










Services on Demand
Journal

Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO
Related links
-
 Similars in
SciELO
Similars in
SciELO  uBio
uBio
Share

 Permalink
PermalinkInVet
On-line version ISSN 1668-3498
InVet vol.13 no.1 Ciudad Autónoma de Buenos Aires Jan./June 2011
COMUNICACIÓN CORTA
Primeras secuencias genómicas de apolipoproteínas en ñandú y choique
Silvestro, C.A.1; Huguet, M.J.1; Miño, O.S.1,2; Sarasqueta, D.V.3; Miquel, M.C.4; Iglesias,G.M.5,*; Jar, A.M.6
1Cátedra de Genética y
6Cátedra de Inmunología, Facultad de Ciencias Veterinarias, Universidad de Buenos Aires. Chorroarín 280,
(1427) Buenos Aires.
2Instituto de Virología, INTA. Los Reseros y Las Cabañas, (1686) Hurlingham, Buenos Aires;
3INTA, Estación Experimental Agropecuaria Bariloche, Modesta Victoria 4450 (8400), San Carlos de Bariloche, Río Negro;
4Actividad privada, ex Cátedra de Genética, Facultad de Ciencias Veterinarias, Universidad de Buenos Aires.
5Cátedra de Biología y Genética, Escuela de Veterinaria, Universidad Nacional de Río Negro, (8360) Choele Choel, Río Negro.
Correspondencia e-mail: Carolina Silvestro carolina@silvestro.com.ar
* Premio "Estímulo a la Investigación Científica 2011" en la categoría estudiante de la Facultad de Ciencias Veterinarias, Universidad de Buenos Aires.
Recibido: 15/08/11
Aceptado: 30/08/11
Resumen
El ñandú (Rhea americana) y el choique (Pterocnemia pennatta) son aves corredoras de gran tamaño autóctonas de Sudamérica, pertenecientes al orden Rheiformes. El objetivo de este trabajo fue identificar las secuencias genómicas de la apolipoproteína tipo II (apoVLDL-II), constituyente de la lipoproteína de muy baja densidad que es la encargada del transporte de triglicéridos al vitelo en formación. Se tomaron muestras de sangre de ñandúes y choique del "programa de cría de rheidos en cautiverio" de la EEA INTA Bariloche. Estos animales derivan de un núcleo formado a partir de huevos de animales silvestres. Se amplificó la región genómica de apoVLDL-II con 2 juegos de cebadores diseñados a partir de secuencias de aves presentes en el GenBank. Los fragmentos se clonaron, secuenciaron y analizaron. Las secuencias de aminoácidos de apoVLDL-II de choique y ñandú con respecto al resto de las aves presentaron un rango de similitud del 56.6% al 91.5%. El análisis filogenético de las secuencias aminoacídicas de apoVLDL-II mostró que choique y ñandú, junto a emu, forman un grupo bien fundamentado. Estas son las primeras secuencias de apolipoproteína en estas especies. El estudio de características de interés productivo es necesario para respaldar una futura mejora genética de la especie.
Palabras clave: Apolipoproteína; ApoVLDL-II; VLDLy; Rhea americana; Pterocnemia pennatta.
First genomic sequences of apolipoproteins in ñandú and choique
Summary
Ñandú (Rhea americana) and choique (Pterocnemia pennatta) are large, flightless birds native to South America, belonging to order Rheiformes. The aim of this study was to identify the genomic sequences of apolipoprotein type II (apoVLDL-II), a constituent of the very low density lipoprotein targeted to yolk, which transports triglycerides to the growing vitello. We sampled blood from animals belonging to the "rhea breeding in captivity program" of EEA INTA Bariloche, which came from a nucleus formed from wild animal's eggs. The genomic region of apoVLDL-II was amplified with 2 sets of primers designed on the bases of GenBank sequences of related species. The fragments were cloned, sequenced and analyzed. We found that apoVLDL-II of choique and ñandu has an amino acid identity between 56.6 and 91.5% with the species compared. Phylogenetic analysis of amino acid sequences shows that choique and ñandu, together with emu, form a well supported group. These are the first apolipoprotein sequences reported from these species. The study of productive characters is required to support future progress of the species.
