










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkInVet
On-line version ISSN 1668-3498
InVet vol.12 no.2 Ciudad Autónoma de Buenos Aires July/Dec. 2010
REVISIÓN BIBLIOGRÁFICA
Efecto de la nutrición sobre la duración del anestro postparto en vacas de cría
Donzelli, M.V.1; Catalano, R.C.2; Burges, J.C.3; Machado, C.F.2
1Becaria INTA EEA Cuenca del Salado, Belgrano 416 (7203) Rauch, e-mail: mvaleriadonzelli@yahoo.com.ar.
2Facultad de Ciencias Veterinarias de Tandil, UNCPBA, Pinto 399 (7000) Tandil
3Unidad Integrada Balcarce INTA-UNMdP, ruta 226, km 73,5 (7620) Balcarce
Recibido: 01/11/2010
Aceptado: 10/02/2011
Resumen
Unos de los aspectos fundamentales para el éxito económico y productivo en los rodeos de cría es la eficiencia reproductiva. El estado nutricional es uno de los principales factores que regulan la respuesta reproductiva y se ha señalado que el mismo puede ser evaluado a través de la condición corporal. Un inadecuado consumo de nutrientes pre y postparto puede afectar la duración del anestro postparto. Es posible que las hormonas IGF-I, tiroideas y prolactina interactúen en el restablecimiento del normal funcionamiento del eje hipotálamo - hipófisis - ovario luego del parto y de esta manera se alargue el período de anestro postparto. La evaluación del estado nutricional a través del monitoreo de la condición corporal es una herramienta de manejo importante para la toma de decisiones las cuales son necesarias tanto para el óptimo resultado reproductivo como en rentabilidad. La incorporación de modelos matemáticos facilita la comprensión de los procesos involucrados para la investigación y la docencia.
Palabras clave: Bovino; Parto; Condición corporal; Estro; Ovulación.
Effect of nutrition on the length of postpartum anestrus in beef cows
Summary
The reproductive efficiency is one of the key issues for achieving productive and economical success in cow-calf beef systems. The nutritional condition of the cow, represented by the body condition score, is one of the main drivers of the productive response. A nutritional deficit pre or postpartum may extend the postpartum anestrus. Thyroid, IGF-I and prolactin hormones may contribute to the onset of postpartum normal link between hypothalamus- hipophisis - ovary. Body condition score is a relevant monitoring variable as a management tool for adequate decision making to achieve optimum outcomes both in reproduction and in profitability. The use of mathematical models helps to understand the involved processes for research and teaching.
Key words: Bovine; Calving; Body condition; Oestrus; Ovulation.
Introducción
La rentabilidad de un sistema de cría está relacionada con la respuesta reproductiva del rodeo50, 28 dependiendo dicha relación del precio relativo de los vientres de descarte y de los terneros vendidos42.
El amamantamiento, el consumo de nutrientes y el estado nutricional son los principales factores que regulan la respuesta reproductiva50. En lo que respecta a este último factor, se ha señalado que el mismo puede ser evaluado a través de la condición corporal (CC), ya que refleja las reservas corporales disponibles para el metabolismo, crecimiento y lactación53. Además, existe una estrecha relación entre CC al parto y el reinicio de la ciclicidad postparto que ha sido evaluada en numerosos estudios40, 20, 28, 2. Cuando existen inadecuadas reservas corporales al parto, se produce un incremento del intervalo parto primer celo y un bajo consumo de nutrientes postparto y la lactancia pueden incrementar la duración de dicho intervalo en vacas con baja CC. Ante esta situación, la vaca permanece en anestro postparto, que es el período en el cual no muestra signos de actividad sexual y la ovulación no ocurre debido a que los folículos en crecimiento no llegan a madurar39. Es necesario una compleja interacción entre hormonas, compuestos metabólicos y otros factores para lograr la maduración folicular, el estro y la ovulación postparto en vacas de cría50.
Dado que en nuestros sistemas de producción, la nutrición sigue afectando significativamente la duración del anestro postparto y que existen nuevas investigaciones sobre el tema, se consideró de interés realizar una revisión bibliográfica con el objetivo de estudiar los efectos de la CC sobre la respuesta reproductiva postparto en vacas de cría. Esta revisión se concentrará en resultados productivos y reproductivos generales, mencionando de forma somera algunos procesos fisiológicos que relacionan la nutrición y el reinicio de ciclicidad. Para consideraciones más particulares de estos puntos se recomiendan otros trabajos54, 51, 37.
