










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkInVet
versión On-line ISSN 1668-3498
InVet v.10 n.2 Ciudad Autónoma de Buenos Aires jul./dic. 2008
Hallazgos en la coherencia cerebral en caninos con comportamiento agresivo
Rivero, J.E.1; Vidal Figueredo, R.J.2; Pellegrino, F.C.3
1 Ayudante de segunda, Área de anatomía- FCV, U.B.A. Chorroarin 280, Ciudad Autónoma de Bs. As.
2 Ayudante de Primera, Área de anatomía- FCV, U.B.A.
3 Profesor Titular, Área de anatomía- FCV, U.B.A.
Recibido: 18/07/08
Aceptado: 02/12/08
Resumen
La finalidad de este trabajo es realizar un estudio comparativo del electroencefalograma (EEG), entre animales sanos y agresivos, utilizando la coherencia cerebral interhemisférica para el análisis cuantitativo.
Se trabajó con 24 animales sin trastornos de comportamiento y 24 animales agresivos sin alteraciones orgánicas evidentes. Todos resultaron normales al examen físico, neurológico y ninguno tenía antecedentes de enfermedad neurológica.
Al igual que lo que sucede en los animales sin trastornos del comportamiento, los animales agresivos presentaron los valores de frecuencia del pico de potencia cruzado (PPC) similares en los canales dorsales y temporales. Por el contrario se hallaron valores de coherencia disminuidos y de espectro de fase aumentados entre los canales temporales de los animales agresivos respecto de los animales sin alteraciones del comportamiento.
El estudio de coherencia cerebral se presenta como una herramienta sencilla y no invasiva, capaz de evidenciar alteraciones objetivas y cuantificables en el trazado de base del EEG.
Palabras clave: Caninos agresivos; Coherencia Cerebral intrahemisferica; Electroencefalografía cuantificada; Espectro de potencia.
Cerebral coherence finding asocied to agressive behaviour in dogs
Summary
We used the interhemispheric coherence to investigate the difference between normal and aggressive dogs in base to quantify the base line of electroencephalogram.
Dorsal and temporal group was defined by significant statistic difference in the frequency spectrum. Temporal channels showed low interhemispheric coherence and phase delay in aggressive dogs.
Coherence is an easy and noninvasive technique capacious to proved and quantified alteration in electroencephalogram base line.
Key words: Cerebral interhemispheric coherence; Aggressive dogs; Quantitative electroencephalography; Spectral analysis.
Introducción
En medicina veterinaria la agresividad es el trastorno del comportamiento más frecuente en caninos, y constituye un problema de importancia creciente(14). Esto se debe a que un perro agresivo puede poner en peligro la integridad física de una persona, llegando en casos extremos a ocasionarle la muerte.
La medicina veterinaria tradicionalmente ha enfocado el problema de la agresión desde un punto de vista exclusivamente sanitarista, apuntando al control de una enfermedad zoonótica de extrema gravedad, la rabia. Sin embargo debido a que el único procedimiento metodológico utilizado en los casos de perros mordedores es la denominada «observación antirrábica», la agresión canina en sí misma sigue siendo un problema relevante.
Según el Ministerio de Salud de la Nación, se reportan al año alrededor de 60000 mordeduras de perros (casi siete por hora)(14). La mitad de las agresiones se produce contra chicos menores de 14 años y la mayoría de los mordedores son mascotas de la familia o animales que tienen dueño. Un informe de la provincia de Buenos Aires considera que alrededor del 70% de las mordeduras no se denuncian(14). Un estudio realizado en Pensil- vania, Estados Unidos, mostró que sobre un total de 3000 niños en edad escolar el 45% habla sido mordido por un perro durante su vida; de ellos el 30% había sido mordido por su propio perro(52). En Argentina, en un estudio realizado en Capital Federal y Gran Buenos Aires el 30% de los propietarios de perros manifestó que sus animales habían manifestado episodios agresivos(15).
Esta realidad nos indica que resulta de imperiosa necesidad ampliar el enfoque acerca del comportamiento agresivo en los caninos. Para ello es necesario comprender que la agresión de los perros hacia los seres humanos es un fenómeno complejo que incluye numerosas variables (genética, orgánica, aprendizajes, vínculo con los humanos, etc.), que puede ser de diversos tipos (agresión jerárquica, irritable, por temor, sobreprotectora, por causas orgánicas o funcionales, etc.) y que no sólo puede ser el vehículo de transmisión de una zoonosis sino que debería ser considerada como una zoonosis en si misma generada por posibles trastornos del comportamiento canino.