Key words: Apolipoprotein; ApoVLDL-II; VLDLy; Rhea americana; Pterocnemia pennatta.
Introducción
El ñandú (Rhea americana) y el choique (Pterocnemia pennatta) son aves corredoras de gran tamaño autóctonas de Sudamérica, pertenecientes al orden Rheiformes; junto con el emu y el avestruz, pertenecen al superorden Palaeognatae y son considerados las aves vivientes más primitivas.
El interés sobre el conocimiento de estas especies ha crecido notablemente en los últimos años, debido a la concientización sobre la importancia de la conservación de especies nativas y a su desarrollo como alternativa para la producción de gran cantidad y variedad de insumos para diferentes industrias (frigorífica, peletera, ornamental, etc.)7,13,15. Las características genéticas, que dan base a las características productivas y reproductivas, son prácticamente desconocidas en el choique y en el ñandú. Dentro de estas características, las proteínas participantes de la vitelogénesis son de particular interés debido a la importancia del vitelo tanto para la reproducción como para la producción en las aves4,11.
Las lipoproteínas de muy baja densidad (VLDL) constituyen un grupo dentro de los complejos macromoleculares encargados del transporte de lípidos en el organismo. Estas poseen un diámetro de 30 a 70 nm y se encargan principalmente del transporte de triglicéridos de síntesis endógena a los diferentes tejidos12. En las aves existe un grupo particular de VLDL encargada de transportar y depositar triacilglicéridos endógenos directamente al vitelo (yema de huevo) en formación, denominada VLDLy4,11. Esta macromolécula se sintetiza en el hígado, y en las gallinas de postura hace que la yema del huevo crezca desde los 7 mm hasta los 35 mm de diámetro en los 7 días previos a la ovulación.
Algunas propiedades peculiares de la VLDLy, como su diámetro de 25-44 nm (tres veces menor a la VLDL plasmática) y su resistencia a la actividad lipolítica de la lipoproteinlipasa plasmática le permiten ser endocitadas de manera intacta por los folículos ováricos en desarrollo y depositarse en el vitelo sin ninguna disminución de su contenido lipídico4. Estas propiedades de la VLDLy se deben a la presencia de dos proteínas de síntesis inducida por estrógeno en su membrana anfotérica: la apolipovitelenina-I o apolipoproteína tipo II (apoVLDL-II) y la apolipoproteína B (apo-B) que forman polímeros (apoVLDL-II: apo-B) en una proporción de 46:1 moléculas3,6,17. Se postula que apoVLDL-II es la encargada de proteger a los triglicéridos transportados por la VLDLy de la acción de la lipoproteinlipasa durante el trayecto desde el hígado al oocito, así como también limitar el crecimiento de la VLDLy, originando una partícula de menor tamaño y de fácil endocitosis. De esta manera, apoVLDL-II se constituye en una proteína fundamental para el depósito de los lípidos en la formación del vitelo. Este proceso es de gran importancia, ya que es el alimento del futuro embrión y además influye sobre otras características productivas como por ejemplo, la incubabilidad14.
El objetivo del presente trabajo fue obtener y analizar las primeras secuencias de apoVLDLII en ñandú y choique como un primer acercamiento al estudio de los caracteres productivos de estas especies.
Materiales y métodos
Animales
Se utilizaron ñandúes y choiques adultos pertenecientes al "programa de cría de rheidos en cautiverio" de la EEA INTA Bariloche. Estos animales derivan de un núcleo formado a partir de huevos de animales silvestres.
Obtención de muestras
Se tomaron muestras de sangre entera con EDTA por punción de la vena braquial de las aves, en la cara interna del ala. Se utilizaron 20µl de sangre para extraer el ADN genómico, mediante el kit comercial Illustra blood genomicPrep Mini Spin Kit (GE Healthcare, Buckinghamshire, Reino Unido) con el que se obtuvieron entre 30 y 90 µg de ADN de excelente pureza.