Nutrición pre y postparto y su relación con la condición corporal y la respuesta reproductiva
La subnutrición o un inadecuado consumo de nutrientes causa pérdida de peso y estado corporal y esto se asocia con ausencia de manifestación de celos en vacas de cría durante el postparto44, 6, 26. Vacas con una CC de 4 o menos (escala de 1 a 9) durante el postparto no conciben a un nivel aceptable y tienen más de 12 meses de intervalo entre partos comparando con vacas con una CC de 5 o más. Además, la CC al parto está asociada con la lactación, salud y el vigor del ternero recién nacido21. Cuando las vacas tienen una CC moderada al parto (5 a 7 en escala de 1 a 9), el número de días desde el parto al primer estro es de 15 a 35% menor que en vacas que paren con una condición corporal menor a 550. En vacas de cría, la nutrición preparto es una mejor determinante del largo del anestro comparada con la nutrición postparto16, 17, 43, 48. Según Hess et al., (2005)22, la longitud del intervalo entre partos estuvo correlacionada con la CC al parto (r = 0,75; p < 0,001) y con el balance energético preparto estimados a partir de los cambios en la CC preparto (r= 0,52; p= 0,01). La CC al parto fue la única variable a considerar para estimar los efectos del plano nutricional preparto sobre el intervalo entre partos (p < 0,001; R2 =0,57), lo cual coincide con lo observado por otros autores23. Asimismo, Lake et al., (2004)29 realizaron un ensayo con vacas manejadas nutricionalmente para lograr una CC de 4 y 6 al parto y luego fueron alimentadas a niveles de mantenimiento durante la lactación. Las tasas de concepción al primer servicio no fueron afectadas (p= 0,22), pero la tasa de preñez fue menor (p= 0,01) para las vacas con CC de 4 (63,9%) comparadas con las vacas de CC 6 (88,9%).
Asimismo es importante destacar el efecto de la nutrición postparto y la influencia de la CC en el servicio. Herd y Sprott, (1996)21 señalan que la CC al servicio afecta la respuesta reproductiva en términos de servicios por concepción, intervalo entre partos y porcentaje de vacas vacías y que durante la época de servicio también es necesario una nutrición adecuada para lograr éxito reproductivo.
Por otra parte, también se han realizado trabajos que permiten estudiar la interacción del efecto de la nutrición preparto y postparto. Lalman et al. (1997)30 determinaron la influencia del cambio de peso vivo y CC sobre la duración del anestro postparto en vaquillonas en condiciones de subnutrición. Para ello fueron manejadas nutricionalmente para llegar con una CC de 4 (escala 1 a 9) al parto. Luego del mismo, se les asignó al azar 4 dietas de diferente concentración energética para crear un rango de cambio de CC. Los resultados indicaron que un incremento en la concentración energética de la dieta aumentó el peso y la CC de las vacas hacia los 90 días postparto y el intervalo parto primer estro disminuyó (p < 0,01) a medida que la concentración de energía metabolizable aumentó. Además, en vaquillonas en balance energético negativo preparto, la CC al parto fue el mejor indicador del período de anestro comparado con el cambio de peso vivo y CC. Las vacas alimentadas en un alto plano nutricional 55 días antes y 40 días después del parto presentaron en promedio 35 kg más y un intervalo parto primer celo más corto que vacas alimentadas con un plano nutricional medio38. Una subnutrición severa en el último trimestre de la gestación y en el postparto puede resultar en ausencia de folículos ováricos mayores que 8 mm de diámetro que son los encargados de producir importantes cantidades de estradiol41, 25. Ciccioli et al. (2003)12 estudiaron la influencia de la CC al parto y la nutrición postparto sobre las funciones endocrinas y ováricas y la respuesta reproductiva. Para ello, seleccionaron al azar vacas de baja CC (CC 4) y con CC moderada (CC 5), para recibir uno de dos tratamientos nutricionales después del parto. Ambos grupos fueron alimentados para ganar 0,45 kg/d (M, n=17) o 0,90 kg/d (H, n=17) respectivamente en los primeros 71 días postparto. Posteriormente, todas las vacas fueron alimentadas con la dieta M después de los primeros 21 días posteriores al primer estro. Las vacas que parieron con CC de 4 o 5 tuvieron similar función endocrina y reproductiva al primer estro, pero durante el tratamiento, las vacas alimentadas con la dieta H, ganaron peso e incrementaron la CC y tuvieron un intervalo al primer estro más corto y una mayor tasa de preñez al primer estro. Lents et al. (2008)32 realizaron un ensayo similar al citado anteriormente para lo cual administraron dos dietas de diferente concentración proteica y energética a vacas con pobre y moderada CC. De esta manera estudiaron el efecto de la CC al parto y la suplementación proteica al postparto sobre el estro y las características de los folículos dominantes en vacas de cría. Se observó que la duración del período parto primer estro fue menor y la tasa de preñez fue mayor en las vacas con mejor CC y alimentadas con la dieta de mejor calidad.
Relación entre amamantamiento y condición corporal
Por otra parte, se ha considerado al amamantamiento como un factor estrechamente relacionado con la CC y la respuesta reproductiva postparto. Loudon (1987)33 propuso que los efectos inhibitorios del amamantamiento y la subnutrición interactúan y Short et al. (1990)49 establecieron que el amamantamiento prolonga el anestro postparto y que el efecto es de mayor magnitud en vacas primíparas y con pobre CC. Además, las reservas corporales al parto influencian el efecto del amamantamiento sobre la función ovárica50. Si las vacas presentan una CC mayor o igual que 5 al parto y los terneros son destetados a los 35 días postparto, todas las vacas ovulan alrededor de los 25 días post destete7. Contrariamente, sólo el 40 % de las vacas con CC menor a 5 ovulan 25 días después del destete. A pesar que el intervalo del destete a la primera ovulación es mayor en vacas delgadas comparado con vacas en mejor estado corporal, el destete es una herramienta de manejo útil para incrementar la tasas de preñez en vacas con pobre estado corporal50. Escobedo-Amezcua et al. (2010)18 realizaron un estudio con el objetivo de determinar los efectos del método de amamantamiento sobre el peso vivo, CC y el reinicio de la actividad ovárica postparto en vacas Bos taurus X Bos indicus. Para ello, las mismas fueron asignadas al azar a uno de dos tratamientos: amamantamiento continuo (C) con la permanencia del ternero las 24 horas o restringido (R) en el cual el ternero sólo permanecía con la madre por un tiempo de 30 minutos. Los resultados indicaron que la restricción del amamantamiento mejoró la condición corporal de las vacas, la tasa de ovulación y la tasa de concepción fueron mayores (p<0,001), el intervalo parto primer celo fue más corto (p<0,005) y como resultado final, la tasa de preñez fue mayor para el tratamiento R (p<0,001). Los autores concluyeron que la restricción del amamantamiento afectó la CC y las tasas de ovulación y concepción en vacas Bos taurus X Bos indicus.
