










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO  uBio
uBio
Compartir

 Permalink
PermalinkInVet
versión On-line ISSN 1668-3498
InVet vol.12 no.1 Ciudad Autónoma de Buenos Aires ene./jun. 2010
ARTÍCULO DE INVESTIGACIÓN
Dinamica folicular y momento de la ovulación en cerdas púberes y pluríparas posdestete
Williams, S.I.1; Fernandez, V.C.; De La Sota, R.L.
1Instituto de Teriogenología, Departamento de Producción Animal, Facultad de Ciencias Veterinarias, Universidad Nacional de La Plata, CC 296, La Plata (1900), Argentina. E-mail: swilliams@fcv.unlp.edu.ar
Recibido: 12/08/09
Aceptado: 26/08/10
Resumen
El proceso del desarrollo folicular en la cerda comprende el reclutamiento y la selección, y no se describen ondas foliculares, a excepción del período prepuberal. El estudio del estado ovárico y de la dinámica folicular mediante el uso de la ultrasonografía es uno de los avances más recientes en ginecología. El objetivo del presente trabajo fue estudiar el número y el tamaño de folículos, la dinámica folicular desde el comienzo del estro hasta la ovulación, en cerdas nulíparas púberes y en pluríparas luego del destete. Las ecografías fueron realizadas por la vía transabdominal (nulíparas) y transrectal (pluríparas), para registrar el número, la localización y el tamaño de los folículos > 3 mm y confeccionar mapas ováricos. Desde el momento que mostraron síntomas de estro (0 h) hasta el momento de la ovulación, las ecografías fueron realizados cada 8 horas. Se observó una disminución en el número de folículos de 3-5 mm, probablemente debido a atresia y a que algunos folículos desarrollaron hasta 6-7 mm; y una disminución del número de folículos de 6-7 mm, debido a que dichos folículos crecieron hasta 8-9 mm. Considerando las diferencias observadas entre cerdas nulíparas y posdestete en el momento de la ovulación (36,5±1,8 vs. 29,1±2,6 h) y en el tamaño de los folículos ovulatorios (5,6±0,5 vs. 7,0±1,0 mm;), se puede concluir que la aplicación de la ultrasonografía adquiere importancia para el seguimiento del desarrollo folicular y para predecir el momento de la ovulación y así, utilizar esta información en el diseño de protocolos para inseminación artificial a tiempo fijo.
Palabras claves: Dinámica folicular; Ovulación; Ultrasonografía; Cerda.
Follicular dynamics and time of ovulation in gilts and postweaning sows
Summary
Follicular development in sows, includes the process of recruitment and selection; however, follicle waves were described only in prepubertal period. Recent technical advances have made possible to study ovarian activity in pig using real time ultrasonography. The objective of this study was to determine the number and follicles size at estrus and the follicular development pattern till the ovulation time in gilts and weaning sows. From the onset of estrus onward, ovaries were scanned every 8 h and number, size, location of ovarian follicles and ovulation time were recorded Ovaries were visualized by transabdominal (gilts) and transrectal (sows) scanning. For transrectal scanning, a sector transducer was attached to a polyvinyl chloride fixed-angle transrectal stabilizing cord. Ovulation time was defined as the time of the first scanning when no presumptive ovulatory follicles were found minus 4 h, and ovulation was confirmed by a further scanning 8 h later. The number of follicles 3-5 mm decreased from the onset of oestrus onward, perhaps, because some of them became atretic and because others moved to the pool of follicles 6-7 mm. The number of follicles medium size (6-7 mm) increased slowly at the onset of estrus, then plateaued, and finally decreased, because they moved to a the pool of follicles 8-9mm, and remained till time of ovulation. Despite differences between gilts and sows in ovulation time (36.5±1.8 vs 29.1±2.6 h, respectively) and preovulatory follicles size (5.6±0.5 vs 7.0±1.0 mm, respectively), it was concluded that ultrasonography is very useful to study follicular dynamics from the onset of estrus to ovulation and thus to use this information to implement fixed-time artificial insemination protocols.