Diseño de cebadores y amplificación de secuencias
Debido a la escasa información genética disponible sobre las apolipoproteínas aviares, se realizó un alineamiento múltiple a partir de la totalidad de las secuencias de apoVLDLII publicadas en la base de datos GenBank2. Las secuencias utilizadas de ganso común (Anser anser), pato salvaje (Anas platyrhynchos), codorniz (Coturnix coturnix) y gallo (Gallus gallus), corresponden a las regiones codificantes obtenidas por trascripción reversa del ARN mensajero del gen (ADNcopia o ADNc). Se utilizó además una segunda secuencia del gallo proveniente de las secuencias genómicas completas (contig genómico) obtenidas en el proyecto "Genoma Pollo". A este alineamiento se le agregó la secuencia nucleotídica codificante que hemos podido predecir a partir de la secuencia polipeptídica disponible de apoVLDL-II de emu (Dromaius novaehollandia)5. La importancia de esta última secuencia se debe a que esta especie pertenece al superorden Paleognathae, al igual que el ñandú y el choique. Las secuencias se empalmaron utlizando el programa BioEdit v7.0.999 y se alinearon mediante el programa ClustalX101; el alineamiento obtenido se corrigió manualmente utlizando el programa BioEdit v7.0.99.
A partir del análisis de estos alineamientos, se seleccionaron las regiones más conservadas del gen para diseñar los cebadores que se usaron para amplificar, por PCR, el gen completo de apoVLDL-II del ñandú y del choique.
Se diseñaron dos pares de cebadores que permiten amplificar dos segmentos solapantes. El par de cebadores denominados 2-3 directo (5'-CGCGGGGCTGAACTGGTAC-3') y reverso (5'-CAGCATCCGCCAAGAACTGACCA-3') permite amplificar la secuencia comprendida entre los nucleótidos 3201 al 3767 del contig genómico del gallo (Nº de Acceso NW_001471529), en el extremo 5' del gen, constituido por los exones 2 y 3 y el respectivo intrón. El par de cebadores 3-4 directo (5'-TGGTTGGTCATCCCCGATGCA-3') y reverso 5'-CATCCTTTGGTCTTTGATGCTGGGTT-3') permite amplificar la secuencia comprendida entre los nucleótidos 3721 al 4601 en el extremo 3' del contig genómico del gallo, constituido por los exones 3 y 4, más el correspondiente intrón. Las regiones amplificadas se esquematizan en la Figura 1.
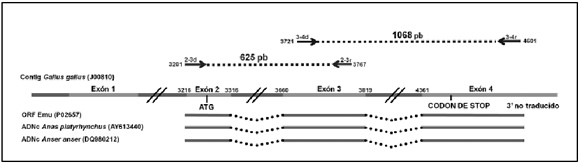
Figura 1. Esquema del gen de la apoVLDL-II: Se puede observar la posición del sitio de unión de los cebadores 2-3 directo (2-3d) y reverso (2-3r), 3-4 directo (3-4d) y reverso (3-4r). Los segmentos amplificados por PCR de choique y ñandú están representados como líneas punteadas entre flechas. Se aprecia la superposición del extremo 3´ del fragmento 2-3 con el extremo 5´ del framento 3-4. Se esquematizan los exones de algunas de las secuencias utilizadas en el diseño de cebadores: los exones como líneas macizas y los intrones como líneas punteadas.
La amplificación por PCR de ambos segmentos se realizó en un volumen final de 50µl de buffer Taq 1X que contenía: 100-500 ng de ADN molde, 25 picomoles de cada cebador, 25mM de dNTPs, 2mM de Cl2Mg y 5 U de Taq Polimerasa (Invitrogen Co, Carlsbad, EE.UU.). La reacción se llevó a cabo en un termociclador Techne TC412 (Barloworld Scientific Ltd., Staffordshire, Reino Unido) bajo un programa que consistía en 35 ciclos de un minuto a 94ºC, un minuto a 58ºC y un minuto a 72ºC, más una extensión final de 10 minutos a 72ºC. El resultado de la reacción se controló por electroforesis en un gel de agarosa al 1% teñido con bromuro de etidio.
