la variedad Celeste midió valores cercanos a cero; mientras que Bing del sexto y octavo ADI registró incrementos del 5,2 % y 1,1 % sobre la media, respectivamente, y en el séptimo se redujo un
1,5 %, con los correspondientes 371, 246 y 127 GDH > 30 °C según ADI, respectivamente. El comportamiento interanual de EP-Fd de Bing fue semejante al de SP de Celeste, indicando que el calor provoca defectos diferentes con intensidades distintas, según variedad. Alto GDH > 30 °C de post-cosecha incrementa pistilos dobles en Bing y cicatrización carpelar parcial en Celeste. En tal sentido, Southwick y Uyemoto (1999) sugirieron que estos defectos poseen un componente climático y genético (Engin et al., 2009; S0nstebya y Heideb, 2019).
Los portainjertos G 5 o G 6 indujeron incrementos de 1,6 % y 3,2 % sobre la media de Ep-Fd, respectivamente, mientras que el resto registró valores inferiores a la media.
La combinación de Celeste con los portainjertos de la serie no diferenció y registró valores cercanos a cero. Pero la variedad Bing, susceptible, sobre G 5 o G 6 la incrementó el 4,8 % y el 7,8 %, respectivamente sobre la media (Tabla 1). Nuevamente la evidencia indica que Ep-Fd diferenció portainjerto solo cuando la variedad fue susceptible.
La variedad Bing mostró una relación EP-Fd con GDH > 30 °C que ajustó con un modelo lineal cúbico (R2 = 0,25; p < 0,0001). No obstante, al ingresar la información aportada por los grupos homogéneos de portainjerto y sometida a una prueba de comparación de regresiones anidadas, se obtuvo un modelo (R2 = 0,48; p < 0,0001) que mejoró la calidad de información (p < 0,005) a través de la ordenada de origen. La Figura 12 muestra que Bing sobre G 5 o G 6 superó la media varietal de Ep-Fd aun, con bajas cantidades de GDH > 30 °C; mientras que sobre el resto de los portainjertos requirió 340 GDH > 30 °C solo para alcanzar la media. Por lo tanto, variedades susceptibles a Ep-FD no deberían combinarse con G 5 o G 6 en las condiciones locales.
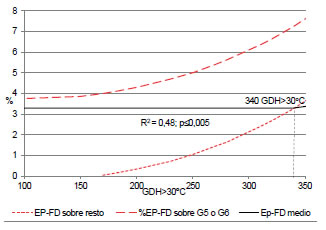
Figura 12: Relación GDH > 30 °C - Ep-Fd de Bing según grupo homogéneo de portainjerto
CONCLUSIONES
Los portainjertos influyen en el nivel tanto de daño como de defecto de las cerezas. Existen portainjertos capaces de reducir ambos simultáneamente y, en consecuencia, disminuir las pérdidas. Sin embargo, algunos portainjertos incrementan los daños, mientras que otros aumentan los defectos.
Según el tipo de daño o defecto analizado, el comportamiento de cada variedad y de cada portainjerto varió, mostrando diferentes grados de susceptibilidad. La interacción portainjerto-variedad diferenció combinaciones, pero solo cuando la variedad fue susceptible al daño o defecto evaluado. Esto indica que una variedad de alto valor económico, pero susceptible a un determinado tipo de daño o defecto, debe combinarse con un portainjerto que no acentúe la pérdida.
El nivel de daño depende de las condiciones climáticas, especialmente de las lluvias durante la temporada de cosecha y pre-cosecha. Mientras que el tipo y el nivel de defecto depende de las condiciones climáticas, especialmente de las temperaturas registradas durante la post-cosecha del año precedente.
En zonas con veranos cálidos, se debe evitar el cultivo de variedades susceptibles a defectos, así como también aquellos portainjertos que los inducen. En condiciones de alta oferta de GDH > 30 °C durante la post-cosecha, el tipo de defecto es altamente dependiente de la variedad.
AGRADECIMIENTO
Este trabajo fue posible gracias al apoyo incondicional de los Viveros Puertas, Agroviveros S.A. y Bobafrut S.R.L.
