










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkInVet
versión On-line ISSN 1668-3498
InVet v.11 n.2 Ciudad Autónoma de Buenos Aires jul./dic. 2009
ARTÍCULO DE INVESTIGACIÓN
Análisis de los valores de coherencia cerebral intrahemisférica en caninos adultos. Dinámica dual de la actividad eléctrica cerebral
Vidal Figueredo, R. J.1; Pellegrino, F. C.2
1 Veterinario; Ayudante de primera, Área de Anatomía. Facultad. de Ciencias Veterinarias. Universidad de Buenos Aires, Argentina. Chorroarin 280 (1427), Buenos Aires, e-mail: rvidal@fvet.uba.ar
2 Ph. D. Medico Veterinario; Profesor Titular, Área de Anatomía. Facultad de Ciencias Veterinarias. Universidad de Buenos Aires, Argentina.
Recibido: 18/11/09
Aceptado: 16/02/10
Resumen
Este trabajo tiene como finalidad investigar y describir el comportamiento de la actividad eléctrica intrahemisférica en caninos adultos normales utilizando como variable de estudio el análisis de coherencia cerebral para analizar la actividad de base del trazado electroencefalográfico. Hemos añadido algunas hipótesis que intentan relacionar los hallazgos con las conexiones anatómicas involucradas.
Se trabajó con 24 animales sanos mayores de un año de edad. Todos los perros resultaron normales al examen físico y neurológico, y ninguno de ellos tenía antecedentes de enfermedad neurológica. Los registros electroencefalográficos fueron obtenidos con un programa diseñado para electroencefalografía computada y reconstrucción por mapeo cerebral, con 12 canales simultáneos de registro.
Las variables del estudio de la coherencia intrahemisférica permiten la identificación de dos grupos de canales con diferencias en su actividad eléctrica, los canales halo (Fp-T-O) y los canales dorsales (O-P-F-Fp). Los canales dorsales presentan un comportamiento en un rango frecuencia que sitúa su actividad entre la banda theta y la banda alpha (0 7,58 Hz), diferente al de los canales halo que se ubica en la banda theta (0 6,63 Hz). El espectro de fase indica que en esta región la señal se distribuiría desde la corteza temporal hacia rostral y hacia caudal mediante dos sistemas anatómicos diferentes.
Palabras clave: Coherencia cerebral intrahemisférica; Caninos; Electroencefalograma cuantificado; Espectro de potencia; Espectro de coherencia; Espectro de fase.
Analysis of cerebral intrahemispheric coherence in adult dogs.dual dinamic of the cerebral electrical activity
Summary
We investigated the intra-hemispheric coherence in the scalp electroencephalogram of 24 adult dogs. In additional way, we have decreed some hypothesis that acquaintance the results and the anatomic connections involved.
We worked with 24 healthy adult animals, all of them mayors of 1 year old. All dogs were normal at physic and neurological examination, and none had history of neurological illness.
The electroencephalographic record was obtained with a computer electroencephalographic and brain mapping soft, with 12 simultaneous channels of registry.
Halo (Fp-T-O) and Dorsal (O-P-F-Fp) group of electrode was defined by significant statistic difference. The dorsal group showed a performance in a frequency seems to be placed between theta and alpha band (0 7,58 Hz), different to halo group who showed a typical pattern of behavior in the limits of theta band (0 6,63 Hz).
The phase spectrum show that in the temporal region different pattern of anatomical pathways connect anterior and caudal areas.
Keywords: Cerebral intrahemispheric coherence; Dogs; Quantitative electroencephalography; Spectral analysis.
Introducción
La posibilidad de digitalizar y almacenar el trazado electroencefalográfico en soportes magnéticos ha permitido el desarrollo de una serie de herramientas matemáticas para analizarlo posteriormente a su adquisición. Estas técnicas facilitan la remoción de artefactos, el análisis de la fase y las frecuencias, la detección, el análisis y la predicción de crisis epilépticas, y hasta la posibilidad de superponer los datos a las técnicas de neuroimagen. Estas posibilidades permiten a partir de la cuantificación del electroencefalograma (EEG), un mejor conocimiento de la dinámica normal y patológica de la actividad cerebral.
Entre las diferentes variantes de análisis matemático del EEG, el análisis del espectro de potencias por frecuencia es uno de los métodos más utilizados. La frecuencia es el número de veces que un proceso se repite durante un período de tiempo. La teoría del algoritmo de Fourier permite descomponer una señal periódica en un conjunto de ondas, que sumadas entre sí reproducen a la señal original.
El análisis de coherencia permite analizar el EEG desde el punto de vista de sus frecuencias. Se define como la correlación normalizada que existe entre dos pares de canales electroencefalográfico, e informa sobre los componentes frecuenciales comunes a ellos, junto con la magnitud de su asociación. Sus elementos son: el espectro de potencia cruzada (la expresión de las armónicas que tienen en común ambas señales); el espectro de coherencia (que indica la simetría entre las señales); y el espectro de fase (una expresión de la sincronía de las señales)14.
Las funciones de coherencia han sido utilizadas en diversas investigaciones acerca de la generación de las señales del EEG y su relación con las funciones cerebrales, entre las que se destacan los estudios del origen del ritmo Theta hipocampal en estructuras límbicas humanas9,13,76,77, el origen del ritmo alpha córticotalámico34,35,37; el origen de las ondas con frecuencia de 7-14 Hz en estadíos de sueño en humanos13, la diferenciación de los componentes frecuenciales de ritmos estrechamente vecinas entre sí58 y la determinación de las relaciones topográficas entre los diferentes componentes del EEG13 . Muchos autores han utilizado la función de coherencia como herramienta para comparar estados diferentes de actividad cerebral. Se ha empleado este estudio para evaluar la eficiencia neuronal y la posible asociación con la inteligencia en sujetos que realizaban el test de Weschler67. También se la utilizó para evaluar la actividad theta en individuos que realizaban un acto motor voluntario56; otros autores la han empleado como herramienta neurofisiológica para diagnóstico diferencial entre la demencia con Cuerpos de Lewy y la enfermedad de Alzheimer28. Se ha utilizado también el análisis del espectro de fase para estudiar los diferentes ritmos oscilatorios que se observan en los períodos de consolidación de la memoria en humanos63.