Key words: Folicular dynamics; Ovulation time; Ultrasound; Gilts; Sows.
Introducción
Durante el desarrollo folicular del ciclo estral, tres características parecen ser las más importantes para todas las especies: 1) la secuencia de los eventos (reclutamiento, selección y dominancia); 2) la necesidad secuencial de gonadotrofi nas (FSH para el reclutamiento, LH para la dominancia) y 3) la amplia variedad de requerimientos (número de ondas por ciclo, número de folículos por onda), así como requerimientos temporales (momento de selección y duración de la dominancia). Cuando se compara el patrón del desarrollo folicular entre las especies animales, en la bovina, ovina y equina se describen ondas foliculares, mientras que en la porcina sólo se observan durante el período prepuberal7.
El proceso del desarrollo folicular en la cerda, comprende dos de los procesos mencionados anteriormente: el reclutamiento y la selección. El término "reclutamiento" se refiere a la población de folículos pequeños y medianos presentes en la superficie del ovario que pueden ser seleccionados como folículos ovulatorios, mientras que, "selección", se refiere a aquellos folículos que escapan a la atresia y ovulan9. Una vez que los folículos maduran a más de 1 mm comienzan a ser visibles en la superficie del ovario. En cerdas nulíparas (prepúberes y púberes) y en cerdas posdestete, hay un pool de aproximadamente 50 folículos de 1-6 mm en la superficie del ovario9.
Los ovarios de la hembra porcina, se caracterizan por el gran número de folículos, comparado con otras especies8. Durante la fase luteal, pueden observarse 30-90 folículos de 1-2 mm, y 30-50 folículos de 2-7 mm9; 5; 11. Durante la fase folicular, el número de folículos pequeños y medianos disminuye dramáticamente, dejando un total de aproximadamente 20 folículos, en su mayoría folículos ovulatorios 11 . Los folículos ovulatorios alcanzan 7-10 mm previo a la ovulación3; 6. En las hembras porcinas, debido a que existe un desarrollo coordinado al comienzo de la fase luteal11, hay un crecimiento continuo y atresia de los folículos ováricos durante el resto de la fase luteal (días 7 a 15 del ciclo estral), sin evidencia de ondas ni de dominancia folicular5; 6; 10 y sin estar asociado a cambios en las concentraciones plasmáticas de FSH10. El cuerpo lúteo de la cerda produce varias hormonas, entre ellas inhibina y estradiol, que regulan negativamente la FSH y mantienen las concentraciones en niveles por debajo de los necesarios para el reclutamiento folicular; esto podría explicar la ausencia de ondas foliculares durante la fase luteal en la cerda7. Los folículos que ovularán incrementan su crecimiento entre los días 14-16 del ciclo estral4.
Los folículos ovulatorios son reclutados del pool de folículos antrales, que se desarrollan durante la fase luteal del ciclo, en el cual se observan folículos de aproximadamente 5 mm. Hacia el final de la fase luteal, los niveles de progesterona caen y los niveles crecientes de LH y FSH del sistema hipotálamo-hipófisis producen el reclutamiento de estos folículos. Los folículos pequeños no reaccionan ante este reclutamiento por carecer de los suficientes receptores de LH y de FSH, y terminan en atresia21.
Durante la dominancia folicular, ocurre el crecimiento y la maduración folicular. Existe una relación directa entre la presencia de folículos dominantes y la ausencia de reclutamiento, ya que la cauterización de los folículos dominantes induce el reclutamiento. La magnitud de la dominancia generalmente se define por la diferencia de tamaño entre los folículos dominantes y los subordinados, siendo esta diferencia para la cerda de apenas 3 a 4 mm entre los dominantes y los subordinados7.