Definimos al canino agresivo como «aquel individuo que manifiesta actitudes conductuales dirigidas a lesionar una entidad tomada como objetivo»(29). Nos hemos interesado en el estudio de aquellos perros agresivos que no presentan alteraciones orgánicas evidentes, sospechando que tal vez existan aspectos objetivos y cuantificables de su función eléctrica cerebral (evaluada a través del electroencefalograma -EEG-) que se desvíen de los valores hallados para caninos sin alteraciones de comportamiento.
El EEG, a pesar de sus aparentes fluctuaciones azarosas es, desde el punto de vista estadístico, un proceso muy ordenado. La composición de su espectro de frecuencias es predecible por medio de ecuaciones normativas para cada área cerebral. Por lo tanto el EEG, registrado simultáneamente desde varias regiones cerebrales, nos provee de una visión funcional del sistema nervioso central (SNC) característica de la especie, de su desarrollo y su maduración.(30,36,40)
Considerando la interacción entre regiones corticales como el reflejo de la interacción de sistemas neuroanatómicos multifuncionales, y que los mismos dependen de la combinación de diferentes neurotransmisores, el EEG provee potencialmente acceso cuantitativo al estado funcional de numerosos sistemas. Desvíos de los parámetros electrofisiológicos normativamente predecibles indicarían desbalances específicos en las reacciones neuroquímicas que median la comunicación en estos sistemas.(36,40)
Actualmente se considera que las diferentes formas de cuantificar el EEG pueden tener gran utilidad potencial en la detección de la actividad normal y la disfunción, permitiendo la detección temprana de muchas enfermedades, su caracterización cuantitativa y el seguimiento del grado de la anormalidad, como resultado o no de una terapéutica.
Entre los métodos matemáticos aplicados al análisis del EEG, ha adquirido importancia la utilización del análisis de coherencia que evalúa la asociación lineal entre los registros obtenidos a parir de canales ubicados en diferentes posiciones(10,11,12,16,27), independientemente de la amplitud de los registros obtenidos en cada localización(10).
El análisis de coherencia informa sobre los componentes frecuenciales comunes a ambos canales (pico de potencia cruzada [PPC]), junto con la magnitud de su asociación en lo que se refiere a simetría y sincronía (espectro de coherencia y espectro de fase). Esta herramienta ha sido correlacionada con la existencia de generadores comunes y con el grado de interconexión biológica entre las regiones corticales representadas por dichos canales(10,27).
En este trabajo nos proponemos evaluar la actividad eléctrica cerebral en caninos agresivos, utilizando la coherencia interhemisférica como herramienta para el estudio de posibles alteraciones funcionales del sistema nervioso central.
Materiales y métodos
Muestra
Se trabajó con 24 animales sin trastornos de comportamiento mayores de un año de edad, de distintas razas, y 24 animales agresivos sin alteraciones orgánicas. Todos los perros resultaron normales al examen físico general y neurológico. Ninguno de ellos tenía antecedentes de enfermedad neurológica; a todos se les realizó análisis de laboratorio (hemograma, perfil bioquímico sanguíneo y perfil tiroideo) que resultaron dentro de los valores normales en todos los casos. A todos los animales se les realizó EEG, que en ninguno de los casos presentó alteraciones al análisis visual.
Equipamiento
Para obtener los registros EEG se utilizó un programa especialmente diseñado para electroencefalografía computada y reconstrucción por mapeo cerebral (AKONIC BIO- PC versión modificada 7.0), con 12 canales simultáneos de registro. El sistema permite promediar y procesar muestras múltiples. El equipo incluye 12 canales EEG unidos a una interfase analógico-digital y el sistema de computación. Los parámetros técnicos del registro fueron los siguientes: Rechazo de modo común > de 100 db; respuesta de frecuencia de 0,5 128 Hz; filtro antialiasing: 36 db/octava; filtro de alta frecuencia (FAF): 12 db/octava; filtro de baja frecuencia (FBF): 6 db/octava; ruido propio < 1 uV; frecuencia de muestreo: 256 s; velocidad del papel: 30 mm/s; impedancia de los electrodos: < 5 kΩ.