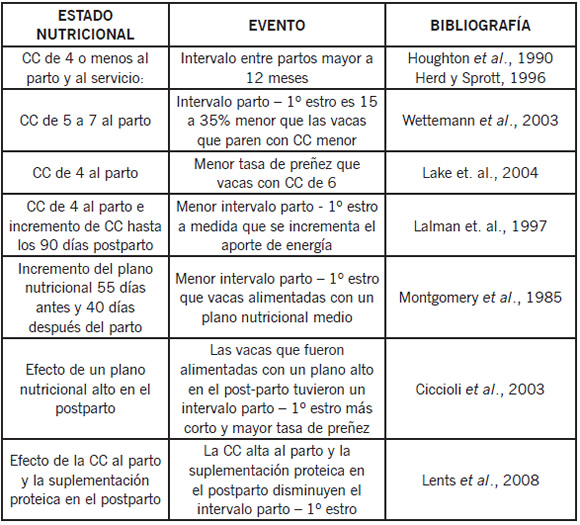
En la Tabla 1 se presenta un resumen de los resultados que los distintos autores mencionados han observado al evaluar el efecto del estado nutricional pre y postparto sobre la respuesta reproductiva.
Tabla 1. Efecto del estado nutricional y su efecto sobre la respuesta reproductiva en vacas de cría según distintos autores
Procesos fisiológicos que relacionan la nutrición con la reproducción postparto
La reanudación del crecimiento folicular después del parto resulta del incremento de la secreción de FSH entre los 3 a 4 días posteriores al parto en vacas con adecuado estado nutricional31. Pero la habilidad de crecer y transformarse en folículos dominantes depende de la secreción pulsátil de LH lo cual es el resultado de la liberación de GnRH desde el hipotálamo26, 13.
Las reservas corporales regulan la secreción hipotalámica e hipofisiaria que controlan la función ovárica45. Un mecanismo importante por el cual el déficit energético condiciona la actividad reproductiva es por suprimir la descarga de GnRH y como consecuencia, la frecuencia de pulsos de LH necesarios para lograr el crecimiento folicular44, 47. Una nutrición deficiente en el postparto modifica los cambios neuroendocrinos normales, particularmente la frecuencia de descarga de LH46 lo que determina prolongados intervalos de anestro postparto. Jolly et al. (1991)24 indicaron que vacas restringidas en su consumo energético durante el postparto tuvieron concentraciones de FSH y LH deprimidas, lo que se asoció con un mayor tiempo de anestro comparado con vacas que tuvieron dietas adecuadas. Lucy (2008) (35) propone otro mecanismo de acción relacionado con una menor sensibilidad ovárica a la acción de las gonadotrofinas y señala a la insulina e IGF-I como posibles mediadores de esta acción. Por otro lado, Flores et al. (2008)19 estudiaron el efecto de la somatrotrofina bovina (STb) sobre la dinámica folicular y su relación con la concentración de IGF-I, las hormonas tiroideas, la prolactina y la CC. Los autores determinaron que vacas con mejor CC (6,1 ± 0,1; escala 1 a 9) tuvieron mayor concentración de IGF-I independientemente si estaban o no cíclicas. A su vez señalaron que las cíclicas presentaron mayor concentración de IGF-I comparadas con vacas en anestro para una misma CC. También observaron que el diámetro de los folículos dominantes fue mayor en vacas en anestro tratadas con STb comparadas con vacas en anestro no tratadas, que la T3 y la prolactina fueron mayores en vacas con moderada CC y que el diámetro de los folículos dominantes estuvo correlacionado positivamente con la IGF-I y la prolactina. Este estudio permite suponer que el estado de subnutrición está relacionado con el eje hipotálamo-hipófisis-gónada por mecanismos hormonales metabólicos que quizás incluyen a IGF-I, hormonas tiroideas y prolactina.
Decisiones de manejo
La CC puede ayudar a identificar problemas en el rodeo. Al evaluar la tasa de preñez, el productor puede determinar si la misma estuvo condicionada por el estado nutricional o por otros factores28. Si las vacas tienen una CC de 5 o más tendrán 90% de preñez y por lo tanto factores nutricionales no estarían limitando la tasa de preñez28. Aquellos animales que no tengan adecuada CC deberían ser separados del rodeo y ser asignados a las pasturas de mejor calidad para acelerar la ganancia de peso e incremento de la CC28.