El estudio del estado ovárico y de la dinámica folicular mediante el uso de la ultrasonografía, es uno de los avances más recientes en ginecología. El desarrollo folicular y el momento de ovulación han sido estudiados por Weitze et al. (1989), utilizando la ultrasonografía transabdominal. Soede et al (1992) demostraron que, la vía transrectal resultó ser más apropiada para el estudio de la dinámica folicular, y que permite estudiar en tiempo real el desarrollo folicular y el momento de la ovulación sin interferir en los procesos estudiados23; 14; 15; 16.
La identificación de estructuras ováricas por medio de la ultrasonografía en cualquier momento del ciclo y durante la gestación, es útil para controlar el período periovulatorio o para verificar la actividad ovárica, en caso de fallas reproductivas. Las aplicaciones podrían ser: 1) como rutina en granjas sin problemas reproductivos, para comprobar la eficiencia de ciertos manejos; 2) como profilaxis o prevención de fallas reproductivas (por ejemplo, el uso de la sincronización hormonal de celo o el acortamiento del período de lactancia) y 3) en casos de repentina disminución de la eficiencia reproductiva, por repeticiones cíclicas o bien fallas en la detección de celo13.
El objetivo del presente trabajo fue estudiar, por medio de la ultrasonografía, el número y el tamaño de folículos, la dinámica folicular desde el comienzo del estro hasta la ovulación en cerdas nulíparas púberes y en pluríparas luego del destete.
Materiales y métodos
Cerdas nulíparas
Se utilizaron hembras púberes, de genética definida. Las cerdas (n=7) estaban alojadas en corrales individuales (instalaciones ad-hoc, de la FCV, UNLP), alimentadas con una fórmula comercial para hembras gestantes y fueron tratadas con gonadotrofinas (400 UI de eCG-200 UI de hCG, Duogestal®, Syntex, SA), presentando celo en promedio a los 4 días del tratamiento. Las ecografías se iniciaron en el día 15 del ciclo estral. Los animales fueron monitoreados una vez al día, para confeccionar mapas ováricos y registrar el número, la localización y el tamaño de los folículos > 3 mm. Se controló la presentación de celo dos veces por día, y desde el momento que mostraron síntomas de estro (0 h), las ecografías fueron realizadas cada 8 horas. El momento de la ovulación se definió como la hora de la primera ecografía sin visualizar imágenes de folículos presuntamente ovulados, menos 4 h. La ovulación se confirmó con una ecografía posterior. Para el control del desarrollo folicular, se utilizó un ecógrafo Tringa S50 (Pie Medical, Holanda) con sonda sectorial y una frecuencia de 5-7.5 MHz. Todos los diagnósticos fueron realizados por la vía transabdominal.
Se registró el número de folículos según clase (3, 4, 5, 6, 7, 8 y 9 mm) y el número total de folículos, desde el primer escaneo hasta la confirmación de la ovulación.
Cerdas pluríparas
El estudio se realizó en una granja comercial y se utilizaron cerdas (n=11) de genética definida y de más de un parto. A partir del destete, las cerdas fueron controladas dos veces por día para corroborar las manifestaciones de celo (según la rutina de trabajo del establecimiento). El inicio del estro (0 h) fue establecido como el momento en que las cerdas fueron observadas con signos de celo. Desde la 0 h, ambos ovarios de cada hembra fueron ecografiados cada 8 h y se registró el número, la localización y el tamaño de los folículos > 3 mm de diámetro, para confeccionar los mapas ováricos. El momento de la ovulación se definió como la hora de la primera ecografía sin visualizar imágenes de folículos presuntamente ovulados, menos 4 h. La ovulación se confirmó con una ecografía posterior.
Las ecografías se realizaron con un ecógrafo Pie Medical S100 (Maastrich, Holanda), con una sonda sectorial y una frecuencia de 5.0-7.5 MHz (Fotos 1 y 2). Se utilizó la vía transrectal, colocando la sonda en una guía de cloruro de polivinilo, con un ángulo fijo de 25°.