En cuanto al uso en la práctica clínica, la función de coherencia en el análisis rutinario del EEG ha sido bastante limitado hasta el momento. Sus aplicaciones más importantes han sido: a) el desarrollo del EEG en bebés57 y en cachorros de perros50,64; en niños con trastornos del neurodesarrollo15 y con síndrome disatencional e hiperactividad16; b) la determinación de asimetrías y/o asincronías intra/interhemisféricas, que son muchas veces los únicos índices de anomalías remanentes cuando no se observan componentes paroxísticos54; c) la evaluación de la propagación de fenómenos epileptógenos a través del cuerpo calloso21.
El objetivo de este trabajo es describir el comportamiento de la actividad cerebral en caninos adultos normales mediante el estudio de la coherencia intrahemisférica, con la finalidad de elaborar una base de datos que permita detectar alteraciones cuantitativas en individuos con distintas enfermedades neurológicas.
Materiales y métodos
Población
Se trabajó con 24 animales sanos mayores de un año de edad (11 machos y 13 hembras) de distintas razas, con edades de 1 a 9 años (media: 4.5 años); la frecuencia y las características de la actividad eléctrica cerebral es comparable en distintas razas de perros, aunque el voltaje puede variar37. Todos los perros resultaron normales al examen físico y neurológico, y ninguno de ellos tenía antecedentes de enfermedad neurológica.
Electrodos
Se utilizaron electrodos de aguja de acero inoxidable de aproximadamente 150 mm de longitud, tanto para los de exploración, como para los de referencia y los de tierra (Akonic S.A., electrodos de aguja subdérmicos)29.
Para registrar los locus temporales (T3 y T4) se emplearon agujas monopolares de electromiografía (EMG) (Akonic S.A., Buenos Aires, Argentina) con cubierta aislante de teflón excepto en su punta50,51.
Equipamiento
Para obtener los registros EEG se utilizó un programa especialmente diseñado para electroencefalografía computada y reconstrucción por mapeo cerebral (AKONIC BIOPC versión modificada 7.0), con 12 canales simultáneos de registro. El sistema permite promediar y procesar muestras múltiples. El equipo incluye 12 canales EEG unidos a una interfase analógico-digital y el sistema de computación.
Los parámetros técnicos del registro fueron los siguientes: Rechazo de modo común > de 100 db; respuesta de frecuencia de 0,5 a 128 Hz; filtro antialiasing: 36 db/octava; filtro de alta frecuencia (FAF): 12 db/octava; filtro de baja frecuencia (FBF): 6 db/octava; ruido propio < 1 uV; frecuencia de muestreo: 256 s; velocidad del papel: 30 mm/s; impedancia de los electrodos: < 5 kΩ.
Restricción de los animales
La restricción de los animales se realizó por medio de inyección subcutánea de xylazina (Rompún®, Bayer Argentina S.A., Buenos Aires, Argentina), basándonos en trabajos previos11,22,26,69 y en nuestra propia experiencia48,50,51, en dosis de 1 mg/kg. La xylazina es un agonista de receptores á2 adrenérgicos11,23,26 que no tiene efectos directos sobre las neuronas corticales piramidales69.
Procedimiento
Terminología y posición de los electrodos de registro. La colocación de los electrodos se realizó de acuerdo a estudios y recomendaciones previas.48,49, 50,52
Registro. El registro básico se efectuó empleando montaje de referencia común, con electrodo de referencia nasal. Esto es indispensable para la cuantificación del EEG, ya que de este modo todos los electrodos tienen una misma referencia, que es utilizada como patrón para comparar el voltaje en los diferentes canales10. El tiempo mínimo de registro fue de 30 minutos.
Análisis matemático
Análisis espectral. Para cuantificar los EEG de cada animal se seleccionaron al menos 15 segmentos estacionarios de 2 segundos deduración libres de artificios (épocas), promediando los resultados de cada época63. Luego de la selección se realizó la estimación espectral aplicando a los segmentos seleccionados la Transformada Rápida de Fourier (FFT [del inglés Fast Fourier Tranform])16.
Análisis de la coherencia
Con los datos obtenidos a partir del espectro de potencia se realizó en cada animal y en cada hemisferio cerebral el análisis de coherencia intrahemisférica para la siguiente configuración de pares de canales: en la posición dorsal se combinaron los electrodos Occipitales-Parietales (O-P); Occipitales- Frontales (O-F) y Occipitales-Frontopolares (O-Fp); a este conjunto lo denominamos "canales dorsales". En la posición lateral se combinaron los electrodos Frontopolares- Temporales (Fp-T) y Temporales-Occipitales. (T-O); a este conjunto lo denominamos "canales halo"50.
Para la obtención de los valores de coherencia se determinaron primero las armónicas comunes a cada par de canales, superponiendo sus espectros de potencia (espectro de potencia cruzada). Se seleccionó entonces la armónica en común de mayor potencia, que corresponde a una determinada frecuencia (Pico de Potencia Cruzada -PPC). A esa frecuencia corresponde también un determinado valor de coherencia propiamente dicha (espectro de coherencia), y un determinado espectro de fase, expresado en grados o, de manera más práctica, en milisegundos. La combinación del número de caninos estudiados x el N° de pares de electrodos analizados genera un total de 240 observaciones (24 perros x 10 pares de canales= 240 observaciones).
Análisis estadístico
En una primera etapa se realizó la estadística descriptiva de los datos con la finalidad de evaluar el comportamiento de las variables; las herramientas gráficas utilizadas (box and whisker plot y rankit plot) indicaron que las distribuciones de las variables en estudio son aproximadamente simétricas. Los valores de will-shapiro tendientes a 1 confirmaron que el análisis paramétrico, basado en la normalidad de los datos, es una herramienta adecuada. En una segunda etapa se realizó análisis de varianza a un solo criterio de clasificación (ANOVA) para los diferentes componentes del estudio de coherencia cerebral; todos los resultados fueron evaluados al 0.05 %. Para la diferencia de medias se realizó la prueba de Tukey con un valor del error á del 0.05 %.