Una nutrición adecuada puede permitir al productor lograr mayor eficiencia y sustentabilidad en el tiempo. Para ello, es importante la identificación de factores nutricionales que son potencialmente detrimentales de la capacidad reproductiva del rodeo y desarrollar programas nutricionales basados en una dieta óptima para mantenimiento de peso y aumentar así la eficiencia reproductiva22. Por ejemplo, dietas ricas en lípidos estimulan el crecimiento folicular en vacas de cría34 y durante el último tercio de la gestación, es un método para aliviar el efecto negativo de un inadecuado manejo nutricional sobre la respuesta reproductiva22.
Alexander et al. (2002)1 demostraron un mejoramiento en las tasas de preñez cuando vacas de cría fueron suplementadas con lípidos (91,6%) durante la última fase de la gestación comparado con las vacas control. Bellows (1999)5 encontró tasas de preñez de un 70 % en vacas de cría suplementadas con lípidos y 50 % de preñez en vacas control. Con estos resultados, concluyeron que la suplementación con lípidos durante la última fase de la gestación es efectiva para lograr éxitos reproductivos en el período de servicio.
Por otra parte, en rodeos de parición invernal y luego de haber realizado el destete de los vientres, es posible someter a restricción alimentaria otoño-invernal sin perjudicar el porcentaje de preñez11. Es decir, restringir cuando el animal por efecto del destete y por una preñez muy pequeña tiene bajos requerimientos con la finalidad de reservar forraje para suministrarlo durante el postparto a fines de invierno e inicio de primavera. La condición necesaria para someter a la restricción a una vaca preñada es que al momento de iniciarla estén en buen peso y estado corporal y que luego de la parición se mejore su nivel de alimentación. La restricción alimentaria invernal de la vaca de cría es un fenómeno que se produce comúnmente y en forma casi natural en todos los rodeos de cría como resultado de que el invierno es el período del año con menor crecimiento de las pasturas y pastizales. Este proceso natural compromete la performance reproductiva de los rodeos. El manejo de este fenómeno con los recaudos correspondientes puede presentar resultados distintos a los que comúnmente se obtienen. Según Carrillo et al. (1998)11, durante treinta años de estudio en la Reserva 6 de la EEA Balcarce, el manejo de la alimentación con restricción alimentaria durante el otoño-invierno y sobrealimentación durante el postparto no retrasó la nueva concepción y permitió una adecuada tasa de preñez durante más de treinta años de experiencia. El intervalo parto-celo y la tasa de preñez de vacas en menor condición corporal al final del período de parición pueden ser mejorados si la alimentación durante la lactancia permite una ganancia de peso (Figura 1)10.
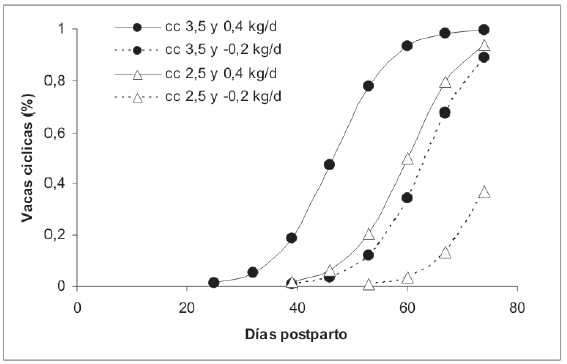
Figura 1. Porcentaje de vacas cíclicas según los días postparto en vacas de cría con diferente condición corporal y cambio de peso vivo (Burges y Romera, 2003).
Datos más recientes obtenidos en trabajos realizados por Donzelli et al. (2009)15 muestran que es posible obtener altos porcentajes de preñez en vacas Aberdeen Angus restringidas en el invierno. El objetivo del trabajo fue evaluar el comportamiento reproductivo postparto (agosto a diciembre de 2007) en módulos ganaderos que fueron diseñados con diferente carga animal objetivo (CO). En ellos se evaluó un sistema de manejo basado en pastoreo continuo y en reglas de decisión vinculadas a la altura de las pasturas y a la CC de los animales9. Las vacas pastorearon durante todo el año, excepto a fin de otoño e invierno cuando se alimentaron a corral hasta el parto con el heno producido el año anterior en cada CO. Este protocolo se aplicó en 6 potreros de 10 ha cada uno con diferente CO (0,9; 1,3; 1,7; 2,1; 2,5; 2,9 cabezas/ha) integrada por vacas multíparas, primíparas y vaquillonas de reposición. Cuando las condiciones nutricionales y/o de pastoreo se tornaron críticas, actuaron las reglas de descarte de animales por lo cual el peso vivo (PV) y la CC (CC: escala 1-5) no fueron un fiel reflejo de la CO sino de las condiciones propias del sistema. Además, la carga real (CR) fue menor a CO en algunos casos. Como se puede observar en la Tabla 2, el grupo con menor porcentaje de preñez es aquél que presentó mayor CR, menor estado corporal al inicio y al final del servicio y menor altura de las pasturas en el período postparto.