BIBLIOGRAFÍA
Ampatzidis, Y. y Whiting, M. (2013). Training system affects sweet cherry harvest efflciency. HortScience, 48(5), 547-555. https://doi.org/10.21273/HORTSCI.48.5.547
Anderson, A., Lindell, C., Moxcey, K., Siemer, W., Linz, G., Curtis, P, Carroll, J., Burrows, C., Boulanger, J., Steensma y K. and Shwiff, S. (2013). Bird damage to select fruit crops: The cost of damage and the beneflts of control in flve states. Crop Protection 52, 103-109. https://doi. org/10.1016/j.cropro.2013.05.019
Beppu, K. y Kataoka, I. (1999). High temperature rather than drought stress is responsible for the occurrence of double pistils in Satohnishiki' sweet cherry. Science Horticulturae, 81(2), 125-134. https://doi.org/10.1016/S0304-4238%2899%2900007-2
Bound, S., Close, D., Quentin, A., Measham, P. y Whiting, M. (2013). Crop Load and Time of Thinning Interact to Affect Fruit Quality in Sweet Cherry. Journal of Agricultural Science, 5(8), 216230. http://dx.doi.org/10.5539/jas.v5n8p216
Chiang, A., Schnettler, B., Mora, M. y Aguilera, M. (2018). Perceived quality of and satisfaction from sweet cherries (Prunus avium L.) in China: Confirming relationships through structural equations. Ciencia e investigación Agraria, 45(3), 210-219. http://dx.doi.org/10.7764/rcia.v45i3.1930 Centre Technique Interprofessionel des Fruits et Legumes (2005). Code du Couleur Cerise, (Éventail). Paris, Francia. Ref: 22006.
Correia, S., Schouten, R., Silva, A. P y Gongalves, B. (2018). Sweet cherry fruit cracking mechanisms and prevention strategies: A review.Scientia Horticulturae, 240, 369-377. https://doi. org/10.1016/j.scienta.2018.06.042
Crisosto, C., Crisosto, G. y Metheney, P (2003). Consumer acceptance of Brooks' and Bing' cherries is mainly dependent on fruit SSC and visual skin color. Postharvest Biology and Technology, 28(1), 159-167. http://dx.doi.org/10.1016/S0925-5214(02)00173-4
Di Rienzo, J., Casanoves, F, Balzarini, M., Gonzalez, L., Tablada, M. y Robledo, C. (2018). InfoStat (versión 2018) [Software]. Córdoba, Argentina: Grupo InfoStat, FCA, Universidad Nacional de Córdoba. https://www.infostat.com.ar/ Engin, H. y Ünal, A. (2008a). Double Fruit Formation and the Occurrence of Two Pistils: Examination by Scanning Electron Microscopy in Sweet Cherry. Acta Horticulturae, 795, 651-654. https://doi.org/10.17660/ActaHortic.2008.795.101 Engin, H. y Ünal, A. (2008b). The Effect of Irrigation, Gibberellic Acid and Nitrogen on the Occurrence of Double Fruit in Van' Sweet Cherry. Acta Horticulturae, 795, 645-650. http://dx.doi. org/10.17660/ActaHortic.2008.795.100
Engin, H., en, F, Pamuk, G. y Gokbayrak, Z. (2009). Investigation of Physiological Disorders and Fruit Quality of Sweet Cherry. European Journal of Horticultural Science, 74(3), 118-123. https://www. pubhort.org/ejhs/2009/file_1034231.pdf
Fajt, N., Folini, L., Bassi, G. y Siegler, H. (2014). Lapins' on ten cherry rootstocks in the Alpe Adria Región. Acta Horticulturae, 1020, 371-376. https:// doi.org/10.17660/ActaHortic.2014.1020.51
Gainza, F., Opazo, I., Guajardo, V, Meza, P, Ortiz, M., Pinochet, J.y Muñoz, C. (2015). Review: Rootstock breeding in Prunus species: Ongoing efforts and new challenges. Chilean Journal of Agricultural Research, 75(Suppl.1), 6-16. http:// dx.doi.org/10.4067/S0718-58392015000300002 Grant, J., Caprile, J., Coates, W., Anderson, K., Klonsky, K. y De Moura, R. (2001). Sample costs to establish an orchard and produce sweet cherries. San Joaquin Valley-North. University of California Cooperative Extension. CH-VN-11.
Instituto Nacional de Tecnología Agropecuaria (2007). Tabla de Colores de cerezas. Guía para la aplicación de ácido giberélico y para el inicio de la cosecha de variedades rojas. Laboratorio de Fruticultura, Estación Experimental Agropecuaria Mendoza, INTA. Cooperadora E.E.A. Mendoza.