Foto 1: Folículos preovulatorios, imagen transrectal.
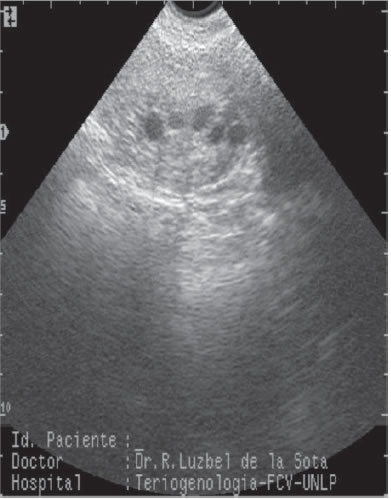
Foto 2: Folículos preovulatorios, imagen transabdominal
El número de folículos de 3 a 5, 6, 7, 8 y 9 mm y el número total de folículos, desde la hora 0 hasta 48 h de observado el estro, fueron analizados con los procedimientos PROC MEANS y PROC REG de SAS®.
Resultados
Cerdas nulíparas
Los folículos de 3 y 4 mm fueron los más numerosos y los únicos visualizados al inicio de los escaneos (día 15 del ciclo estral) y entre las 40 y 20 hs anteriores al inicio del estro, sólo se observaron folículos entre 3 y 5 mm. Con respecto a la dinámica de los folículos de mayor diámetro, los folículos de 6 mm fueron visualizados a las 20 h previas al inicio del estro. Posteriormente, se visualizaron folículos de 7 y 8 mm (al inicio del estro), mientras que los folículos de 9 mm sólo fueron observados en algunas de las hembras y próximo al momento de ovulación (Figura 1).

Figura 1: Cuadrado medio mínimo del número de los folículos (3 a 9 mm) en cerdas nulíparas desde -48 hasta 48 h del inicio del estro.
La ovulación ocurrió a las 36,5±1,8 (promedio±ES) horas desde el inicio del estro y el tamaño de los folículos preovulatorios fue de 5,65±0,06 (promedio±ES) mm.
Cerdas pluríparas
El número de folículos de 3-5 mm fue explicado por una función polinomial cúbica y tuvo un incremento continuo desde las 8 hasta las 32 h posteriores a la observación del estro y luego disminuyó abruptamente (n = 2,15 + [- 0,2897*x] + [0,0244*x2] + [-0,0004*x3]; r2=0,97, P<0,01, Figura 2A). El número de folículos de 6 mm fue explicado por una función cuadrática y disminuyó lentamente desde las 8 hasta las 48 h (Figura 2B) (n = 2,46 + [-0,0069*x] + [- 0,0006*x2]; r2=0,82, P<0,01, Figura N 2B). Un perfil similar siguieron los folículos de 7 mm y su número también fue explicado por una función cuadrática (n=2,36 + [0,0529*x] + [-0,0028*x2]; r2=0,88, P<0,01; Figura 2C). El número de folículos de 8 mm fue explicado por una función polinomial cuadrática y disminuyó desde 8 hasta las 24 h y luego se incrementó hasta las 32 h (n= 2,76 + [-0,1144*x] + [0,0028*x2]; r2=0,90, P<0,01, Figura 2D). El número de folículos de 9 mm fue explicado por una función cuártica y se incrementó desde las 16 hasta las 32 h y luego disminuyó rápidamente en las siguientes 8 h (n= 1,66 + [0,3247*x] + [-0,0661*x2] + [0,0036*x3] + [0,00006*x4]; r2=0,99, P<0,01, Figura 2E).

Figura 2: Cuadrado medio mínimo y error estándar del número de folículos de 3-5 (A), 6 (B), 7 (C), 8 (D) y 9 (E) mm en cerdas pluríparas desde -8 hasta 48 h desde el inicio del estro
El número total de folículos fue explicado por una función cúbica y tuvo una moderada disminución durante las primeras 32 h y luego disminuyó de manera abrupta en las siguientes 8 h (n= 8,93 + [-0,3398*x] + [0,0197*x2] + [- 0,0003*x3]; r2=0,99, P<0,01, Figura 3).