Procedimiento
Terminología y posición de los electrodos de registro. El tipo de electrodos y su colocación se realizó de acuerdo a estudios y recomendaciones previas(32,34,35,36).
Registro. El registro básico se efectuó empleando montaje de referencia común, con electrodo de referencia nasal. Esto es indispensable para la cuantificación del EEG, ya que de este modo todos los electrodos tienen una misma referencia, que es utilizada como patrón para comparar el voltaje en los diferentes canales(6). El tiempo mínimo de registro fue de 30 minutos.
Restricción química de los animales. Se realizó por medio de inyección subcutánea de xylazina (Rompún®, Bayer Argentina S.A.), basándonos en trabajos previos(7,17,18,23,44) y en nuestra propia experiencia(33), en dosis de 1 mg/kg. Se considera que la xylazina es un agonista de receptores á2 adrenérgicos(7,17,23) que no tiene efectos directos sobre las neuronas corticales piramidales(45).
Análisis espectral del EEG
Para cuantificar los EEG de cada animal se seleccionaron al menos 15 segmentos estacionarios de 2 segundos de duración libres de artificios (épocas), promediando los resultados de cada época(40). Luego de la selección se realizó la estimación espectral aplicando a los segmentos seleccionados la Transformada rápida de Fourier.(6)
Análisis de la coherencia
Con los datos obtenidos a partir del espectro de potencia se realizó en cada animal el análisis de coherencia interhemisférica para cada par de canales Temporales (T3-T4), Frontopolares (Fp1-Fp2), Frontales (F3-F4), Parietales (P3-P4), Occipitales (O1-O2)(36). En cada uno de ellos se determinaron las armónicas comunes a ambos, superponiendo los espectros de potencia de cada canal (espectro de potencia cruzada). Se seleccionó entonces la armónica en común de mayor potencia, que corresponde a una determinada frecuencia (pico de potencia cruzada -PPC-). A esa frecuencia corresponde también un determinado valor de coherencia propiamente dicha (espectro de coherencia), que varía entre 0 y 1, y un determinado espectro de fase, expresado en grados o, de manera más práctica, en milisegundos. En consecuencia, para cada par de canales se obtuvieron 3 valores: la armónica de mayor potencia (PPC, expresado en hertz), la coherencia correspondiente a esa frecuencia y el desfasaje existente entre las 2 señales consideradas (expresado en grados o milisegundos), el sino + o - indica a qué hemisferio llega primero la señal eléctrica (levo y dextro procedencia).(10)
Análisis estadístico
En una primera instancia se realizaron los estudios de estadística descriptiva correspondientes para observar el comportamiento de las variables en estudio. Las herramientas graficas utilizadas (box and whisker plot y rankit plot) indican que las distribuciones de la variables en estudio son aproximadamente simétricas. Los valores de will-shapiro tendientes a 1 confirman que el análisis paramétrico, basado en la normalidad de los datos, es adecuado.
En una segunda instancia se utilizó un modelo de parcelas divididas, considerando a cada animal como una unidad, y a cada canal como una subunidad dentro de cada animal. Dada la imposibilidad de aleatorizar, y la existencia de una matriz de variancias y covariancias (por la dependencia entre las respuestas en las distintas subunidades dentro de cada unidad), el problema se planteó como medidas repetidas o parcelas divididas en el tiempo(13) donde se evaluó el factor estado de los caninos de la muestra (normales/agresivos) y el factor canal. En los casos en que no se cumplió el supuesto de «esfericidad» (compound simetry) se corrigieron los grados de libertad en la comparación de los canales o en la interacción si la misma existía(19,42). Para evaluar diferencia entre las diferentes combinaciones de canales se realizaron contrastes generales. El nivel de significación establecido como significativo en todas las pruebas fue p≤ 0,05.