Resultados
No hemos encontrado diferencias significativas entre ambos hemisferios cerebrales para el valor de la frecuencia a la que se presenta el PPC (f: 0.16; p: 0.687), para el espectro de coherencia (f: 0.17; p: 0.716) ni para el espectro de fase para dicho valor de frecuencia (f: 0.29; p: 0.58). (Ver gráfico 1 ángulo superior derecho)
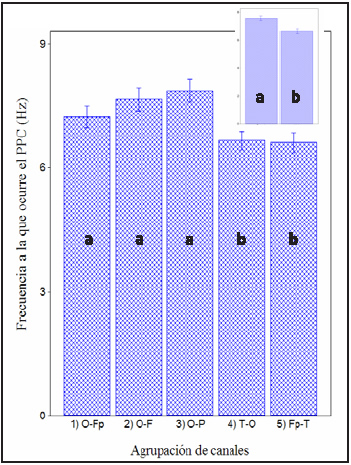
Gráfico 1: El Gráfico de barras muestra los valores de la frecuencia a la que se da la armónica en común de mayor potencia (PPC) para las diferentes agrupaciones de canales. El análisis de varianza muestra diferencia significativa entre los canales dorsales y halo (f: 4,91; p: < 0.001) Las combinaciones de electrodos con media similar se encuentran indicado con letras iguales. Las barras muestran el error estándar de la media. El gráfico del ángulo superior derecho muestra las diferencias que se observan al agrupar los canales en grupo dorsales (a) y grupo halo (b).
Al analizar el PPC, los canales dorsales muestran un valor de frecuencia (0 7,58 Hz ± 0,15) significativamente diferente a los canales halo (6,63 Hz ± 0.16) (f: 4,91; p:< 0.001); el valor de la coherencia también presenta diferencias significativas (f: 21,04; p: < 0,001) entre ambos grupos de canales (0 0,77 ± 1,36 y 0 0,64 ± 2,05 respectivamente). (ver gráfico 1 y 2).
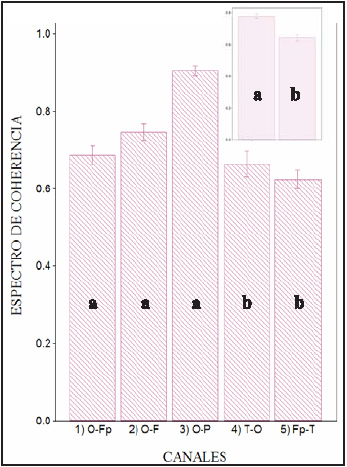
Grafico 2: El Gráfico de barras muestra los valores del espectro de coherencia para los diferentes grupos de canales. El análisis de varianza muestra diferencia significativa entre los canales dorsales y halo, indicado con letras (f:21,04; p:<0,001) Las combinaciones de electrodos con media similar se encuentran indicado con letras iguales. Las barras muestran el error estándar de la media. El gráfico del ángulo superior derecho muestra las diferencias que se observan al agrupar los canales en grupo dorsales (a) y grupo halo.
Con respecto al espectro de fase, existe diferencia en el comportamiento de los canales dorsales y los canales halo (f: 6,00; p: <0.001) (ver gráfico 3A). El valor promedio de los canales dorsales es de 5,10 ms (± 0,88). No hemos hallado diferencias significativas entre ellos (p: 0.079) (ver gráfico 4B). Dentro del grupo de los canales halo existen diferencia significativas entre los canales Fp-T (17,34 ms.) y los canales T-O (8,66 ms.) (p: 0.01) (ver gráfico 3 B). El valor de estos últimos no es diferente al de los canales dorsales (p: <0.001).
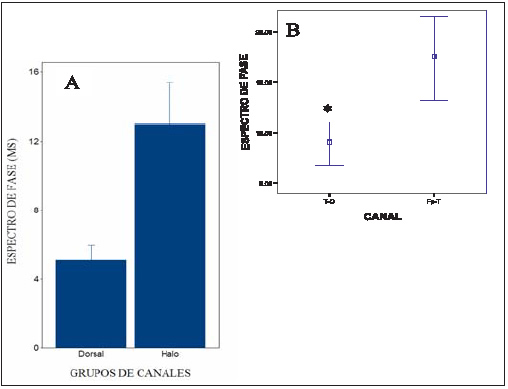
Gráfico 3: A) El grafico muestra las diferencias en el espectro de fase para los grupos de canales Halo y Dorsales. (f:6,00; p:<0.001). B) Comportamiento del espectro de fase en los canales Halo. El asterisco señala diferencias significativas según ANOVA (P: 0,01). Los rombos marcan la media, y las barras el error estándar de la media.
Al analizar el comportamiento del grupo de los canales dorsales, observamos que los electrodos O-P presentan valores de coherencia significativamente mayores (0,90) que los restantes pares de canales (O-F: 0,74; O-Fp: 0.68) (p:< 0.001) (ver gráfico 4 A). Dentro del grupo de los canales halo no se observan diferencias significativas (p: 0.23).
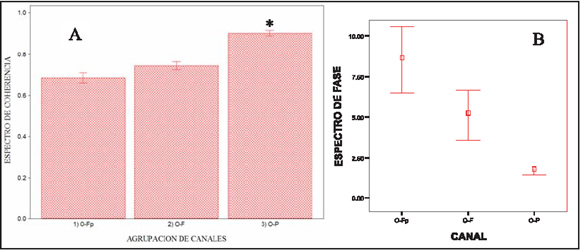
Gráfico 4: comportamiento del espectro de coherencia (A) y del espectro de fase (B) en los canales dorsales. El asterisco señala las diferencias significativas según ANOVA (p:<0.001). Las barras muestran el error estándar de la media. En el espectro de fase, si bien no se observaron diferencias significativas, presenta una clara tendencia a aumentar el retraso en la señal en forma proporcional a la distancia con respecto al electrodo occipital.