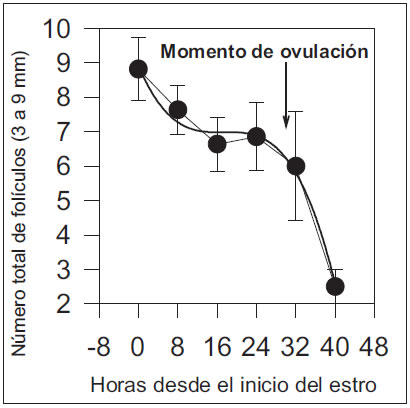
Figura 3: Cuadrado medio mínimo y error estándar del número total de folículos (3 a 9 mm) en cerdas pluríparas desde -8 hasta 48 h del inicio del estro.
La disminución del número total de folículos ováricos durante el período estudiado, se debió principalmente a una reducción en el número de folículos de 6 a 9 mm desde el inicio del estro hasta la ovulación. Los folículos de 6 a 7 mm crecieron a 8 y 9 mm, y así el número de folículos en crecimiento disminuyó y aumentó el número de folículos de mayor diámetro. La disminución de los folículos de 6 a 7 mm ocurrió en el período comprendido entre la 0 y las 24 h desde el inicio del estro y el aumento a folículos de 8 y 9 mm ocurrió desde las 24 hasta las 32 h desde el inicio del celo. Estos dos eventos permitieron formar un "pool" de folículos preovulatorios. Algunos de los folículos de 6 a 7 mm se atresiaron y, entonces, redujeron su tamaño y constituyeron el "pool" de folículos de 3 a 5 mm. El aumento de tamaño del "pool" de folículos de 3 a 5 mm podría deberse a que entran en este grupo los folículos atrésicos, ya que el número de folículos de 6 a 7 mm se mantuvo y no aumentó.
El tamaño folicular disminuyó durante el período estudiado y fue explicado por una función cuadrática (tamaño folicular promedio acumulado, n = 6,78 + [-0,0107*x] + [-0,0010*x2]; r2=0,85, P<0,01, Figura 4). La ovulación ocurrió a las 29,1±2,6 h (promedio±ES) a partir del inicio del estro y el tamaño de los folículos preovulatorios fue de 7,0±1,0 mm, con un intervalo destete-estro cercano a los 6 días (134,2±6,1 h; rango 96-216 h).
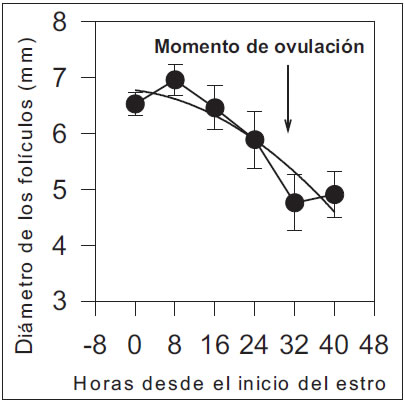
Figura 4: Cuadrado medio mínimo y error estándar del diámetro de los folículos (3 a 9 mm) en cerdas pluríparas desde -8 hasta 48 h del inicio del estro.
Discusión
En este trabajo, cuando se estudió la dinámica folicular en cerdas nulíparas, los folículos visualizados al inicio de las ecografías (hacia finales de la fase luteal) fueron sólo los de 3 a 5 mm. Estos datos coinciden con lo reportado por Clark et al (1982) y por Yen et al (1995), quienes describieron que los folículos de tamaño pequeño y mediano (< 6.5 mm) rápidamente aparecen luego de la ovulación e incrementan el número durante la fase luteal, mientras que, los folículos grandes están ausentes. Sin embargo, luego de la luteólisis, los folículos pequeños y medianos rápidamente desaparecen, mientras que los > 6.5 mm aparecen y aumentan en número cerca del momento de la ovulación. La selección de folículos en cerdas nulíparas maduras y púberes es un proceso único que ocurre en presencia de cuerpos lúteos (CL). Hay varias evidencias que demuestran que, la presencia del CL altera la vascularización e indirectamente puede influir en el crecimiento de los folículos18. Además, la población de folículos difiere dramáticamente antes y después de la formación del CL, y antes y después de la luteólisis.