Resultados
Los valores de pico de potencia cruzada (PPC) son similares en los animales normales y en los agresivos (F: 1,67; P: 0,20). En ambos grupos la frecuencia a la que se presenta en los canales temporales (T) (0 5,11 Hz ± 0,26) es inferior a la de los canales dorsales (0 7,18 Hz ± 0,18) (P:<0,001). (F: 20,83; P:<0,001) (Ver gráfico 1) Los valores hallados para la frecuencia del PPC en los electrodos T es de 5.11 Hz, lo cual incluye al valor del PPC dentro de la banda electroencefalográfica theta (4-7 Hz). Los electrodos dorsales registran información eléctrica fundamentalmente de la neocorteza dorsal(33); el PPC para este grupo de canales se presenta a una frecuencia de 7.50 Hz, que lo sitúa entre las banda theta y alpha (4 a 7 Hz y 8 a 13 Hz, respectivamente) Al analizar el espectro de coherencia se observa interacción entre los factores evaluados (F: 4,66; P: 0,001). Los canales T de los perros agresivos presentan un valor inferior (0 0.48 ± 0,038) en relación a la hallada en el mismo par de canales en los animales normales (0 0,59 ± 0,037) (ver gráfico 2). Respecto al análisis del espectro de fase se observa efecto estado (F: 6,11; P: 0,017) y efecto canal (F: 158,06; P:<0,001), sin que haya interacción entre ellos. En ambos grupos de animales los canales T presentan valores superiores a los del resto de los canales. Pero además, los canales T de los animales agresivos (0 28,57 ms ± 7,07) muestran valores significativamente (P < 0.001) superiores en relación al mismo par de canales de los animales sin alteraciones (0 13,45 ms ± 4,04) (Ver gráfico 3 A y B).
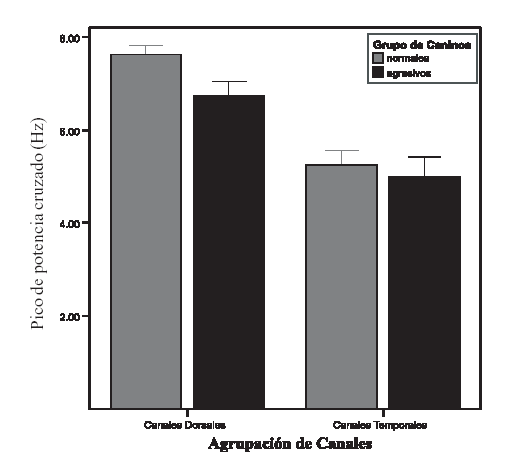
Gráfico 1: El gráfico de barras muestra el comportamiento del pico de potencia cruzado (PPC) para los animales normales y los animales agresivos. En ambos grupos de caninos los canales dorsales presentan el PPC a un valor más elevado que los canales temporales (F: 20,83; P:<0,001). No existen diferencias significativas en el valor del PPC al evaluar las agrupaciones de canales para ambos grupos de animales (F: 1,67; P: 0,20). Las barras muestran le valor de la media y las líneas el desvío estándar de la media.
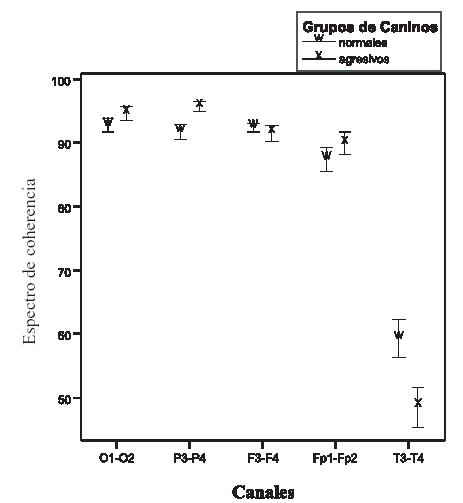
Gráfico 2: el gráfico de líneas muestra el comportamiento de espectro de coherencia para los diferentes canales. Las letras iguales indican similar comportamiento entre los grupos de canales; el * indica diferencias en el valor entre los grupos de caninos evaluados. Ambos casos según ANOVA (P< 0,05). Las barras muestran el error estándar de la media
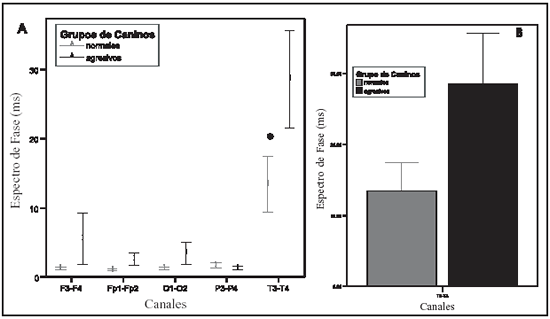
Gráfico 3: A) el gráfico de líneas muestra el comportamiento del espectro de fase para las distintas combinaciones de canales. No existes diferencia significativa entre los canales dorsales. En ambos grupos caninos los canales temporales son significativamente diferentes a los dorsales (*). B) el gráfico de barras muestra la diferencia en el comportamiento del espectro de fase de las canales T3- T4, obsérvese el aumento de la fase en el grupo de caninos agresivos. (P < 0.001)
Discusión
Al analizar la frecuencia a la que se presenta el PPC no hemos hallado diferencias entre los dos grupos de perros estudiados. En los perros agresivos, entonces, se pueden agrupar los pares de canales del mismo modo en que se agrupan en los animales sin alteraciones. Tenemos así al grupo de canales temporales y el resto de ellos, denominados dorsales.