Los productos obtenidos de los tamaños esperados se purificaron utilizando el kit QiaExII Gel extraction Kit (Qiagen, Hilden, Alemania), se ligaron en el vector pGEM T-Easy (Promega Co, Madison, EE.UU.) y se clonaron en E. coli JM109.
Secuenciación y análisis
Se seleccionaron 8 clones de ñandú y 8 clones de choique que contenían los amplicones de interés. Estos se purificaron mediante el kit Wizard Miniprep Extraction Kit (Promega, Madison, EE.UU.) y se enviaron para su secuenciación (Macrogen Inc, Seul, Corea del Sur). Las secuencias se analizaron con los programas de bioinformática ClustalX10, BioEdit v7.0.99 y BlastN1. El análisis filogenético se llevó a cabo mediante el método de máxima parsimonia, utilizando el programa TNT8. El soporte de los grupos se calculó con el método de Bootstrap y Jackknifing usando 100 repeticiones en cada caso.
Resultados
Se amplificaron en forma exitosa los segmentos 2-3 (625 bp) y 3-4 (1068 bp) de apoVLDL-II de ñandú y de choique. Las secuencias obtenidas se corrigieron manualmente de acuerdo con sus respectivos cromatogramas y mostraron coincidencia en sus regiones solapantes. Esto permitió empalmar las secuencias de los dos segmentos, de forma de generar una única secuencia consenso para ñandú y otra para choique, con una longitud total de 1595 bp cada una.
Se pudo demostrar la identidad de las secuencias de nucleótidos de apoVLDL-II tanto de ñandú como de choique. Sin embargo, el cálculo de homología no pudo realizarse debido a que las secuencias aviares disponibles en las bases de datos son escasas y en su mayoría parciales y de distintos largos. Por este motivo, se optó por realizar estos estudios sobre las secuencias aminoacídicas de apoVLDL-II disponibles en la base de datos GenBank. Para esto se eliminaron las regiones intrónicas y se tradujo la secuencia aminoacídica de apoVLDL-II de ñandú y choique. Se obtuvo un marco de lectura abierto de 108 codones, que es el marco de lectura completo y coincide con las secuencias polipeptídicas de otras aves. Los porcentajes de homología obtenidos con respecto a las secuencias disponibles de cada especie aviar, se muestran en la Tabla 1, agrupados según los grupos naturales definidos por la base de datos Taxonomy Browser16.
Tabla 1. Porcentajes de similitud entre las secuencias polipeptídicas de apoVLDL-II.
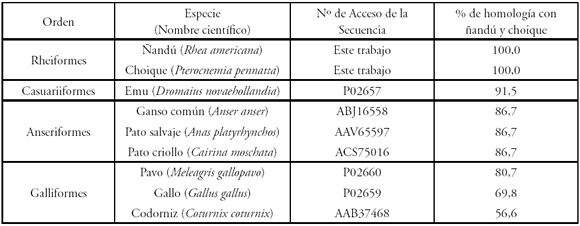
* Las secuencias parciales se encuentran publicadas en el GenBank bajo los números de acceso GU108456 (ñandú, 580 bp) y GU108457 (choique, 581 bp). Silvestro, C.A., Huguet, M.J., Feld, A., Sarasqueta, D.V., Miquel, M.C., Iglesias, G.M., 15/10/2009.
La región codificante de esta proteína mostró una alta conservación; solo se encontraron dos sustituciones aminoacídicas entre ñandú y choique, una ubicada en la posición 4 (serina por tirosina) y la otra en la posición 13 (leucina por isoleucina). Ambas posiciones corresponden al péptido señal de la proteína. Las secuencias del choique y del ñandú mostraron 7 sustituciones con respecto a la secuencia del emu, en las posiciones 44, 47, 55, 63, 68, 92 y 96. Las secuencias de ñandú, choique y emu presentan un aminoácido de más, asparagina en la posición 32. Las distintas sustituciones en las secuencias de aminoácidos pueden observarse en la Figura 2.

Figura 2. Alineamiento de las secuencias polipeptídicas de apoVLDL-II aviares: La secuencia de ñandú se utiliza como base del alineamiento. Con (.) se representan los aminoácidos idénticos a la secuencia de ñandú. Las sustituciones se marcan con las letras correspondientes al aminoácido en código de una letra. Con (-) se representa al bache generado por un aminoácido ausente.