A pesar de las diferencias genéticas, la FSH parece regular los folículos pequeños-medianos, mientras que la combinación de FSH y LH actúa en el mantenimiento del pool de los medianos. Sin embargo, durante el último período de la selección folicular, los folículos pequeños y medianos cambian su dependencia de la FSH a la LH, ya que, rápidamente pierden receptores de FSH y adquieren los de LH. Luego de este período, los pulsos de LH parecen controlar la maduración folicular final y estabilizar la tasa de ovulación en los folículos saludables > 4 mm. Pareciera que, este proceso es similar en hembras prepúberes y púberes, pero no está claro cómo difiere en el mantenimiento y en el control del pool reclutado antes del momento de la selección17.
El diámetro promedio de los folículos disminuyó con en el tiempo debido a que los folículos ovularon o se atresiaron, y por lo tanto, desaparecieron o disminuyeron en tamaño. Según Soede et al. (1998) el tamaño de los folículos al momento de la ovulación es variable y está altamente relacionado con el tamaño de los folículos presentes 16 h antes de la ovulación. Los folículos < 7 mm presentes 16 h antes de la ovulación aumentan su tamaño hasta la ovulación en la mayoría de las cerdas, mientras que los folículos > 7 mm no lo hacen o bien, lo disminuyen. Esto puede ser interpretado como un aumento compensatorio en las últimas horas antes de la ovulación en cerdas con folículos de menor tamaño. En muchas cerdas, el tamaño folicular parece disminuir aproximadamente 16 h antes de la ovulación. Esto puede deberse a variaciones en las mediciones, o bien, a que los folículos pierden turgencia en los últimos monitoreos previo a la ovulación22.
Sólo los folículos gonadotrofina-dependientes son reclutados. A pesar que, el reclutamiento y la gonadotrofina-dependencia de los folículos están asociados en muchas especies, el número de folículos reclutados es variable según las especies animales, y es de más de 50 en las cerdas7. En todas las especies, los folículos seleccionados parece que son los primeros en desarrollar receptores de LH en las células de la granulosa. Los folículos desarrollan receptores de LH cuando alcanzan 4, 5 a 6, 8 y 25 mm de diámetro en la oveja, cerda, vaca y yegua, respectivamente. En todas las especies, hay una asociación entre este tamaño y el tamaño en que los folículos son seleccionados. En contraste, la intensidad de la selección es variable entre especies. Puede ser muy baja en algunos equinos, mientras que es bastante alta en la vaca (un folículo de cinco) y en la cerda (12 folículos seleccionados a partir de 50).
En estudios del desarrollo folicular en animales cíclicos con restricción alimenticia durante la fase de predominio de progesterona (fase luteal), Hazelger et al (2005) encontraron que la restricción produjo una reducción en el número y en el tamaño de los folículos grandes, hacia el final de la dominancia de la progesterona. Las hembras nulíparas, también presentaron una disminución en la tasa de ovulación. Por lo tanto, el tamaño de los folículos antrales hacia el final de la fase luteal (antes de la selección) puede ser un indicativo del número de folículos que serán capaces de responder a la acción del eje hipotálamo-hipófisis y determinante de la tasa de ovulación21.
El momento promedio de ovulación en las cerdas nulíparas observado en este estudio (36,5±1,8 h, media±ES) fue inferior al promedio citado por Almeida et al. (2001) (44 h, rango de 30 a 60 h).