Hussein, Z., Fawole, O. A. y Opara, U. L. (2020). Harvest and Postharvest Factors Affecting Bruise Damage of Fresh Fruits. Horticultural Plant Journal, 6(1), 1-13. https://doi.org/10.1016/j.hpj.2019.07.006
InnovaChile (2010). Programa de difusión tecnológica en Cerezos. Fundación para el desarrollo frutícola. http://www.fdf.cl/biblioteca/ publicaciones/2010/archivos/Poster_Danos_ Cerezas.pdf
Kappel, F. y Lang, G. (2008). Performance of the NC-140 Regional Sweet Cherry Rootstock Trial Planted in 1998 in North America. Acta Horticulturae, 795, 317-320. https://doi.org/10.17660/ActaHortic.2008.795.45
Koumanov, K., Staneva, I., Kornov, G. D. y Germanova, D. (2018). Intensive sweet cherry production on dwarfing rootstocks revisited. Scientia Horticulturae, 229, 193-200. https://www.researchgate.net/publication/322194047_ Intens¡ve_sweet_cherry_product¡on_on_dwarf¡ng_ rootstocks_revisited
Ljubojevi, M., Ognjanov, V, Bara, G., Duli, J., Miodragovi, M., Sekuli, M. y Jovanovi, N. (2016). Cherry tree growth models for orchard management improvement. Turkish Journal of Agriculture and Forestry, 40, 839-854. https://doi.org/10.1016/j. scienta.2017.11.009
López-Ortega, G., García-Montiel, F, Bayo-Canhaa, A., Frutos-Ruiz, C. y Frutos-Tomás, D. (2016). Rootstock effects on the growth, yield and fruit quality of sweet cherry cv. Newstar' in the growing conditions of the Region of Murcia. Scientia Horticulturae, 198, 326-335. https://doi. org/10.1016/j.scienta.2015.11.041
Lucero, G., Lucero, H. y Pizzuolo, P (2015). Enfermedades que afectan al cerezo en Mendoza. En Tacchini, F (Eds.), Producción y comercialización de cereza en Mendoza, Argentina (261-288). Fundación Instituto de Desarrollo Rural. https://www.idr.org.ar/wp-content/ uploads/2015/07/Manual-de-Producci%C3%B3n-y-Comercializaci%C3%B3n-de-Cerezas-Mendoza-part-1.pdf
Martínez Palanca, J. y Vilardell Cordech, P. (2008). Evaluación de aplicaciones de TaloSint para favorecer la formación de russeting en la pera Conference. Phytohemeroteca 197. https://www. phytoma.com/la-revista/phytohemeroteca/197-marzo-2008/evaluacion-de-aplicaciones-de-talosint-para-favorecer-la-formacion-de-russeting-en-la-pera-conference
Measham, P, Bound, A., Gracie, J. y Wilson, S. (2009). Incidence and type of cracking in sweet cherry (Prunus avium L.) are affected by genotype and season. Crop Pasture Science, 60(10), 10021008. https://doi.org/10.1071/CP08410
Measham, P, Gracie, A., Wilson, S. y Bound, A. (2010). Vascular flow of water induces side cracking in sweet cherry (Prunus avium L.). Advances in Horticultural Science, 24(4), 243-248. http://dx.doi. org/10.1400/153230
Measham, P., Bound, S., Gracie, A. y Wilson, J. (2013). Crop load manipulation and fruit cracking in sweet cherry (Prunus avium L.). Advances in Horticultural Science, 26(1), 25-31. http://dx.doi. org/10.13128/ahs-12749
Measham, P., Wilson, S., Gracie, A. y Bound, S. (2014). Tree water relations: Flow and fruit. Agriculture Water Management, 137, 59-67. https:// doi.org/10.1016/j.agwat.2014.02.005
Mendenhall, W. y Sincich, T (1995). Statistics for engineering and the sciences (4th. ed.). Prentice Hall.
Michailidis, M., Karagiannis, E., Tanou, G., Sarrou, E., Karamanoli, K., Lazaridou, A., Martens, S. y Molassiotis, A. (2020). Sweet cherry fruit cracking: follow-up testing methods and cultivar-metabolic screening. Plant Methods 16(1), 1-14. https://doi.org/10.1186/s13007-020-00593-6 Mertz, L. (2016). Saving cherries from the birds: New information provides help with bird management. Good Fruit Grower. https://www. goodfruit.com/saving-cherries-from-the-birds/
Neilsen, G., Neilsen, D., Kappel, F, Toivonen, P. y Herbert, L. (2010). Factors Affecting Establishment of Sweet Cherry on Gisela 6 Rootstock. HortScience, 45(6), 939-945. http:// dx.doi.org/10.21273/HORTSCI.45.6.939
Olmstead, M. A., Lang, N. S., Lang, G. A., Ewers, F W. y Owens, S. A. (2006). Examining the Vascular Pathway of Sweet Cherries Grafted onto Dwarfing Rootstocks. HortScience, 41(3), 674-679. http:// dx.doi.org/10.21273/HORTSCI.41.3.674