(50) Para el resto de los elementos del análisis de coherencia no hemos hallado ninguna diferencia entre los canales dorsales de ambos grupos de perros. El espectro de coherencia siempre fue mayor a 0,8, mientras que el espectro de fase nunca fue en promedio superior a 2,23 ms. (± 0,52). En cuanto a los canales que registran actividad de la región temporal, los animales agresivos presentan una significativa disminución de la coherencia interhemisférica (0,4) que se asocia a un notorio incremento del espectro de fase (28 ms). Sin embargo, como se mencionó más arriba, la frecuencia a la que se presenta el PPC es similar, ubicándose en la banda de frecuencia theta. Las principales áreas involucradas en la generación del ritmo theta son las neuronas piramidales de CA1 y CA3 en el hipocampo (1,2,3,21,26,51), que se hallan bajo el control del área septal, que actúa como marcapasos (1,20,26). A su vez la región septal se halla modulada por estructuras centroencefálicas, distribuidas a lo largo del tronco encefálico(4,28,47,48,49). Los hallazgos relativos al espectro de coherencia y al espectro de fase en los canales T podrían reflejar dos situaciones posibles:
a) que la señal, aun generándose en la misma región anatómica, atenúe o pierda la modulación centroencefálica, acentuándose de este modo su carácter focal. En varios trabajos se ha demostrado que el marcapasos septal puede ser modulado por la modificación de las aferencias provenientes del tronco encefálico,(37,38) demostrando a su vez que dichas modificaciones se extienden a las células piramidales del hipocampo, modificando su actividad y por lo tanto el ritmo theta. Efectos similares se han visto al modificar la actividad del núcleo supramamilar (37).
b) Que exista otra región anatómica distinta a los campos CA1 y CA3 que genera una señal situada en el mismo rango frecuencial, pero de características diferentes al ritmo theta original.
En algunos trabajos se ha mencionado que el ritmo theta puede aparecer en áreas neocorticales cuando el núcleo reticular del tálamo produce la hiperpolarización de las membranas mediada por acido gammaaminobutirico (GABA)(22).
En cualquiera de los casos, los hallazgos son compatibles con diversos estudios que destacan la susceptibilidad de las regiones temporales a diversas patologías, en virtud de sus características anatomofisiologicas(31). Esta región, como parte del sistema límbico, es el sustrato anatómico de muchas de las conductas típicas de la especie
Por otra parte, se ha descripto que anormalidades tales como disminución de la perfusión, isquemia y alteraciones en el metabolismo de la glucosa en áreas corticales temporoparietales( 8,9,25,39), o atrofia del hipocampo( 5,24,46) generan focos de actividad lenta asociados al aumento en el espectro de potencia de la banda theta.
Conclusiones
Hemos encontrado diferencias en los patrones de actividad cerebral entre los caninos agresivos y los caninos sin trastorno de comportamiento, cuantificadas a partir del trazado EEG de base. Tales diferencias se encuentran en la corteza temporal que, como se menciono anteriormente, es más susceptible a sufrir patologías que otras regiones cerebrales. Los valores hallados para los distintos elementos de la coherencia interhemisférica en los caninos agresivos para los canales T podrían deberse a la pérdida de modulación centro-encefálica, o a la aparición de un nuevo generador eléctrico que produce oscilaciones en el rango frecuencial del ritmo theta. La coherencia cerebral se presenta como una herramienta sencilla y no invasiva, capaz de evidenciar alteraciones en el trazado de base del EEG, detectando diferencias sutiles cuando se comparan los valores hallados en los animales agresivos contra una base de datos normativa obtenida a partir de individuos normales y sin alteración de comportamiento. De esta manera es posible cuantificar en forma objetiva las relaciones normales y anormales de distintas estructuras encefálicas, aún en individuos sin alteraciones al análisis visual de un trazado EEG.