Conclusiones
La coherencia expresa la correlación cruzada entre dos canales electroencefalográficos en el dominio de las frecuencias y refleja el número y la fuerza de las conexiones de los generadores neuronales, que a menudo se encuentran a distancia del sitio en donde se obtiene la señal17. Desde el punto de vista matemático, la coherencia expresa la relación entre dos series de tiempo que, a través de la FFT, se analizan en el dominio de las frecuencias17. Desde el punto de vista biológico la coherencia ha sido definida como la tendencia de las señales eléctricas (tales como las del EEG) de aumentar y decaer en sincronía. Se calcula generalmente para un valor de frecuencia determinado, y es medida entre 2 canales59, 62. En forma práctica, se interpreta como una herramienta para medir la similitud entre 2 señales: valores de coherencia por encima de 0.60 indicarían una fuerte semejanza, asociada a la presencia de un generador común para la armónica evaluada. El espectro de fase nos permite conocer qué tan desfasadas (corridas en el tiempo) se hallan las armónicas en común. En general se acepta que tiempos mayores a 15 ms. involucran la participación de estructuras locales que intervienen en la modulación de la señal; tiempos menores indican la existencia de un generador común a los canales estudiados14.
En este estudio no hemos hallado diferencia alguna en los valores de la coherencia intrahemisférica entre ambos hemisferios cerebrales. Este hecho indicaría que en los períodos de baja actividad atencional (como ocurre durante la adquisición del EEG en el perro), ambos hemisferios presentan una dinámica similar en lo que respecta a los generadores neuronales y al sistema de distribución de la señal analizada.
El análisis de la coherencia cerebral intrahemisférica indica la existencia de dos dinámicas diferentes en la actividad cerebral intrahemisférica de los caninos. Los canales que hemos denominado "dorsales" tienen un generador común cuya actividad se encuentra alrededor de los 7,58 Hz, mientras que los canales que hemos denominado "halo" poseen un generador común diferente al anterior, cuya actividad se centra alrededor de los 6,63 Hz.
Es decir, al analizar el comportamiento de la coherencia en las distintas combinaciones de electrodos se detectan 2 grupos claramente diferenciables: los electrodos halo presentan un PPC a una frecuencia 0 de 6,63 Hz (6,30- 6,95) (ver gráfico 1 ángulo superior derecho). Esta combinación de canales monitorea la actividad de la neocorteza de las áreas prefrontales (región dorso-lateral del giro prorreo), temporales (giro silviano y ectosilviano) y occipitales (región dorso-lateral del giro marginal y giro occipital). Los electrodos T registran también actividad evocada de la corteza arquipalial (hipocampo) y paleopalial (lóbulo piriforme)50,52. La coherencia es alta en toda la región (0,62 para los canales Fp-T y 0.66 para los canales TO), lo que sugiere la presencia de un generador común para la actividad eléctrica a la que se presenta el PPC en la región en cuestión. (ver tabla 1)
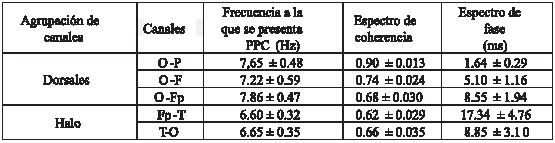
Tabla 1: La tabla muestra un resumen de los valores obtenidos para el estudio de coherencia cerebral intrahemisférico. En ella pueden observarse el valor de la media y el error estándar de la media al que se presenta el PPC, junto con el valor del espectro de coherencia y de fase para cada canal.
El grupo de electrodos dorsales presenta un valor de PPC entre 7,22 y 7,86 Hz (0 7,54Hz). (ver gráfico 1 y tabla 1) Los electrodos dorsales monitorean la actividad de las áreas dorsales de los hemisferios cerebrales. En correspondencia con la posición de los electrodos, las principales áreas de proyección son la región dorso-lateral del giro prorreo (electrodos Fp), el giro precruzado (electrodo F), la región dorso-lateral del giro ectomarginal (electrodos P), el giro marginal y la región dorso-lateral del occipital (electrodos O)50-52. Los valores de coherencia de los canales dorsales se hallan en todos los casos por arriba de 0,6, observándose una tendencia a disminuir en forma caudo-rostral (O-P: 0,90; O-F: 0,74; O-Fp: 0,68). Estos valores sugieren la existencia de un generador común para la armónica a la que se presenta el PPC de esta región.
Respecto al espectro de fase, la diferencia en los valores entre los electrodos del grupo de canales halo (17,34 mseg para los canales Fp-T y 8,42 mseg para los canales T-O) (ver gráfico 3 B) sugiere que la distribución de la señal se realiza mediante vías anatómicas distintas hacia rostral y hacia caudal, a partir del área de proyección de los electrodos T. en cuanto al comportamiento del espectro de fase para los canales dorsales se observa una tendencia a aumentar el tiempo de llegada de la señal a los electrodos a medida que se alejan respectivamente del electrodo occipital (O-P: 1,64 ms; O-F: 5,10 ms; O-Fp: 8,55 ms).
Discusión
En cuanto a los valores que hemos hallado para los canales halo, trabajos realizados en diferentes especies han demostrado que el ritmo característico de la corteza hipocampal y de otras áreas corticales límbicas (corteza cingular, corteza entorrinal, corteza del giro dentado y complejo subicular) se encuentra entre los 4-7 Hz, y lo han denominado ritmo theta hipocampal o actividad rítmica lenta (RSA -rhythmic slow activity-)1,2,3,25,39,78. Teniendo en cuenta que en nuestra experiencia, los electrodos temporales en caninos no sólo permiten monitorear información proveniente de la neocorteza del área temporal, sino que también registran la actividad evocada de ciertas regiones del arquipalio (hipocampo) y del paleopalio (lóbulo piriforme) 50, por lo tanto es esperable que la armónica dominante en la agrupación de canales halo se ubique dentro del rango de la actividad theta.