En el presente trabajo, el tamaño promedio de los folículos preovulatorios (7.0±1.0 mm) observados en hembras pluríparas, fue similar al obtenido por Nissen et al. (1997) (7.6±0.8 mm) y Soede et al. (1998) (7.1±0.9 mm). Sin embargo, Mburu et al. (1995) encontraron que el diámetro de los folículos más grandes durante la ovulación fue de 9,3 mm. La ovulación promedio en el presente trabajo ocurrió a las 29,1±2,6 h (±ES) a partir del inicio del estro, dato inferior a lo hallado por Soede y col (1998) quienes observaron que la ovulación ocurrió a las 35±8 h después de iniciado el estro, o a lo reportado por Dalin y col (1995) con un intervalo de 39±12.4 h desde el inicio del estro hasta la finalización de la ovulación.
El intervalo destete-estro observado en este estudio (134,2±6,1 h) fue inferior al informado por Nissen et al. (1997) (92±12 h).
Conclusiones
Sobre la base de los resultados obtenidos se puede concluir que, el número total de folículos disminuyó a lo largo del período estudiado debido a que sólo fueron seleccionados los folículos que posteriormente alcanzaron un tamaño necesario para la ovulación. El uso de la ultrasonografía para el estudio de la dinámica folicular resulta ser una herramienta de gran utilidad para conocer el crecimiento de los folículos y predecir el momento de ovulación.
Considerando las diferencias observadas entre cerdas nulíparas y pluríparas posdestete en el momento de la ovulación y en el tamaño de los folículos ovulatorios, es de resaltar la importancia que adquiere la aplicación de la ultrasonografía. Además, el tamaño folicular preovulatorio podría ser predecible y en el futuro, con el uso de tratamientos para la inducción y la sincronización del celo, podría estimarse el momento de la ovulación y así, utilizar esta información para el diseño de protocolos para inseminación artificial a tiempo fijo.
1. Almeida, F.R.; Mao, J.; Novak, S.; Cosgrove, J.R.; Foxcroft, G.R. Effects of different patterns of feed restriction and insulin treatment during the luteal phase on reproductive, metabolic and endocrine parameters in cyclic gilts. J Anim Sc. 2001. 79 (1):200-12. [ Links ]
2. Clark, JR; Brazier, SG; Wiginton, LM; Stevenson, GR; Tribble, LM. Time of ovarian follicle selection during the porcine estrous cycle. Theriogenology. 1982. 18:697-708. [ Links ]
3. Corner, GW. The ovarian cycle of swine. Science. 1921. 53:420- 21. [ Links ]
4. Cox, NM.. Control of follicular development and ovulation rate in pigs. J Reprod Fertil Supp. 1997. 52:31-46. [ Links ]
5. Dalin, AM.: Ovarian follicular activity during the luteal phase in gilts. J Vet Med. 1987. 34:592-601. [ Links ]
6. Dalin, A.M.; Nanda, T.; Hultén, F.; Einarsson, S. Ovarian activity at naturally attained oestrus in the sow. An ultrasonographic and LH study. Acta Vet Scand. 1995. 36:377-82. [ Links ]
7. Driancourt, MA. Regulation of ovarian follicular dynamics in farm animals. Implications for manipulation of reproduction. Theriogenology. 2001. 55:1211-39. [ Links ]
8. Evans, ACO. Characteristics of ovarian follicle development in domestic animals. Reprod Dom Anim. 2003. 38:240-46. [ Links ]
9. Foxcroft, G.R.; Hunter, M.G. Basic physiology of follicular maturation in the pig. J Reprod Fertil Suppl. 1985. 33:1-19. [ Links ]
10. Guthrie, H.D.; Cooper, B.S. Follicular atresia, follicular fluid hormones, and circulating hormones during the midluteal phase of the estrous cycle in pigs. Biol Reprod. 1996. 55:543-47. [ Links ]