A partir de estas secuencias se realizó un análisis filogenético que mostró la formación de tres grupos biológicos diferentes. Uno de estos agrupa a los conocidos taxonómicamente como Galliformes (gallo, pavo y codorniz), el otro reúne a los Anseriformes (ganso, pato salvaje y pato criollo) y el último agrupa a los Rheiformes (ñandú y choique) junto con el emu (Casuariiforme), los tres pertenecientes al superorden Paleognathae. El árbol de filogenia se puede observar en la Figura 3.
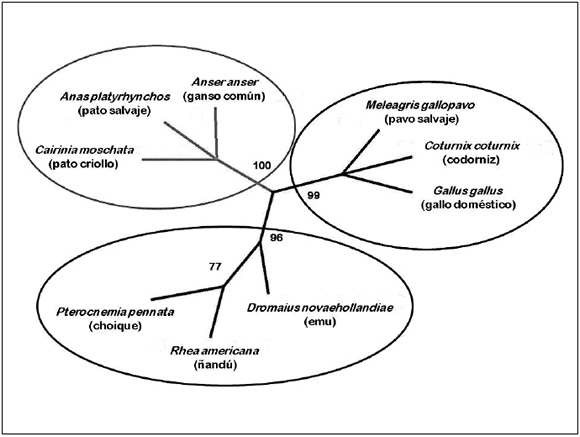
Figura 3. Árbol filogenético de las secuencias proteicas de apoVLDL-II: Se observa el agrupamiento de las secuencias en tres grupos biológicos. Los valores indicados en los nodos corresponden a los valores de soporte de ramas (Boostrap y Jacknife).
Discusión
Cuando comenzamos a trabajar sobre las secuencias de apoVLDL-II de ñandú y choique, encontramos que no había secuencias disponibles de especies relacionadas como el avestruz o el emu; de esta última sólo encontramos una secuencia parcial de aminoácidos, obtenida a partir de la secuenciación de la proteína cristalizada5. Esta falta de información nos llevó a buscar una especie que sirviese como referencia genómica.
En el estudio de genes y proteínas aviares se utiliza el gallo doméstico (Gallus gallus) como referencia, posiblemente por ser la especie más estudiada a nivel internacional. Sin embargo, las secuencias polipeptídicas tanto de ñandú como de choique mostraron un porcentaje de homología con respecto al gallo del 69,8%, mientras que con respecto al grupo de los anseriformes (pato y ganso) fue del 86,7%. Estos resultados nos indican que otras especies de aves como el pato y el ganso serían más apropiadas que el gallo como referencia genómica para futuros estudios moleculares en Rheiformes.
Los porcentajes de homología referidos en el párrafo anterior, se traducen como 28 y 27 sustituciones aminoacídicas entre las secuencias de ApoVLDL-II del ñandú y choique, respectivamente, en comparación con la secuencia del gallo. Estas diferencias decrecen a 13 sustituciones cuando se comparan las secuencias de ApoVLDL-II del ñandú y choique con respecto al pato salvaje, y a 12 sustituciones respecto al ganso común. Por otra parte, el análisis de las secuencias muestra la presencia de un aminoácido de más (asparagina en la posición 32) en el ñandú y el choique que aparece además en la proteína perteneciente a emu, lo que sugeriría una variante propia de las aves del superorden Palaeognathae, aunque se requieren estudios adicionales para su comprobación. Resulta de interés estudiar si este aminoácido extra podría estar involucrado en un cambio conformacional o funcional de la proteína.
Conclusiones
Este trabajo constituye el primer informe de la secuencia genómica y de la caracterización molecular y filogenética de la apolipoproteína II proveniente de ñandú y de choique.
Las proteínas de interés productivo y reproductivo, como son las involucradas en la vitelogénesis, poseen una gran importancia. La caracterización de los genes que codifican para estas proteínas podría aportar información valiosa para futuras mejoras genéticas en el ñandú y en el choique y para su aprovechamiento productivo, sin descuidar su conservación como especies autóctonas de Sudamérica.