Los dipolos generadores de este ritmo se encuentran en el hipocampo (neuronas piramidales de CA1) y en las neuronas granulares del giro dentado, aunque su normal presentación depende de la integridad de la porción septal del rinencefalo, que actúa como marcapaso del SRA, conectándose con los generadores mediante un sistema de fibras originadas en el núcleo diagonal y en el núcleo septal medial 1,24,39. Este marcapaso se encuentra modulado por una vía retículoseptal originada en los núcleos reticulares pontinos (particularmente el núcleo pontino oral 72 y en los núcleos del rafe medio (vía serotoninérgica) 5,40,70,72 que actúan sobre los núcleos septales directa o indirectamente, a través del núcleo supramamilar 71. Este sistema modula el SRA, sincronizándolo o desincronizándolo, influenciado por el estado de vigilia y motivación. En una revisión reciente, Pam and McNaughton concluyen que la actividad theta no podría generarse normalmente sin la integración de la intensa actividad ascendente proveniente de la sustancia reticular. En la actualidad existe evidencia suficiente como para afirmar que el ritmo theta es controlado por una extensa red neuronal que se extiende por el tronco encefálico hasta el diencéfalo, y desde allí asciende a través de la región septal hasta alcanzar el hipocampo 45.
Las conexiones anatómicas entre el hipocampo y la corteza prefrontal han sido estudiadas por diferentes autores desde Papez (1937) hasta la actualidad 39,46,47,61,73,75. Recientemente se ha sugerido, en base al acoplamiento de fase, la existencia de asociaciones monosinápticas entre el hipocampo y las neuronas piramidales de la corteza prefrontal. Los valores comunicados en referencia al desfasaje de la información (alrededor de 20 ms) son semejantes a los que nosotros hemos encontrado entre los canales Fp y T (17,34 ms) 18,27,65,68 (Ver tabla 1).
En relación a las conexiones témporo-occipitales, desde hace mucho se conoce la existencia del fascículo uncinado, uno los tres fascículos de fibras largas de asociación que relacionan distintas áreas corticales de los hemisferios cerebrales 6,43. A su vez, el área de la corteza visual no sólo se encuentra conectada a la corteza retroesplenial del giro del cíngulo 39, sino también a la corteza del complejo subicular. En muchas especies se han descripto un gran número de conexiones recíprocas (output-inputs) entre el circuito hipocampo-entorinal y las áreas sensoriales de la corteza, entre ellas la corteza de la región occipital. Dichas conexiones también podrían participar en la diseminación de la señal 39. Estas vías, teniendo en cuenta los valores del espectro de fase, son más rápidas que las que se dirigen a la corteza órbitofrontal.
Respecto al valor de frecuencia en donde se halla en PPC para los canales dorsales (7,58 Hz); este valor, desde el punto de vista matemático, corresponde a 7 u 8 Hz, ya que las bandas electroencefalográficas se encuentran definidas para valores enteros de frecuencia. Este fenómeno, como hemos sugerido en trabajos anteriores 75 podría interpretarse de dos maneras: a) que se encuentra en el límite inferior de la banda alpha o en el límite superior de la banda theta; b) que en la neocorteza dorsal coexisten en forma simultánea ambos ritmos sin que ninguno de ellos se constituya en la frecuencia dominante. Hay suficiente evidencia científica como para sustentar ambos postulados. Lopes Da Silva y col. han comunicado que el ritmo alpha se genera en la corteza de la región occipital 34,37,38,66. Esta señal estaría producida por un proceso estocástico u aleatorio, que aparece como el resultado de propiedades de filtro de redes neuronales neocorticales ("filtro alpha").
Por otra parte, hallazgos en humanos obtenidos con electrodos intracraneanos indicarían que la actividad theta neocortical no sólo es producto del efecto del volumen de conducción, sino que difunde en la corteza a través de conexiones intracerebrales, e incluso se ha observado que puede tener generadores neocorticales propios - por ejemplo durante el sueño REM- en la región del giro parahipocampal 8.
Los valores del espectro de coherencia y del espectro de fase hallados para las diferentes combinaciones de canales dorsales (ver gráfico 4 A y B) sugieren la existencia de un generador común para la armónica a la que se presenta el PPC de esta región, y la existencia de una red de conexiones corticales compuesta por los sistemas cortos y largos de fibras de asociación (tracto occípito-frontal y tracto cingular) que conforman la sustancia blanca subcortical, que actuaría homogeneizando la actividad intrahemisférica en dichas áreas 43.
La llamativa similitud de la actividad registrada entre los electrodos occipitales y parietales en ambos hemisferios, sumado a su elevada sincronía, podría deberse a la cercanía de las áreas registradas y al elevado número de conexiones intracorticales (fibras de asociación cortas) que existen entre las áreas sensoriales primarias y las áreas corticales de asociación 43.
La elaboración de una base de datos con caninos adultos normales permitirá estandarización de los valores de coherencia cerebral, esto permitirá realizar comparaciones en diferentes estados patológicos.
Finalmente queremos remarcar al momento de analizar el EEGc de un canino que, en base a nuestros hallazgos, debería tenerse en cuenta que según la combinación de electrodos elegidos la frecuencia a la que se presenta el PPC varía. Si se desea analizar alguna de las combinaciones de los canales dorsales, es de esperar que el pico de potencia cruzado se halle entre valores de frecuencia 7,27-7,89 Hz, con elevados valores de coherencia (superiores a 0,60) y tiempos 5,10 ms (± 0,88).