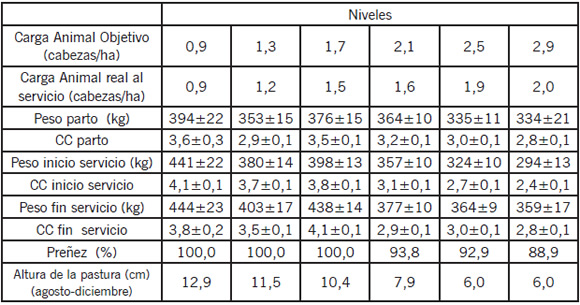
Tabla 2. Peso vivo y condición corporal al parto, inicio y fin de servicio (media ± E.E), altura de las pasturas del período postparto y porcentaje de preñez en módulos productivos manejados con diferente carga animal objetivo (Donzelli et al, datos no publicados).
De la bibliografía consultada surge claramente que existen diferencias entre estudios en lo que respecta a clima, región, raza, estado corporal, etc. lo que hace difícil establecer recomendaciones rígidas de manejo de nutrición pre y postparto. En tal sentido, se ha mencionado que el grado de conocimiento de una temática está dado por la capacidad predictiva en términos cuantitativos14. Dentro de ese enfoque se han utilizado modelos matemáticos para representar la respuesta reproductiva con distinto nivel de detalles tanto en ovinos8 como en bovinos27, 3. Los modelos constituyen una herramienta de síntesis de información4. En la Figura 2 se muestra de forma esquemática los distintos factores representados en el modelo de Khan y Lehrer (1984)27, muchos de los cuales han sido revisados en este documento. De manera de utilizar estos modelos para el apoyo de la docencia, la extensión y la investigación, Machado et al. (2009)36 desarrollaron un software que permite cuantificar los efectos de la variación de estos factores. En la Figura 3 se muestra una pantalla de carga de datos del software mencionado donde se ingresaron 9 lotes de vacas en un arreglo factorial 3 x 3 que se diferencian en el tiempo postparto (15, 45 y 75 días) y en la condición corporal (4, 5 y 6, en la escala 1 a 9). De esto se desprende que este tipo de modelos simplificados permite experimentar con combinaciones que resultarían difícil desde el punto de vista operativo y de recursos bajo investigación de campo52. En la Figura 4 se observa la pantalla de resultados del software anteriormente mencionado donde se muestra una de las variables respuesta (porcentaje de preñez) y la comparación entre dos grupos de animales. Es importante señalar que el modelo es una versión simplificada de la realidad y para un uso eficiente del mismo resulta clave disponer de información adecuada para la calibración y evaluación. Lamentablemente, la información no siempre está fácilmente disponible. En tal sentido, los autores de esta revisión participan en el proceso de calibración de modelos de simulación, como el mencionado, incluyendo también datos propios15.
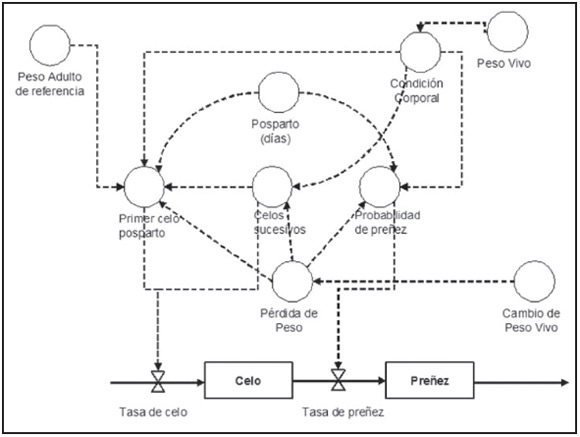
Figura 2. Factores y estructura de un modelo reproductivo (Kahn y Lehrer, 1984). Las líneas punteadas indican flujo de información y las continuas, flujo de animales en diferentes estados.

Figura 3. Pantalla de ingreso de datos de 9 lotes de vacas con diferentes tiempos postparto y estado corporal. En la mitad derecha de la pantalla se observa un detalle del lote de vacas con 75 días de postparto al momento del inicio del servicio y CC 6 (escala 1 a 9). Fuente: Machado et al., 2009
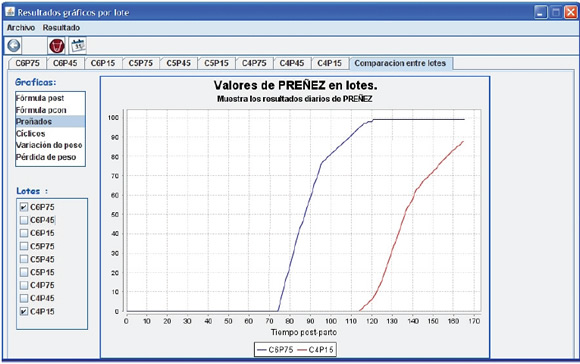
Figura 4. Pantalla de resultados del modelo donde se compara el porcentaje acumulado de preñez de vacas con CC 6 y 75 días postparto (C6P75) y con CC 4 y 15 días postparto (C4P15). Fuente: Machado et al., 2009.