11. Guthrie, H.D.; Grimes, R.W.; Cooper, B.S.; Hammond, J.M. Follicular atresia in pigs: measurement and physiology. J Anim Sci. 1995. 73:2834-44. [ Links ]
12. Hazeleger, W.; Soede, N.M.; Kemp, B. The effect of feeding strategy during the pre-follicular phase on subsequent follicular development in the pig. Dom Anim Endocr. 2005. 29:362-70. [ Links ]
13. Kauffold, J.; Ritcher, A.; Sobiraj, A. Ultrasonography in swine gynaecology. Applications and perspectives. Proc. of Intern. Conf. On Pig Production, Beijing, 1998. 647-51. [ Links ]
14. Kauffold, J., Rautenberg, T.; Gutjahr, S.; Richter, A.; Sobiraj, A. Ultrasonographic characterization of the ovaries in non-pregnant first served sows and gilts. Theriogenology. 2004. 61:1407-17. [ Links ]
15. Kauffold, J.; Rautenberg, T.; Richter, A.; Wahner, M.; Sobiraj, A. Ultrasonographic characterization of the ovaries and the uterus in prepubertal and pubertal gilts. Theriogenology. 2004. 61:1635-48. [ Links ]
16. Kauffold, J.; Rautenberg, T.; Hoffmann, G.; Beynon, N.; Schellenberg, I.; Sobiraj, A. A field study into the appropriateness of transcutaneous ultrasonography in the diagnoses of uterine disorders in reproductively failed pigs. Theriogenology. 2005. 64:1546-58. [ Links ]
17. Knox, R.V. Recruitment and selection of ovarian follicles for determination of ovulation rate in the pig. Domestic Animal Endocrinology. 2005. 29:385-97. [ Links ]
18. Murphy, B.D.; Gevry, N.; Ruiz-Cortes, T.; Cote, F.; Downey, B.R.; Sirois, J. Formation and early development of the corpus luteum in pigs. Reprod Suppl. 2001. 58:47-63. [ Links ]
19. Mburu, J.N.; Einarsson, S.; Dalin, A.M.; Rodriguez-Martinez, H. Ovulation as determined by transrectal ultrasonography in multiparous sows: relationships with oestrus symptoms and hormonal profiles. J Vet Med. 1995. 42:285-92. [ Links ]
20. Nissen, A.K.; Soede, N.M.; Hytell, P.; Schmidt, M.; Dh'oore, L. The influence of time of insemination relative to time of ovulation on farrowing frequency and litter size in sows, as investigated by ultrasonography. Theriogenology. 1997. 47:1571-82. [ Links ]
21. Soede, N.M.; Bouwman, E.G.; Lanjendijk, P.; van der Laan, I.; Canora, A.; Kemp, B. Follicular development during Luteal Phase and Altrenogest Treatment in Pigs. Reprod Dom Anim. 2007. 42:329-32. [ Links ]
22. Soede, N.M.; Hazeleger, W.; Kemp, B. Follicle size and the process of ovulation in sows as studied with ultrasound. Reprod Dom Anim. 1998. 33:239-44. [ Links ]
23. Soede, N.M.; Kemp, B. In synchronized pigs the duration of ovulation is not affected by insemination and is not a determinant for early embryonic diversity. Theriogenology. 1993. 39:1043-53. [ Links ]
24. Soede, N.M.; Noordhuizen, J.T.P.M.; Kemp, B. The duration of ovulation in pigs, studied by transrectal ultrasonography, is not related to early embryonic diversity. Theriogenology. 1992. 4:653-66. [ Links ]
25. Weitze, K.F.; Habeck O., Willmen, T., Rath D. Detection of ovulation in the sow using transcutaneous sonography. Zuchthygiene. 1989. 24:40-42. [ Links ]
26. Yen, H.W.; Christian, S., Zimmerman, D.R. Elevation of plasma FSH with a low level of FSH-P during the early to mid follicular phase blocks the loss of greater numbers of medium follicles in control line gilts compared to gilts selected for high ovulation rate. Lincoln: University of Nebraska. 1995. 4-8. [ Links ]