Si se desea estudiar el comportamiento de los electrodos que pertenecen al grupo halo (Fp-T y T-O) es de esperar que los valores de la pfrecuencia a la que se presenta el PPC se encuentren entre 6,03- 6,95 Hz, con valores de coherencia cercanos a 0,60 y tiempos mayores el espectro de frecuencia de los canales Fp-T (17,34 ± 4,76) que para los canales T-O (8,85 ± 3,10). (ver tabla 1)
1. Arnolds, D.E; Lopes da Silva, F.H; Aitink, J.W; Kamp, A. Hippocampal EEG and behaviour in dog. II. Hippocampal EEG correlates with elementary motor acts. Electroencephalogr Clin Neurophysiol. 1979 May; 46(5):571-80. [ Links ]
2. Arnolds, D.E; Lopes da Silva, F.H; Aitink, J.W; Kamp, A. Hippocampal EEG and behaviour in dog I. Hippocampal EEG correlates of gross motor behaviour. Electroencephalogr Clin Neurophysiol. 1979 May;46(5):552-70. [ Links ]
3. Arnolds, D.E; Lopes da Silva, F.H; Aitink, J.W; Kamp, A. Hippocampal EEG and behaviour in dog. III. Hippocampal EEG correlates of stimulus-response tasks and of sexual behaviour. Electroencephalogr Clin Neurophysiol. 1979 May; 46(5):581-91. [ Links ]
4. Artemenko, D.P; Role of the hippocampal neurons in the theta -wave generation. Neurophisiology 1978 4:531-539. [ Links ]
5. Assaf, SY, Miller JJ. Role of a raphe serotonin system in control of septal unit activity and hippocampal desynchronization. Neuroscience 1978. 3:539-550. [ Links ]
6. Barone, R.; Ensayo de Anatomía funcional y comparada del Sistema nervioso Central. Universidad Nacional de Río Cuarto. 1989. [ Links ]
7. Bland, B. M. and Whisha, I. Q.; Generators and topography of hippocampal theta (SRA) in the anaesthetized and freely moving rat. Brain research. 118: 259-280, 1976. [ Links ]
8. Bódizs R, Kántor S, Szabó G, Szütilde A, Erõss L, Halász P, Rhythmic hippocampal slow oscillation characterizes REM sleep in humans. Hippocampus 2001, Vol 11 6: 747- 753. [ Links ]
9. Brazier, M.A.B. "A History of the Electrical Activity of the Brain. The first half century". Pitman, London. 1961 [ Links ]
10. Casaglia, D.C. La adquisición de datos para electroencefalografía computada. En "Análisis computado del EEG". Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. 1995. 54-76. [ Links ]
11. Celly, C.S., McDonnel, W.N., Young, S.S., Black, W.D. The comparative hypoxemic effect of four a2 adrenoceptor agonists (xylazine, romifidine, detomidine and medetomidine) in sheep. J.Vet.Pharmacol. Therap. 1997. 20: 464-471. [ Links ]
12. Charles Leclerc, Sidney J. Segalowitz, James Desjardins, Maryse Lassonde, Franco Lepore. EEG coherence in early-blind humans during sound localization; Neuroscience Letters 2005. 376: 154- 159. [ Links ]
13. Dumermuth, G.; Walz, W.; Scollo-Lavizzari, G. And Kleiner, B.; Spectral analysis of EEG activity during Sleep Stages in Normal adults. Eurpo. Neurology; 1972. 7: 265-296. [ Links ]
14. Etchepareborda, M. "El análisis de la coherencia interhemisférica". En "Análisis computado del EEG". Cáp. 6 Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 83-91. [ Links ]
15. Etchepareborda, M.; Etchepareborda, J.; González, S. "El análisis de la coherencia interhemisférica en pacientes con disfasia infantil. Presentación de subtipos". Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina. 1992. [ Links ]
16. Etchepareborda, M.; Etchepareborda, J.; González, S. "Relación entre el espectro de coherencia de la actividad bioeléctrica cerebral y los hallazgos neurofisiológicos en pacientes con síndrome disatencional e hiperactividad". Rev. Neurol. Barcelona. 1994. 22 (115): 266-271. [ Links ]
17. Evans J. R.; Abarbanel A. Quantitative EEG and Neurofeedback. Ed Academic Press. San Diego, California.U.S.A. 1999. ISBN 0122437990X. Ch 1 "An Overview of Quantitative EEG and Its Applications to Neurofeedback". [ Links ]
18. Ferino, F., Thierry, A., and Glowinski, J. Anatomical and electrophysiological evidence for a direct projection from Ammon's horn to medial prefrontal cortex. Exp. Brain Res. 1987 65, 421-426. [ Links ]
19. Ferrero, R.G.A.; Ferrero, A.L. "Análisis computado del EEG". Editado por Ferrero, R.G.A.; Ferrero, A.L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As., Argentina. 1995. [ Links ]
20. Gotman, J. Análisis del EEG asistido por computadora (Monitoreo automático de espigas). En "Análisis computado del EEG". Cáp. 21. Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As., Argentina. 1995. Sección V. 241-257. [ Links ]
21. Gotman, J. Noninvasive method for evaluating the localization and propagation of epileptic activity. Epilepsy 2003, 44 supl 12: 21-29. [ Links ]
22. Green, J.D. and Arduini, A. Hippocampal electrical activity in arousal J. Neurophysiol.1954; 17: 533-557. [ Links ]
23.Greene, S.A., Thurmon, J.C.; Xylazine, A review of its pharmacology and use in veterinary medicine. J. Vet. Pharmacol. Therap. 1988. 11: 295-313. [ Links ]
24. Holmes, J. E. and Adey W. R.; Electrical activity of the entorhinal cortex during conditioned behaviour. Am. J. Physiol. 1960 Nov; 199: 741-744. [ Links ]
25. Holsheimer J, Stok C.J, Lopes da Silva F. H. Theta rhythm related hippocampal cell discharges in the urethane anaesthetized rat: evidence for a predominant entorhinal input. Electroencephalogr. Clin. Neurophysiol. 1983 Apr; 55(4):464- 467. [ Links ]
26. Jarvis, N., England, G.C.W. Reversal of Xylazine sedation in Dogs. The Veterinary Record, April 1991. 128: 323-325. [ Links ]