Rana, R. L., Andriano, A. M., Giungato, P. y Tricase, C. (2019). Carbon footprint of processed sweet cherries (Prunus avium L.): From nursery to market. Journal of Cleaner Production, 227, 900910. https://doi.org/10.1016/j.jclepro.2019.04.162 Sansavini, S. y Lugli, S. (2008). Sweet Cherry Breeding Programmes in Europe and Asia. Acta Horticulturae 795, 41-58. https://doi.org/10.17660/ ActaHortic.2008.795.1
Sansavini, S. y Lugli, S. (2014). New rootstocks for intensive sweet cherry plantations. Acta Horticulturae, 1020, 317-320. https://doi.org/10.17660/ActaHortic.2014.1020.59
Schumann, C. y Knoche, M. (2020). Swelling of cell walls in mature sweet cherry fruit: factors and mechanisms. Planta 251(3), 1-16. https://doi. org/10.1007/s00425-020-03352-y
Simon, G., Hrotkó, K. y Magyar, L. (2004). Fruit quality of sweet cherry cultivars grafted on four different rootstocks. Acta Horticulturae, 658(1), 365-370. http://dx.doi.org/10.17660/ActaHortic.2004.658.53
Simon, G. (2006). Review on rain induced fruit cracking of sweet cherries (Prunus avium L.), its causes and the possibilities of prevention. International Journal of Horticultural Science, 12(3), 27-35. http://dx.doi.org/10.31421/IJHS/12/3/654
S0nstebya, A. y Heide, O. M. (2019). Temperatura effects on growth and floral initiation in sweet cherry (Prunus avium L.). Scientia Horticulturae 257, 108762. https://doi.org/10.1016/j. scienta.2019.108762
Southwick, S. M., Shackel, K. A., Yeager J. T., D., Asai W. K. y Katacich, M. (1991). Over tree sprinkling reduces abnormal shapes in Bing' sweet cherries. California Agriculture 45(4), 24-26. http://dx.doi.org/10.3733/ca.v045n04p24
Southwick, S. y Uyemoto, J. (1999). Cherry crinkle-leaf and deep suture disorders. University of California. Agricultural and Natural Resources Publications.
Steiner, M., Magiar, L., Gyeviki, M. y Hrotkó, K. (2015). Optimization of light interception in intensive sweet cherry orchard. Scientific Papers. Serie B, Horticulture, 59, 105-108. http://horticulturejournal. usamv. ro/pdf/2015/art17.pdf
Wilkinson, L. (2002). TableCurve 2D (versión prueba) [Software]. SYSTAT Software Inc.
Tersoglio, E. y Naranjo, G. (2007). Características del frío invernal de las zonas productoras de cerezas de la provincia de Mendoza, Argentina. Parte I. Información Técnica Económica Agraria, 103(4), 186-197.
Tersoglio, E. y Setien, N. (2016). Efecto de la combinación Adara-M2624 y otros siete portainjertos sobre las características del dosel de dos variedades de cerezo. Agriscientia, 33(2), 113-125. https://doi.org/10.31047/1668.298x.v33. n2.16578
Villar, L., Lienqueo, I., Llanes, A., Rojas, P, Perez, J., Correa, F, Sagredo, B., Masciarelli, O., Luna, V. y Almada, R. (2020). Comparative transcriptomic analysis reveals novel roles of transcription factors and hormones during the flowering induction and floral bud differentiation in sweet cherry trees (Prunus avium L. cv. Bing). PLoS ONE,15(3), e0230110. https://doi.org/10.1371/journal. pone.0230110
Winkler, A., Brüggenwirth, M., Ngo, N. S. y Knoche, M. (2016). Fruit apoplast tension draws xylem water into mature sweet cherries. Scientia Horticulturae, 209, 270-278. http://dx.doi.org/10.1016/j.scienta.2016.06.041
Winkler A., Peschel, S., Kohrs, K. y Knoche, M. (2016). Rain Cracking in Sweet Cherries is not Due to Excess Water Uptake but to Localized Skin Phenomena. Journal of American Society Horticultural Science, 141(6), 653-660. http:// dx.doi.org/10.21273/JASHS03937-16
Winkler, A. y Knoche, M. (2019). Calcium and the physiology of sweet cherries: A review. Scientia Horticulturae, 245, 107-115. https://doi. org/10.1016/j.scienta.2018.10.012
Winkler, A., Blumenberg, I., Schürmann, L. y Knoche, M. (2020). Rain cracking in sweet cherries is caused by surface wetness, not by water uptake. Scientia Horticulturae, 269, 109400. https://doi. org/10.1016/j.scienta.2020.109400
Wirch, J., Kappel, F. y Scheewe, P. (2009). The Effect of Cultivars, Rootstocks, Fruit Maturity and Gibberellic Acid on Pedicel Retention of Sweet Cherries (Prunus avium L.). Journal of American Pomological Society, 63(3), 108-114.
Wyman, O., Münch, R., Pohl, A., Terreaux, G. y Torchalla, J. (2019). Fruit Logistica Trend Report 2019. Surprises in Store. Fruitnet Media International. https://www.oliverwyman.com/content/dam/ oliver-wyman/v2-de/publications/2019/Februar/ FruitLogisticaTrendReport_2019_OliverWyman_ EN.pdf