1. Arnolds, D.E.; Lopes da Silva, F.H.; Aitink, J.W.; Kamp, A. Hippocampal EEG and behaviour in dog. II. Hippocampal EEG correlates with elementary motor acts. Electroencephalogr Clin Neurophysiol. 1979 May; 46(5):571-580. [ Links ]
2. Arnolds, D.E.; Lopes da Silva, F.H.; Aitink, J.W.; Kamp, A. Hippocampal EEG and behaviour in dog I. Hippocampal EEG correlates of gross motor behaviour. Electroencephalogr Clin Neurophysiol. 1979 May;46(5):552-570 [ Links ]
3. Arnolds, D.E.; Lopes da Silva, F.H.; Aitink, J.W.; Kamp, A. Hippocampal EEG and behaviour in dog. III. Hippocampal EEG correlates of stimulus-response tasks and of sexual behaviour. Electroencephalogr Clin Neurophysiol. 1979 May; 46(5):581-591. [ Links ]
4. Assaf, S.Y.; Miller, J.J. Role of a raphe serotonin system in control of septal unit activity and hippocampal desynchronization. Neuroscience 1978. 3:539-550. [ Links ]
5. Breslau, J.; Starr, A.; Sicotte, N. et al: Topographic EEG changes with normal aging and SDAT. Electroencephalogr Clin Neurophysiol 1989; 72:281±289 [ Links ]
6. Casaglia, D.C. La adquisición de datos para electroencefalografía computada. En «Análisis computado del EEG». Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. 1995. 54-76 [ Links ]
7. Celly, C.S.; McDonnel, W.N.; Young, S.S.; Black, W.D. The comparative hypoxemic effect of four a2 adrenoceptor agonists (xylazine, romifidine, detomidine and medetomidine) in sheep. J.Vet.Pharmacol. Therap. 1997. 20: 464-471 [ Links ]
8. Coben, K.A.; Danziger, W.L.; Storandt, M.A.; A longitudinal EEG study of mild senile dementia of Alzheimer type: changes at 1 year and at 2.5 years. Electroencephalogr Clin Neurophysiol 1985; 61:101-112 [ Links ]
9. Erkinjuntti, T.; Larsen, T.; Sulkava, R.; et al: EEG in the differential diagnosis between Alzheimer's disease and vascular dementia. Acta Neurol Scand 1988; 77:36-43 [ Links ]
10. Etchepareborda, M. «El análisis de la coherencia interhemisférica». En «Análisis computado del EEG». Cap. 6 Editado por Ferrero, R.G.A.; Ferrero, A.L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 83-91. [ Links ]
11. Etchepareborda, M.; Etchepareborda, J.; González, S. «El análisis de la coherencia interhemisférica en pacientes con disfasia infantil. Presentación de subtipos». Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As., Argentina. 1992. [ Links ]
12. Etchepareborda, M.; Etchepareborda, J.; González, S. «Relación entre el espectro de coherencia de la actividad bioeléctrica cerebral y los hallazgos neurofisiológicos en pacientes con síndrome disatencional e hiperactividad». Rev. Neurol. Barcelona. 1994. 22 (115): 266-271. [ Links ]
13. Fabrizio, M.C. y Garsd, A. Dos Factores con Medidas Repetidas en uno de ellos en un Diseño Completamente al Azar. Rev. Facultad de Agronomía 1996 16 (3): 243-252. http://www.fvet.uba.ar/ [ Links ]
14. Gerzovich Lis, C.J. Agresión en perros: su problemática. http://www.portaldog.com.ar/etologia/Agresion_en_perros.htm [ Links ]
15. Gerzovich Lis, C.J.; Rosa, I.D.; Cignacco, G.A. El vínculo ser humano-perro (Canis lupus familiaris) y su relación con los problemas de comportamiento. Revista de Medicina Veterinaria. 1997. Vol.78; N°22: 122-132 [ Links ]
16. Gotman, J. Análisis del EEG asistido por computadora (Monitoreo automático de espigas). En «Análisis computado del EEG». Cap. 21. Sección V, Editado por Ferrero, R.G.A.; Ferrero, A.L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As., Argentina. 1995. 241-257. [ Links ]