Agradecimientos
Este trabajo fue financiado a través del proyecto V020, programación científica UBACyT 2008-2011, bajo la dirección de la Dra. María Cristina Miquel (2008-2010) y la co-dirección de la Magister Gabriela M. Iglesias (2008-2010), y la dirección de la Dra. Ana María Jar (2010-2011).
1. Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D. J. Basic local alignment search tool. J Mol Biol. 1990; 215(3):403-10 [ Links ]
2. Benson, D.A.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Wheeler, D.L. GenBank. Nucleic Acids Res. 2008; 36 (Database issue): D25-30. [ Links ]
3. Boyle-Roden, E.; Walzem, L. Integral apolipoproteins increase surface-located triacylglycerol in intact native apon-100-containing lipoproteins. J Lipid Res. 2005; 46:1624-32. [ Links ]
4. Bujo, H.; Hermann, M.; Lindstedt, K.A.; Nimpf, J.; Schneider, W.J. Low density lipoprotein receptor gene family members mediate yolk deposition. Review. J Nutr. 1997; 127(5 Suppl):801S-4S. [ Links ]
5. Dopheide, T.A.; Inglis, A.S. The amino acid sequence of a protein (apovitellenin I) from the low-density lipoprotein of emu egg yolk. Aust J Biol Sci. 1974; 27(1):15-21. [ Links ]
6. Edinger, R.S.; Mambo, E.; Evans, M.I. Estrogendependent transcriptional activation and vitellogenin gene memory. Mol Endocrinol. 1997; 11(13):1985-93. [ Links ]
7. Feld, A.; Silvestro, C.A.; Huguet, M.J.; Miquel, M.C.; Sarasqueta, D.V.; Iglesias, G.M. Conocimientos actuales sobre la genética del ñandú (Rhea americana) y el choique (Pterocnemia pennata). Revisión Biblioráfica. Rev Arg Prod Animal. 2011; 31(1):En prensa. [ Links ]
8. Goloboff, P.; Farris, J.; Nixon, K. TNT, a free program for phylogenetic analysis. Cladistics. 2008; 24:774-86. [ Links ]
9. Hall, T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. 1999; 41:95-8. [ Links ]
10. Larkin, M.A.; Blackshields, G.; Brown, N.P.; et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007; 23(21):2947-8. [ Links ]
11. Li, H.; Deeb, N.; Zhou, H.; Ashwel, C.M.; Lamont, S.J. Chicken quantitative trait loci for growth an body composition associated with the very low density apolipoprotein-II gene. Poult Sci. 2005; 84: 697-703 [ Links ]
12. Lin, C.T.; Chan, L. Localization of apoVLDL-II, a major apoprotein in very low density lipoproteins, in the estrogen-treated cockerel liver by immunoelectron microscopy. Histochemistry. 1982; 76(2):237-46. [ Links ]
13. Sarasqueta, D.V. Aspects of rearing, reproduction and hybridization of Darwin´s rhea or Choique (Rhea pennata syn. Pterocnemia pennata, spp pennata). Paper 3. Pages 35-44. Proceedings of the 3rd. International ratite Science Symposium of the World´s Poultry Science Association (WPSA) and XII World Ostrich Congress. 2005. [ Links ]
14. Sarasqueta, D.V. Incubación y cría de Choiques (Pterocnemia pennata). Comunicación Técnica Nº 108, EEA-Bariloche (INTA). S.C. de Bariloche. Argentina. 1995. [ Links ]
15. Sarasqueta, D.V. Cría de Ñandúes. Capítulo 19 en: Cría de Avestruces, Emúes y Ñandúes. 2ª Edición, Real Escuela de Avicultura, Barcelona España. 1997. [ Links ]
16. Taxonomy Browser. Disponible en línea en : http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/ [ Links ]
17. Walzem, R.L.; Hansen, R.J.; Williams, D.L.; Hamilton, R.L. Estrogen induction of VLDLy assembly in egglaying hens. J Nutr. 1999; 129(2S Suppl):467S-72S. [ Links ]