27. Jay, T., Thierry, A., Wiklund, L., and Glowinski, J. Excitatory amino acid pathway from the hippocampus to the prefrontal cortex. Contribution of AMPA receptors in hippocampo-prefrontal cortex transmission. Eur. J. Neurosci. 1992. 4: 1285-1295. [ Links ]
28. Kai T., Asai Y., Sakuma K., Koeda T., Nakashima K. Quantitative electroencephalogram analysis in dementia with Lewy bodies and Alzheimer's disease Journal of the Neurological Sciences 2005, 237: 89-95. [ Links ]
29. Kuijper, T. La importancia de los electrodos para EEG. En "Análisis computado del EEG". Cáp. 38 Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 424- 435. [ Links ]
30. Landfield, P. W. (1977). Different effects of postrial driving or blocking of the theta rhythm on avoidance learning in rats. Physiology and Behaviour, 18, 439- 45. [ Links ]
31. Landfield, P. W.; McGaugh, J. L.; Tusa, R. J. Theta rhythm: a temporal correlate of memory storage processes in the rat. Science. 1972 Jan 7; 175(17):87-9. [ Links ]
32. Lanfield, P. W. Computer-determined EEG patterns associated with memory-facilitating drugs and with ECS. Brain. Res. Bull. 1976 Jan-Feb; 1(1):9-17. [ Links ]
33. Le Moal, M.; Cardo, B. Rhythmic slow wave activity recorded in the ventral mesencephalic tegmentum in the rat. Electroencephalogr. Clin. Neurophysiol. 1975 Feb;38(2):139- 147. [ Links ]
34. Lopes da Silva, F. H.; Van Lierop, M. T.; Schrijer, C. F.; Storm van Leeuwen, W.; Organization of the thalamic and cortical alpha rhythms: spectra and coherence. Electroencephalography Clinical Neurophysiology. 1973; 35:627-639. [ Links ]
35. Lopes da Silva, F.H, Kamp, A. Hippocampal theta frequency shifts and operant behaviour. Electroencephalogr. Clin. Neurophysiol. 1969 Feb; 26(2):133-43. [ Links ]
36. Lopes da Silva, F.H., Storm van Leeuwen, W. The cortical source of alpha rhytm. Neurosci. Lett. 1977. 6: 237-241. [ Links ]
37. Lopes da Silva, F.H; EEG Analysis: theory and Practice. En "Electroencephalography". Cap 61. Edit by Niedermayer, E.; Lopes da Silva, F. William y Wilkins. Philadelphia. Third edition. 1993. 1097-1123. [ Links ]
38. Lopes da Silva, F.H; Vos, J.E; Mooibroek, J; Van Rotterdam, A.; Relative contributions of intracortical and thalamo-cortical processes in the generation of alpha rhythms, revealed by partial coherence analysis. Electroencephalography Clin. Neurophysiol. 1980 Dec; 50(5-6):449-56. [ Links ]
39. Lopes da Silva, F.H; Witter, M.P; Boeijinga, P. H; Lohman, A. H; Anatomic organization and Phisiology of the Limbic Cortex. Physiol. Rev. 1990, Apr. 17(2) 453-511. [ Links ]
40. Macadar AW, Chalupa LM, Lindsley DB.. Differentiation of brain stem loci which affect hippocampal and neocortical electrical activity. Exp Neurol 1974. 43:499-514 [ Links ]
41. Murer, M.G.; Riquelme, L.A.; Generalidades. En "Análisis computado del EEG". Cáp. 3. Editado por Ferrero, R.G.A.; Ferrero, A.L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 41-53. [ Links ]
42. Niedermayer, E.; Historical Aspects. En "Electroencephalography ". Cap 1. Edit by Niedermayer, E.; Lopes da Silva, F. William y Wilkins. Philadelphia. Third edition. 1993. 1-14. [ Links ]
43. Nieuwenhuys R.; Ten Donkelaar H. J., Nicholson Ch. The central nervous system of the vertebrates. Ed Springer. Berlin. 1998, Vol III, Chapter 22, "Telencephalo". 1871-2023. ISBN 3-540-56013-0. [ Links ]
44. Nuwer, M. R.; El análisis cuantitativo del EEG en la clínica. En "Análisis computado del EEG". Cap 28, Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 323-341. [ Links ]
45. Pan WX, McNaughton N. The supramammillary area: its organization, functions and relationship to the hippocampus. Prog Neurobiol 2004. 74:127-166. [ Links ]
46. Pandya D.N.; Seltzer B. Intrinsic connections and architectonics of posterior parietal cortex in rhesus monkey. J. Comp. Neurol. 1982, 204: 196-210 [ Links ]
47. Papez, J.W., 1937. A proposed mechanism of emotion. Arch. Neurol. Psychiatr. 38, 725-734. [ Links ]
48. Pellegrino, F. C., Epilepsia y Síndromes Epilépticos III. Diagnóstico, Pronóstico y Tratamiento de la Epilepsia y los Síndromes Epilépticos. Selecciones Veterinarias, 1999. Vol.7. Nº 6: 686-704. [ Links ]
49. Pellegrino, F. C.; "Epilepsia y Síndromes epilépticos". En "El Libro de Neurología para la Práctica Clínica", ED. por Pellegrino, F. C.; Suraniti, A.; Garibaldi, L.; parte III, Cap. 17. Editorial Intermédica. Bs. As. Argentina. 2003. pp. 255-280. [ Links ]
50. Pellegrino, F. C.; Estandarización de los patrones electroencefalográficos de los caninos. Tesis doctoral. U.B.A. 2000. [ Links ]
51. Pellegrino, F. C.; Sica, R; Canine electroencephalographic recording technique: findings in normal and epileptic dogs. Clinical Neurophysiology 115 (2004) pp. 477- 487 [ Links ]
52. Pellegrino, F.; Etchepareborda, M.; Anatomical sites for the placement of electroencephalographic record electrodes in canines. Braz. J. Morphol. Sci. 1997.Vol.14, 2: 197-204. [ Links ]
53. Pellegrino, F.; Etchepareborda, M.; Registro electroencefalográfico en caninos con epilepsia rinencefálica. Selecciones Veterinarias. 1996. Vol. 5, Nº 3: 230-239 [ Links ]
54. Pellegrino, F.C., Epilepsia y Síndromes Epilépticos I. Marco conceptual. Mecanismos excitotóxicos y epileptogénicos. Selecciones Veterinarias, 1999. Vol.7. Nº 4: 431-439. [ Links ]