17. Green, J.D. and Arduini, A. Hippocampal electrical activity in arousal. J. Neurophysiol.1954; 17: 533-557 [ Links ]
18. Greene, S.A.; Thurmon, J.C.; Xylazine, A review of its pharmacology and use in veterinary medicine. J. Vet. Pharmacol. Therap. 1988. 11: 295-313. [ Links ]
19. Greenhouse, S.W. & Geisser, S. On Methods in the Analysis of Profile Data. Psychometrika 1959 24 (2): 95-112. [ Links ]
20. Holmes, J.E. and Adey W.R. Electrical activity of the entorhinal cortex during conditioned behaviour. Am. J. Physiol. 1960 Nov; 199: 741-744. [ Links ]
21. Holsheimer, J.; Stok, C.J.; Lopes da Silva, F.H. Theta rhythm related hippocampal cell discharges in the urethane anaesthetized rat: evidence for a predominant entorhinal input. Electroencephalogr. Clin. Neurophysiol. 1983 Apr; 55(4):464-467. [ Links ]
22. Hughes, J.R.; Roy John, E. Conventional and Quantitative Electroencephalography in Psychiatry. J Neuropsychiatry Clin Neurosci 1999; 11:190-208. [ Links ]
23. Jarvis, N.; England, G.C.W. Reversal of Xylazine sedation in Dogs. The Veterinary Record, April 1991. 128: 323-325. [ Links ]
24. John, E.R.; Prichep, L.S.; Ahn, H. et al: Neurometric evaluation of cognitive dysfunctions and neurological disorders in children. Prog Neurobiol 1983; 21:239-290 [ Links ]
25. Leuchter, A.F. Spar, J.D.; Walter, D.O. et al: Electroencephalographic spectra and coherence in the diagnosis of Alzheimer'stype and multi-infarct dementia. Arch Gen Psychiatry 1987; 44:993-998 [ Links ]
26. Lopes da Silva, F.H.; Witter, M.P.; Boeijinga, P. H.; Lohman, A. H. Anatomic organization and Phisiology of the Limbic Cortex. Physiol. Rev. 1990, Apr. 17(2) 453-511. [ Links ]
27. Lopes da Silva. EEG Analysis: theory and Practice. En «Electroencephalography». Cap 61. Edit by Niedermayer, E.; Lopes da Silva, F. William y Wilkins. Philadelphia. Third edition. 1993. 1097-1123 [ Links ]
28. Macadar, A.W.; Chalupa, L.M.; Lindsley, D.B. Differentiation of brain stem loci which affect hippocampal and neocortical electrical activity. Exp Neurol 1974. 43:499-514 [ Links ]
29. Moyer, K.E. Kinds of aggression and their physiological basis. Commun. Behav. Biol. Ed. Carnegie-Mellon University 1968. ASIN: B0007FETXM. Part. A. 2-65, [ Links ]
30. Murer, M.G.; Riquelme, L.A. Generalidades. En «Análisis computado del EEG». Cáp. 3. Editado por Ferrero, R.G.A.; Ferrero, A.L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 41-53. [ Links ]
31. Niedermayer, E. Epileptic seizure disorders. En «Electroencephalography». Cap . Edit by Niedermayer, E.; Lopes da Silva, F. William y Wilkins. Philadelphia. Third edition. 1993. 461-564. [ Links ]
32. Pellegrino, F.; Etchepareborda, M. Anatomical sites for the placement of electroencephalographic record electrodes in canines. Braz. J. Morphol. Sci. 1997.Vol.14, 2: 197-204. [ Links ]
33. Pellegrino, F.C.; Sica, R. Canine electroencephalographic recording technique: findings in normal and epileptic dogs. Clinical Neurophysiol. 2004; 115:77- 487 [ Links ]
34. Pellegrino, F.C. «Epilepsia y Síndromes epilépticos». En «El Libro de Neurología para la Práctica Clínica», ED. por Pellegrino, F. C.; Suraniti, A.; Garibaldi, L.; parte III, Cap. 17. Editorial Intermédica. Bs. As. Argentina. 2003. 255-280. [ Links ]