Conclusión
La reproducción es uno de los componentes más importantes de la eficiencia productiva del rodeo y una de las principales causas que afectan la performance reproductiva es el intervalo entre partos, el cual está directamente influenciado por el anestro postparto. El consumo de nutrientes pre y postparto condiciona la duración del anestro postparto ya que una buena respuesta reproductiva requiere vacas con una CC entre 5 y 7 (escala de 1 a 9) en el período de servicios. Si las reservas corporales son inadecuadas al parto y el consumo de nutrientes postparto es reducido, se verá incrementado el intervalo del parto a la primera ovulación, afectando de esta manera la eficiencia reproductiva y productiva del rodeo. Separar los vientres según la CC 3 meses antes del parto, puede permitir al productor alimentar de manera diferencial y así permitir la recuperación de las reservas de aquellas vacas que lo requieran. Usar la CC como una herramienta de manejo, puede proveer información útil del estado nutricional del rodeo y de esta manera se pueden tomar las decisiones que sean necesarias para lograr un adecuado comportamiento reproductivo y productivo con el objeto de mejorar la rentabilidad de la empresa ganadera. La síntesis del conocimiento en herramientas cuantitativas como los modelos matemáticos, ofrece la posibilidad de complementar la comprensión de los procesos tanto para la investigación, la docencia y la extensión de la respuesta reproductiva en condiciones de campo.
1. Alexander, B.M.; Hess, B.W.; Hixon, D.L.; Garret, B.L.; Rule, D.C.; Mc Farland, M.; Bottger, J.D.; Simms, D.D.; Moss, G.E. 2002. Influence of fat supplementation on beef cow reproduction and calf performance. Prof. Anim. Sci. 18:351-357. [ Links ]
2. Arreguin, J.A.A.; Santos, R.E.; Villa-Godoy, A.; Roman Ponce, H. 1997. Dinámica folicular en vacas cebú con diferente condición corporal y frecuencia de amamantamiento durante el periodo anovulatorio postparto. División de Educación Continua, UNAM, F.M.V.Z (Eds.), VII Curso Internacional de Reproducción bovina. Méx., D.F., pp 210-240. [ Links ]
3. Azzan, S.; Kinder, J. 1990. Modelling reproductive management systems for beef cattle: Agricultural Systems, 34: 103-115. [ Links ]
4. Barioni, L.G.; Woodward, S.J.R.; Martha Jr, G.B.; Machado, C.F.; Oltjen, J.W.; Sainz, R.D. 2004. Current and potential contributions of mathematical modeling to the management of grazing systems. II Symposium on Grassland and ecophysiology and grazing ecology. 11 -14 de october 2004. Curitiba - Paraná - Brasil. 3-25. [ Links ]
5. Bellows, R. A. 1999. Some effects of feeding supplemental fat to beef cattle. Pages 81-87 in Proc. Range Beef Cow Symp. XVI. Greeley, CO. [ Links ]
6. Bishop, D.K.; Wettemann, R.P. 1993.Pulsalite infusion of gonadotropin-releasing hormone initiates luteal activity in nutritional anestrous beef cows. J. Anim. Sci. 71, 2714-2720. [ Links ]
7. Bishop, D. K..; Wettemann, R.P.; Spicer, L.J. 1994. Body energy reserves influence the onset of luteal activity after early weaning of beef cows. J. Anim. Sci. 72:2703-2708. [ Links ]
8. Blackburn, H.; Cartwright, T. 1987. Description and validation of the Texas A&M Sheep Simulation. J. Anim. Sci., 65: 373-386. [ Links ]
9. Burges, J.C.; Romera A.; Continanza, G.; Sciotti, A.E.; Faverín, C.; Fernández, H.H. 2008. Un sistema de manejo pastoril ligado al estado de las pasturas "SIMPLE". 1. Manejo del pastoreo. Rev. Arg. Prod. Anim. 28 (supl. 1): 255-256. [ Links ]
10. Burges, J.C. y Romera, A. 2003. Condición corporal, nivel nutricional y fertilidad de vacas paridas en otoño. Rev. Arg. de Prod. Anim. 23: 304-306. [ Links ]
11. Carrillo, J.; Sciotti, A.E.; Odriozola, E.; Marino, M.A.; Schiersmann, G.C.S. 1998. Reserva 6: Un sistema de producción de cría vacuna a través de 30 años. Aspectos físicos y biológicos. Rev. Arg. Prod. Anim. Vol 18 nº 3-4: 211-225. [ Links ]
12. Ciccioli, N.H.; Wettemann, R.P.; Spitzer, L.J.; Lents, C.A.; White, F.J.; Keisles, D.H. 2003. Influence of body condition al calving and postpartum nutrition on endocrine function and reproductive performance of primiparous beef cows. J.Anim.Sc 81: 3107-3120. [ Links ]
13. Crowe, M.A. 2008. Resumption of ovarian cyclicity in post partum beef and dairy cows. Reprod. Domest. Anim. 43 Suppl 5, 20-8. [ Links ]