55. Petsche, H. C. and Stumpf, C.; Topographic and toposcopie study of origin and spread of the regular sychonized arousal pattern in the rabit. Electroencphalographic clinical neurophysiology. 12: 589-600; 1960. [ Links ]
56. Portella C. E., Guilherme Silva J, Machado S., Velasques B., Vale Bastos V. H., Ferreira-Vorkapic C., Terra P., Lopes V., Cagy M., Carvalho E., Orsini M., Basile L., Piedade R., Ribeiro P. EEG Spectral coherencia inter and intrahemisfepheric during catching object fall task. Arq Neuropsiquiatr 2007;65(1):63-67. [ Links ]
57. Prechtl, H.F.R. and Vos, J.E.; Verlaufsmuster der Ferquenzepektren and Kohärenzen bei schalafenden normalen und neurologisch abnormalen Neugeborenen. In Die Quantifizierung des Elektroenzephalogramms. Ed., G.K. Schenk, 1973, pp 167-188. Konstanz: AEG- Telefunken. [ Links ]
58. Ribak, C.E., Bradburne, M., Harris, A.B. A preferential loss of GABAergic, symmetric synapses in epileptic foci: a quantitative ultraestructural analysis of monkey neocortex. J. Neurosci. 1982. 2: 1725-1735. [ Links ]
59. Riquelme, L. Bases neurofisiológicas de las señales bioeléctricas cerebrales. En "Análisis computado del EEG". Cap 3. Editado por Ferrero, R. G. A.; Ferrero, A. L. FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 32-40. [ Links ]
60. Robinson, T.E; Hippocampal rhythmic slow activity (RSA; theta): a critical analysis of selected studies and discussion of possible species-differences. Brain Res. 1980 May; 203(1):69-101. [ Links ]
61. Room P., Russchen F. T., Groenewegen H. J., Lohman A. H. M.. Efferent connections of the prelimbic (area 32) and the infralimbic (area 25) cortices: An anterograde tracing study in the cat. J. Comp. Neurol. 1985. 242: 40-55 [ Links ]
62. Roy John, E. Prichep, L.S. los principios del análisis neurométrico en EEG y potenciales evocados. En "Análisis computado del EEG". Cap 8. Editado por Ferrero, R. G. A.; Ferrero, A. L. Editor FADEC. Laboratorio para el estudio de las funciones cerebrales superiores. Bs. As. Argentina.1995. 111-135. [ Links ]
63. Schack B., Vath N., Petsche H., Geissler H.-G., Möller E. aPhase-coupling of theta-gamma EEG rhythms during shortterm memory processing. International Journal of Psychophysiology2002, 44:143-163. [ Links ]
64. Senba, H.; Tokuriki, M.; Sasaki, N.; Takeuchi, A.; Usui, K. Developmental EEG of the Beagle dogs. Japanese Journal of Veterinary Science. 1984. 46 (6), pp.771-781. [ Links ]
65. Siapas A.G.; Lubenov E.V., and Wilson M.A. Prefrontal Phase Locking to Hippocampal Theta Oscillations. Neuron 2005, Vol. 46, 141-151. [ Links ]
66 Steriade, Mircea. Cellular substrate of Brain rhytms. In "Electroencephalography". Cap 3. Edit by Niedermayer, E.; Lopes da Silva, F. William y Wilkins. Philadelphia. Third edition. 1993. 27-62. [ Links ]
67. Thatcher R.W., North D., Biver C. EEG and intelligence: Relations between EEG coherence, EEG phase delay and power. Clinical Neurophysiology 2005 116, 2129-2141. [ Links ]
68. Tierney, Patrick L., De'gene' tais, Eric, Thierry Anne-Marie, Glowinski, Jacques and Gioanni, Yves. Influence of the hippocampus on Interneurons of the rat prefrontal cortex. European Journal of Neuroscience 2004. Vol. 20, pp. 514-524. [ Links ]
69. Tourai, K.; Senba, H.; Sasaki, N.; Tokuriki, M.; Ohashi, F.; Takeuchi, A.; Usui, K. Developmental EEG of the Beagle dog under xylazine sedation. Japanese Journal of Veterinary Science. 1985. 77 (3): 459-463. [ Links ]
70. Vertes R.P. An analysis of ascending brain stem systems involved in hippocampal synchronization and desynchronization. J Neurophysiol 1981. 46:1140-1159. [ Links ]
71. Vertes R.P., Albo Z. and Viana Di Prisco. Theta-rhytmically firing neurons in the anterior thalamus: implication for mnemonic functions of Papez's circuit. Neuroscience 2001. Vol. 104, No. 3, pp. 619-625. [ Links ]
72. Vertes Robert P. Hippocampal Theta Rhythm: A Tag for Short- Term Memory. Hippocampus 2005. 15:923-935. [ Links ]
73. Vertes, Robert P., Hoover, Walter B., Viana Di Prisco, Gonzalo. Theta Rhythm of the Hippocampus: Subcortical Control and Functional Significance Behavioral and Cognitive Neuroscience Reviews 2004. Volume 3 Number 3, 173-200. [ Links ]
74. Vidal Figueredo, R., Pellegrino, F. Riquelme, L. Analisis y estandarización de los valores de coherencia cerebral interhemisférica en caninos adultos. InVet 2007; Volumen 9, número 1; xxx ISSN (soporte papel) 1514-6634 ISSN (on line) 1668-3498. [ Links ]
75. Vogt B. A., Pandya D. N. Cingulate cortex of the rhesus monkey: II. Cortical afferents. J. Comp. Neurol. 1987. 262: 271-289. [ Links ]
76. Walter, D.O., Adey, W.R. Analysis of brain wave generation as multiple statistical time series. IEEE Trans. Biomed. Eng. 1965. 12: 8-13. [ Links ]
77. Walter, D.O., Adey, W.R. spectral analysis of electroencephalograms recorded during learning in the cat. Exp. Neurol. 1965 8: 155-181. [ Links ]
78. Winson, J., Patterns of hippocampal tetha rhythm in the freely moving rat. Electroencephalographic clinical neurophysiology. 36: 291- 301, 1974. [ Links ]
79. Yamaguchy, Y.; Yoshii, N.; Miyamoto, K. And Itoigawa, N. A study on the invasive q-waves to the cortex. En Adey, W. R. And Tokizane, T. (Eds.), Structure and function of the Limbic System. Progress in Brain Research. Vol 27, Elsevier, Amsterdam 1967, pp. 357-375. [ Links ]