35. Pellegrino, F.C. Epilepsia y Síndromes Epilépticos III. Diagnóstico, Pronóstico y Tratamiento de la Epilepsia y los Síndromes Epilépticos. Selecciones Veterinarias, 1999. Vol.7. Nº 6: 686-704. [ Links ]
36. Pellegrino, F.C. Estandarización de los patrones electroencefalográficos de los caninos. Tesis doctoral. U.B.A. 2000 [ Links ]
37. Petsche, H.; Gogolak, G. & Van Zwieten, P.A. Rhythmicity of septal cell discharges at various levels of reticular excitation. Electroencephalography and Clinical Neurophysiology, 1965.19: 25-33. [ Links ]
38. Petsche, H.; Stumpf, C. & Gogolak, G. Significance of rabbit's septum as a relay station between midbrain and hippocampus: Control of hippocampus arousal activity by septum cells. Electroencephalography and Clinical Neurophysiology, 1962.14:202-211. [ Links ]
39. Robinson, D.J.; Merskey, H.; Blume, W.T. et al: Electroencephalography as an aid in the exclusion of Alzheimer's disease. Arch Neurol 1994; 51:280-284 [ Links ]
40. Roy John, E.; Prichep, L.S. Los principios del análisis neurométrico en EEG y potenciales evocados. En «Análisis computado del EEG». Cap 8. Editado por Ferrero, R. G. A.; Ferrero, A. L. Editor FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 111-135. [ Links ]
41. Ruckebush, Y.; Malbert, C.H.; Baha, F. Quantified electrocortical activity and loss of vigilance in dog and sheep. Revue de Medicine Veterinaire. 1987.138 (4): 325-332. [ Links ]
42. Steel, R.G.D. & Torrie, J.H. Principles and Procedures of Statistics (with special reference to the biological sciences) Ed Mc Graw Hill 1960 [ Links ]
43. Tatcher, R. Psychopatology of early frontal love damage: dependence on cyles of development. Dev. Psychopatol. 1994 6, 565-596 [ Links ]
44. Tierney, Patrick, L.; Dégenètais, Eric; Thierry, Anne-Marie; Glowinski, Jacques and Gioanni, Yves. Influence of the hippocampus on Interneurons of the rat prefrontal cortex. European Journal of Neuroscience 2004. 20: 514-524 [ Links ]
45. Tourai, K.; Senba, H.; Sasaki, N.; Tokuriki, M.; Ohashi, F.; Takeuchi, A.; Usui, K. Developmental EEG of the Beagle dog under xylazine sedation. Japanese Journal of Veterinary Science. 1985. 77 (3): 459-463. [ Links ]
46. Valdes-Sosa, P.; Bosch, J.; Grave, R. et al: Frequency domain models of the EEG. Brain Topogr 1992; 5:309-319 [ Links ]
47. Vertes, R.P. An analysis of ascending brain stem systems involved in hippocampal synchronization and desynchronization. J Neurophysiol 1981. 46:1140-1159. [ Links ]
48. Vertes, Robert, P. Hippocampal Theta Rhythm: A Tag for Short- Term Memory. Hippocampus 2005. 15:923-935 [ Links ]
49. Vertes, Robert, P.; Hoover, Walter, B.; Viana Di Prisco, Gonzalo. Theta Rhythm of the Hippocampus: Subcortical Control and Functional Significance Behavioral and Cognitive Neuroscience Reviews 2004. 3. 3: 173-200 [ Links ]
50. Vidal Figueredo, R.; Pellegrino, F.; Riquelme, L. Análisis y estandarización de los valores de coherencia cerebral interhemisférica en caninos adultos. -aceptado en INVET2007 -ISSN (soportepapel):1514-6634;ISSN (on line): 1668-3498 [ Links ]
51. Winson, J. Patterns of hippocampal tetha rhythm in the freely moving rat. Electroencephalographic clinical neurophysiology. 1974. 36: 291- 301 [ Links ]
52. Wright, J. Canine Aggression Toward People. Veterinary Clinics of North America: Small Animal Practice 1991. Vol. 21, N° 2. 299-314 [ Links ]