14. Demment, M. W.; Peyraud, J. L.; Laca, E. A. Herbage intake at grazing: a modelling approach. In: Journet, M.; Grenet, E.; Farce, M.H.; Theriez, M.; Demarquilly, C. Eds. Recent development in Nutrition of Ruminants. 1995:121-141. [ Links ]
15. Donzelli, M.V.; Burges, J.C.; Machado, C.; Faverín, C.; Catalano, R. 2009. Comportamiento reproductivo postparto de vacas de cría con diferente carga animal en pastoreo continuo. Rev.Arg. Prod. Anim. Vol 29, Supl,1, p 322-323. [ Links ]
16. Dunn, T.G.; Kaltenbach, C.C. 1980. Nutrition and postpartum interval of ewe, sow, and cow. J. Anim. Sci. 51 (Supl 2), 29-39. [ Links ]
17. Dziuk, P.J.; Bellows, R.A., 1983. Management of reproduction in beef cattle, sheep and pigs. J. Anim. Sci.57 (Suppl. 2), 355-362. [ Links ]
18. Escobedo-Amezcua, F.; Nuncio-Ochoa, M.G.J.; Herrera-Camacho, J.; Gomez-Ramos, B.; Segura-Correa, J.C.; Gallegos-Sanchez, J. 2010. Effect of restricted suckling on body weight, body condition score and onset of postpartum ovarian activity in F1 cows under tropical conditions. J. Anim. Vet. Adv. 9: 79-84. [ Links ]
19. Flores, R.; Looper, M.L.; Rorie, R.W.; Hallford, D.M.; Rosenkrans Jr, C.F. 2008. Endocrine factors and ovarian follicles are influenced by body condition and somatotropin in postpartum beef cows. J.Anim.Sci. 86, 1335-1344. [ Links ]
20. Galina, C.S.; Arthur, G.H.; 1989. Review of cattle reproduction in the tropics. 3. Puerperium. Anim. Breed. Abstr.57, 889-910. [ Links ]
21. Herd, D.B.; Sprott, L.R. 1996. Body Condition, nutrition and reproduction of beef cows. Texas Agricultural Extension Service. B-1526. [ Links ]
22. Hess, B.W.; Lake, S.L.; Scholljegerdes, E.J.; Weston, T.R.; Nayigihugu, V.; Molle, J.D.C; Moss, G.E. 2005. Nutritional controls of beef cow reproduction. J.Anim.Sci. 83: E90-106. [ Links ]
23. Houghton, P.L.; Lemenager, R.P.; Horstam, L.A.; Hendrix, K.S.; Moss, G.E. 1990. Effect of body composition, pre and post partum energy level and early weaning on reproductive performance of beef cows and preweaning calf gain. J.Anim.Sci. 68: 1438-1446. [ Links ]
24. Jolly, P.D.; McSweeney, C.S.; D'Occhio, M.J.; Entwistle, K.W. 1991. Nutritional, suckling and ovarian feedback effects on plasma LH and FSH during post-partum anoestrus in Brahman-cross cows. Proc. Austr. Soc. Reprod. Biol. 23, 152. [ Links ]
25. Jolly, P.D. 1992. Physiological and Nutritional Aspects of Postpartum Acyclicity in Bos indicus Cows. Ph.D.Thesis, James Cook University of North Queensland, Townsville. [ Links ]
26. Jolly, P.D.; McDougall, S.; Fitzpatrick, L.A.; Macmillan, K.L.; Entwistle, K.W. 1995. Physiological effects of undernutrition on postpartum anoestrus in cows. J. Reprod. Fert. 49 (Suppl.), 477-492. [ Links ]
27. Kahn, H.E.; Lehrer, A.R. 1984. A dynamic model for the simulation of cattle herd production systems: Part 3 - Reproductive performance of beef cows. Risk analysis applied to rangeland livestock projections. Agricultural Systems 13: 143-159. [ Links ]
28. Kunkle, W.E.; Sand, R.S.; Owen Rae, D. 1998. Effects of body condition on productivity in beef cattle.University of Florida. SP-144. [ Links ]
29. Lake, S.L.; Hess, B.W.; Rule, D.C.; Scholljegerdes, E.J.; Nayigihugu, V.; Atkinson, R.L.; Murrieta, C.M. 2004. Effects of supplemental hight-linoleate or hight-oleate safflower seeds on adipose tissue fatty acids, apparent mobilization, and potential uptake and storage in pos partum cows. Proc. West. Sect. Am. Soc. Anim. Sci. 55:29-35. [ Links ]
30. Lalman, D.L.; Keisler, D.H.; Williams, J.E.; Scholljegerdes, E.J; Mallett, D.M. 1997. Influence of postpartum weight and body condition change on duration of anestrus by undernourished suckled beef heifers. J. Anim. Sci. 75:2003-2008. [ Links ]
31. Lamming, G.E.; Wathes, D.C.; Peters, A.R. 1981. Endocrine patterns in the post-partum cow. J. Reprod. Fert. 30 (Suppl.), 155-170. [ Links ]
32. Lents, C.A.; White, F.J.; Ciccioli, N.H.; Wettemann, R.P.; Spitzer, L.J.; Lalman, D.L. 2008. Effects of body condition score al parturition and postpartum protein supplementation on estrous behavior and size of the dominant follicle in beef cows. J.Anim. Sci. 86: 2549-2556. [ Links ]
33. Loudon, A. 1987. Nutritional effects on puberty and lactational infertility in mammals: some interspecies considerations. Proc. Nutr. Soc. 46, 203-216. [ Links ]
34. Lucy, M. C.; Savio, J.D.; Badinga, L.; De La Sota, R. L.; Thatcher, W. W. 1992. Factors that affect ovarian follicular dynamics in cattle. J. Anim. Sci. 70:3615-3626. [ Links ]
35. Lucy, M.C. 2008. Functional differences in the growth hormone and insulin-like growth factor axis in cattle and pigs: implications for post-partum nutrition and reproduction. Reprod. Domest. Anim. 43 Suppl 2,31-9. [ Links ]
36. Machado, C.F.; Catalano, R.; Ferragut, S.; Arroqui, M.; Mangudo, P. 2009. Avances en el diseño y desarrollo de una calculadora reproductiva de cría bovina ("Repro-calc") para el apoyo de la docencia, la extensión y la investigación. I Congreso Argentino de Agroinformática. SADIO, Mar del Plata: 12-24. [ Links ]
37. Miller, V.; Ungerfeld, R. 2008. Weekly bull exchange shortens postpartum anestrus in suckled beef cows. Theriogenology 69: 913-917. [ Links ]
38. Montgomery, G.W.; Scott, I.C.; Hudson, N. 1985. An interaction between season of calving and nutrition on the resumption of ovarian cycles in post-partum beef cattle. J. Reprod. Fert. 73, 45-50. [ Links ]
39. Montiel, F.; Ahuja, C. 2005. Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: a review. Anim. Reprod. Sci. 85: 1-26. [ Links ]
40. Moore, C.P.; Campos da Rocha, C. 1983. Reproductive performance of Gyr cows: the effect of weaning age of calves and postpartum energy intake. J. Anim. Sci. 57, 807-814. [ Links ]
41. Perry, R.C.; Corah, L.R.; Cochran, R.C.; Beal, W.E.; Stevenson, J.S.; Minton, J.E.; Simms, D.D.; Brethour, J.R. 1991. Influence of dietary energy on follicular development, serum gonadotrophins, and first postpartum ovulation in suckled beef cows. J. Anim. Sci. 69, 3762-3773. [ Links ]
42. Ponssa, E.; Rodríguez, G.; Sánchez Abrego, D. 2007. Relaciones entre la tasa de preñez y la dinámica del rodeo en un sistema de cría bovina. Evaluación física y económica. Anales de la XXXVIII Reunión Anual de la Asociación Argentina de Economía Agraria. Mendoza, octubre 2007. [ Links ]
43. Richards, M.W.; Spitzer, J.C.; Warner, M.B. 1986. Effect of varying levels of postpartum nutrition and body condition at calving on subsequent reproductive performance in beef cattle. J. Anim. Sci. 62, 300-306. [ Links ]
44. Richards, M.W.; Wettemann, R.P.; Schoenemann, H.M. 1989. Nutritional anestrus in beef cows: body weight change, body condition, luteinizing hormone in serum and ovarian activity. J. Anim. Sci. 67, 1520-1526. [ Links ]
45. Robertson, M.S.; Stumpf, T.T.; Wolfe, M.W.; Cupp, A.S.; Kojima, N.; Werth, L.A.; Kittok, R.J.; Kinder, J.E. 1992. Circulating gonadotrophins during restricted energy intake in relation to body condition in heifers. J. Reprod. Fert. 96, 461-469. [ Links ]
46. Robinson, J.J. 1990. Nutrition in the reproduction of farm animals. Nutr. Res. Rev. 3, 267-268. [ Links ]
47. Schillo, K.K. 1992. Effects of dietary energy on control of luteinizing hormone secretion in cattle and sheep. J. Anim. Sci. 70, 1271-1282. [ Links ]
48. Selk, G.E.; Wettemann, R.P.; Lusby, K.S.; Oltjen, J.W.; Mobley, S.L.; Rasby, R.J.; Garmendia, J.C. 1988. Relationship among weight change, body condition and reproductive performance of range beef cows. J. Anim. Sci. 66, 3153-3159. [ Links ]
49. Short, R. E.; Bellows, R. A.; Staigmiller, R. B.; Berardinelli, J. G.; Custer, E. E. 1990. Physiological mechanisms controlling anestrus and infertility in postpartum beef cattle. J. Anim. Sci. 68:799-816. [ Links ]
50. Wettemann, R.P.; Lents, C.A.; Ciccioli, N.H.; White, F.J.; Rubio, I. 2003. Nutritional and suckling- mediates anovulation in beef cows. J.Anim.Sci. 81 (E. Suppl.2):E48-E59. [ Links ]
51. Wiltbank, M.C.; Gümen, A.; Sartori, R. 2002. Physiological classification of anovulatory conditions in cattle. Theriogenology 57:21-52 [ Links ]
52. Woodward, S.R.J.; Romera, A.; Beskow, W.; Lovatt, S.J. 2008. Better simulation modelling to support farming system innovation: review and synthesis, New Zealand Journal of Agricultural research, 51: 235-252. [ Links ]
53. Wright, I.A.; Rhind, S.M.; Russel, A.J.F.; Whyte, T.K.; McBean, A.J.; McMillen, S.R.; 1987. Effects of body condition, food intake and temporary calf separation on the duration of the postpartum anoestrus period and associated LH, FSH and prolactin concentrations in beef cows. Anim. Prod. 45, 395-402. [ Links ]
54. Yavas, Y.; Walton, J. 2000. Postpartum acyclicty in suckled beef cows. A review. Theriogenology, 54: 25-55. [ Links ]

